
Abstract
CC chemokine receptor type-2 (CCR2) belongs to the G protein-coupled receptors superfamily, localized on cell surface of some immune-related cells, including monocytes and macrophages. CCR2 and its ligand CCL2 are involved in the progression of various diseases such as cancers. Therefore, CCR2-targeted monoclonal antibodies (mAbs) are needed for treatment and diagnosis. Previously, we successfully developed an anti-human CCR2 (hCCR2) mAb, C2Mab-9 (mouse IgG1, kappa), which is applicable for flow cytometry and immunocytochemistry. In this study, we investigated the critical epitope of C2Mab-9. We conducted enzyme-linked immunosorbent assay (ELISA) using several N-terminal peptides of hCCR2, and demonstrated that C2Mab-9 recognizes 11–29 and 21–39 amino acids of hCCR2. We further performed ELISA using 20 peptides, which include alanine substitution of hCCR2. C2Mab-9 lost the reaction to the alanine-substituted peptides of F23A, F24A, D25A, Y26A, and D27A. Among them, F23A, F24A, D25A, and Y26A did not block the C2Mab-9 reaction with U937 cells in flow cytometry. These results indicate that the critical binding epitope of C2Mab-9 includes Phe23, Phe24, Asp25, and Tyr26.
Introduction
Chemokines play pivotal roles in cellular function, including immune responses.(1,2) They are divided into four different subfamilies of XC, CC, CXC, and CX3C, depending on the number and position of cysteine residues at the N-terminus.(3,4) Chemokine receptors are G protein-coupled receptor (GPCR) with seven transmembrane regions.
CC chemokine receptor type-2 (CCR2) is expressed in multiple cells, including epithelial cells, monocytes, macrophages, and dendritic cells. CCR2 is involved in the regulation of migration and infiltration of immune-related cells.(5–8) CCR2 is the primary receptor of C-C motif chemokine 2 (CCL2)/monocyte chemoattractant protein-1. CCL2 also plays an important role in attracting the key immune regulators, including T lymphocytes, natural killer cells, and monocytes.(5,9) CCL2-CCR2 axis is correlated with many diseases such as immune disorders and cancers.(10,11)
In respiratory organs, CCL2 and CCR2-expressing macrophages and neutrophils contribute to the innate immune responses during viral and bacterial infection.(12) CCR2 induces neutrophil organ entry in sepsis.(13) High CCR2 levels in blood samples have been detected in patients with severe COVID-19.(14) Furthermore, CCL2 has been reported to be increased in several tumors, such as bladder,(15) bone,(16) and inflammatory breast cancers.(17) High CCR2 expression has been confirmed in the invasive lesion of breast cancers as well as CCL2 expression.(18) Inhibition of CCR2 has been reported to enhance the effectiveness of immune checkpoint inhibitors, such as an anti-programmed-cell death-1 monoclonal antibody (mAb) in mouse tumor models, and clinical trials are ongoing.(15,19)
We have produced numerous mAbs against membrane proteins, including EGFR,(20) HER2,(21) HER3,(22) CD20,(23) CD44,(24) CD133,(25) and podoplanin,(26–30) and determined their epitopes.(31–34) Moreover, we have also developed anti-GPCR mAbs, including anti-mouse CCR2,(35) mouse CCR3,(36) mouse CCR4,(37) mouse CCR8,(38) and human CCR9,(39) and also determined the binding epitope.(40) In this study, we performed the epitope mapping of C2Mab-9 by using enzyme-linked immunosorbent assay (ELISA) and flow cytometric methods.
Materials and Methods
Cell lines
The U937 (human histiocytic lymphoma) cell line was obtained from the Japanese Collection of Research Bioresources (Osaka, Japan). U937 cells were cultured in a Roswell Park Memorial Institute 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan) that was supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc., Waltham, MA), 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Cells were grown in a humidified incubator, which was supplied with 5% CO2 and 95% air at 37°C.
ELISA
The human CCR2 (hCCR2) peptides (Accession No. NM_001123041), including 3 N-terminal peptides (Table 1) and 20 point mutants (Table 2), were synthesized by utilizing PEPScreen (Sigma-Aldrich Corp., St. Louis, MO). Each peptide was immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific, Inc.) at a concentration of 10 μg/mL for 30 minutes at 37°C. After washing with phosphate-buffered saline (PBS) containing 0.05% Tween20 (PBST), wells were blocked with 1% bovine serum albumin (BSA)-containing PBST for 30 minutes at 37°C. The plates were then incubated with C2Mab-9 (1 μg/mL), followed by a 1:2000 dilution of peroxidase-conjugated anti-mouse immunoglobulins (Agilent Technologies, Inc., Santa Clara, CA). Enzymatic reactions were performed using the ELISA POD Substrate TMB Kit (Nacalai Tesque, Inc.). Optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA).
