
Abstract
An anti-mouse CXC chemokine receptor 6 (mCXCR6) monoclonal antibody (mAb), Cx6Mab-1, was developed recently. Cx6Mab-1 is applicable for flow cytometry, Western blotting, and enzyme-linked immunosorbent assay. The purpose of this study is to determine the binding epitope of Cx6Mab-1 using 2 × alanine mutated mCXCR6. Analysis of flow cytometry revealed that Cx6Mab-1 did not recognize S8A-A9G, L10A-Y11A, D12A-G13A, and H14A-Y15A mutants of mCXCR6. The results clearly indicate that the binding epitope of Cx6Mab-1 includes Ser8, Ala9, Leu10, Tyr11, Asp12, Gly13, His14, and Tyr15 of mCXCR6. The successful determination of the Cx6Mab-1 epitope might contribute to the pathophysiological investigation of mCXCR6.
Introduction
The CXC chemokine receptor 6 (CXCR6), also known as CD186, is a G protein-coupled receptor (GPCR) with seven transmembrane regions. CXCR6 is preferentially expressed on T cells, including memory T cells, cytotoxic T lymphocytes (CTLs), helper T type 1 cells, and natural killer (NK) cells.1–3 In hepatic CXCR6+ NK cells, the survival of the T cells depends on the signaling of CXCR6. 4
T cells that participate in numerous immune responses are recruited through the interaction with a chemokine, CXC chemokine ligand 16 (CXCL16).5,6 CXCL16 is a specific chemokine ligand for CXCR6, which is expressed constitutively on liver sinusoidal endothelial cells, spleen, intestine, lungs, and skin, and is known to have transmembrane and soluble forms.7,8
The roles of CXCR6 and CXCL16 have been actively investigated in their association with diseases including cancer and viral infections. CXCR6 was found as a coreceptor on CD4+ and CD8+ T cells for the human immunodeficiency virus type 2. 9 During acute hepatitis B virus infection, liver-resident CXCR3+CXCR6+ γδ T cells work a protective role by producing high level of IFN-γ. 10 CXCR6+ NK cells contribute to immune memory against antigens such as viruses and haptens. 11 CXCR6-expressing T cells play pivotal roles in infection responses.
In tumors, CXCL16/CXCR6 expression has been confirmed in various cancers. 12 Tumor-infiltrating CXCR6+CD8+ T cells exert antitumor effect in immunotherapy. Anti-programmed cell death-1 antibodies are less effective in CXCR6 knockout mice. 1 In tumor microenvironment, CXCR6 in CTLs and trans-presented interleukin-15 promote the survival and local expansion of CTLs, maximizing subsequent antitumor effects. 13 In contrast, upregulation of CXCL16 supports cancer progression and results in poor prognosis. 12 High expression of CXCL16/CXCR6 has both positive and negative prognostic effects in cancer, requiring further study. The functional antibodies against CXCR6 and CXCL16 are required for therapy and diagnosis.
We have successfully developed various anti-GPCR monoclonal antibodies (mAbs), including anti-mouse CCR2, 14 human CCR2, 15 mouse CCR3,16–18 mouse CCR4, 19 mouse CCR8, 20 human CCR9, 21 and mouse CXCR6 (mCXCR6). 22 Furthermore, we performed epitope mapping about these mAbs.23–25 The estimated epitope of Cx6Mab-1 was also determined by enzyme-linked immunosorbent assay (ELISA) experiments using two alanine scanning methods. 26 In this study, we perform epitope analysis using two sequential alanine (or glycine) substitution strategies in a system of mutated mCXCR6-overexpressed cells that maintain structural features by flow cytometry.
Materials and Methods
Cell lines
Chinese hamster ovary-K1 (CHO-K1) cells were purchased from the American Type Culture Collection (Manassas, VA). The mCXCR6 mutation plasmids, pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan), containing an N-terminal PA tag,27–32 were transfected into CHO-K1 cells using a Neon Transfection System (Thermo Fisher Scientific, Inc., Waltham, MA). The PA tag comprises 12 amino acids, GVAMPGAEDDVV, and is recognized by NZ-1 mAb.27,33
Cells transiently transfected with wild type (WT) or two alanine (or glycine) residues-substituted mCXCR6 mutants were cultured in a Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Thermo Fisher Scientific, Inc.), 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.), and 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA). Then, cells were grown in a humidified incubator, which was supplied with 5% CO2 and 95% air at 37°C.
Production of mCXCR6 2 × alanine mutants
Synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding mCXCR6 (Accession No. NM_030712) with N-terminal PA tag was subcloned into a pCAG-Ble vector. Plasmids were transfected using a neon transfection system. Substitutions of consecutive amino acids with two alanine (or glycine) in the mCXCR6 sequence were produced by QuikChange Lightning Site Directed Mutagenesis Kits (Agilent Technologies, Inc., Santa Clara, CA). Polymerase chain reaction fragments-bearing desired mutations were inserted into the pCAG-Ble vector using KAPA HiFi HotStart ReadyMix (NIPPON Genetics Co., Ltd., Tokyo, Japan).
Flow cytometry
Transiently transfected CHO-K1 cells were detached by 0.25% trypsin in 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.) and collected using 10% FBS containing RPMI 1640 medium. After washing with 0.1% bovine serum albumin (BSA; Nacalai Tesque, Inc.) in phosphate-buffered saline (PBS; Nacalai Tesque, Inc.), cells were treated with anti-mCXCR6 mAb (Cx6Mab-1; 1 μg/mL) or NZ-1 (anti-PA tag; 1 μg/mL) for 30 minutes at 4°C, followed by incubation with Alexa Fluor 488-conjugated anti-rat IgG (1:1000; product no. 4416; Cell Signaling Technology, Inc.). Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Biotechnology Corp., Tokyo, Japan).
Results and Discussion
Epitope mapping of Cx6Mab-1 with alanine-substituted mCXCR6 in CHO-K1
We previously developed an anti-mCXCR6 mAb (Cx6Mab-1) by N-terminal peptide immunization of mCXCR6. 22 To identify the binding epitope of Cx6Mab-1, we produced 10 mutants of mCXCR6, in which sequential two amino acids were replaced to two alanine (or glycine) residues. This method was named as “2 × Ala-scan method.”
Based on past ELISA analysis, we could not determine the epitope with 1 × Ala-scan method 26 ; therefore, we introduced 2 × Ala-scan method using cells in this study. For instance, a mutant (D2A-D3A) indicates the alanine substitution of second Asp and third Asp of mCXCR6 protein. As shown in