Identification of the C2Mab-9 Epitope Using N-Terminal Human CC Chemokine Receptor 2 Peptides
+++, OD655≧0.3; −, OD655 < 0.1.
Identification of the C2Mab-9 Epitope Using Alanine-Substituted Human CC Chemokine Receptor 2 Peptides
+++, OD655≧0.3; −, OD655 < 0.1.
Flow cytometry
U937 cells were washed with 0.1% BSA in PBS. C2Mab-9 (1 μg/mL) was incubated with each peptide (10 μg/mL) for 30 minutes at 4°C. U937 cells were treated with C2Mab-9+each peptide, and further with Alexa Fluor 488-conjugated anti-mouse IgG (1:1000). The fluorescence data were collected using a BD FACSLyric (BD Biosciences, Franklin Lakes, NJ).
Results
Epitope mapping using N-terminal hCCR2 peptides
In our previous study, we developed an anti-hCCR2 mAb (clone C2Mab-9) using synthetic peptide immunization method.(41) C2Mab-9 is suitable for in vitro applications, including flow cytometry and immunocytochemistry. To characterize the binding epitope of C2Mab-9 for hCCR2, we synthesized three N-terminal peptides: 1–19 amino acids (aa), 11–29 aa, and 21–39 aa (Table 1). The results of ELISA demonstrated that C2Mab-9 reacted with 11–29 aa and 21–39 aa of hCCR2 (Fig. 1A). These results are summarized in Figure 1B.

Determination of the C2Mab-9 epitope for hCCR2 by ELISA using N-terminal peptides.
Epitope mapping using alanine-substituted hCCR2 peptides
We further synthesized 20 different alanine-substituted hCCR2 peptides (Table 2). The results of ELISA demonstrated that C2Mab-9 reacted with point mutants, such as T21A, T22A, Y28A, G29A, A30G, P31A, S32A, H33A, K34A, F35A, D36A, V37A, K38A, Q39A, and I40A, as well as the 21–40 aa wild-type sequence (positive control) (Fig. 2A). In contrast, C2Mab-9 did not bind to point mutants, such as the F23A, F24A, D25A, Y26A, and D27A (Fig. 2A), indicating that Phe23, Phe24, Asp25, Tyr26, and Asp27 were determined to be the critical aa, which are included in the C2Mab-9 epitope. The results are summarized in Figure 2B.

Determination of the C2Mab-9 epitope for hCCR2 by ELISA using point mutants.
Flow cytometry using C2Mab-9 and alanine-substituted hCCR2 peptides
We then performed a blocking assay using flow cytometry. C2Mab-9 reacted with the U937 cell line (Fig. 3A). This reaction was completely neutralized by T22A and D27A. In contrast, F23A, F24A, D25A, and Y26A did not block the reaction of C2Mab-9 for U937 cells (Fig. 3A), indicating that Phe23, Phe24, Asp25, and Tyr26 of hCCR2 are critical for C2Mab-9 detection. The results are summarized in Figure 3B.
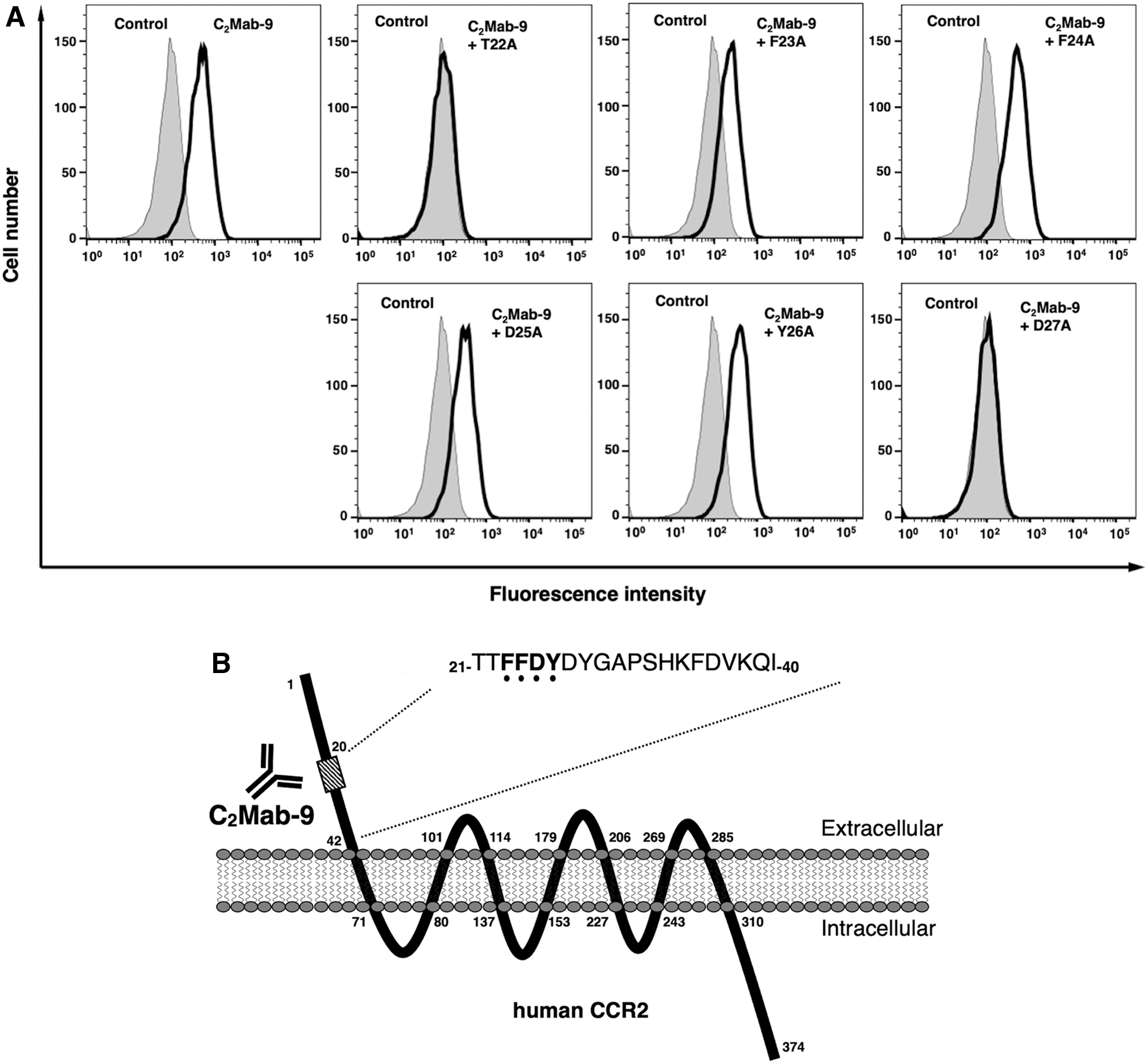
Flow cytometry using C2Mab-9 and peptides of hCCR2.
Discussion
Design of immunogen and epitope identification are essential for the development of mAbs and vaccines.(42,43) Furthermore, the determination of the mAb epitope can lead to the development of peptide tag systems for protein extraction, purification, or detection assays.(44–47)
We previously developed an anti-human CCR9 mAb (clone C9Mab-1) by using Cell-Based Immunization and Screening method and clarified that the epitope of C9Mab-1 is located on the N-terminus of the human CCR9.(39,40) Then, we have developed several anti-GPCR mAbs, including anti-mouse CCR2,(35) human CCR2,(41) mouse CCR3,(48) and mouse CCR4(37) by immunizing mice or rats with the N-terminal peptides of GPCRs.
In this study, we identified the critical epitope of C2Mab-9 as Phe23, Phe24, Asp25, Tyr26, and Asp27 using ELISA (Fig. 2). In contrast, blocking assay using flow cytometry revealed that Phe23, Phe24, Asp25, and Tyr26 of hCCR2 were critical epitope of C2Mab-9 (Fig. 3). This discrepancy might be caused by the condition of peptides: solid phase of ELISA or liquid phase of flow cytometry. We have previously developed various novel epitope mapping system, named RIEDL insertion for epitope mapping (REMAP)(49–52) and histidine-tag insertion for epitope mapping (HisMAP) method.(53) These methods could be applied to determine linear and structural epitopes. Therefore, we should determine the binding epitope of C2Mab-9 using REMAP and HisMAP methods in the future study.
CCR2 is a seven-transmembrane receptor with four extracellular regions (Figs. 2B and 3B). N-terminal domains of some GPCRs, such as CCR2, CCR3, CCR5, and CXCR1, have been determined as their ligand-binding sites.(54) The several aa in the N-terminal region of CCR2B (predominant isoform of CCR2) were reported to be important for CCL2-triggered cell migration and lamellipodium formation,(55) indicating that C2Mab-9 might be advantageous for the functional study about the CCL2-CCR2 axis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Nos. JP22ama121008 (to Y.K.), JP22am0401013 (to Y.K.), JP22bm1004001 (to Y.K.), JP22ck0106730 (to Y.K.), and JP21am0101078 (to Y.K.).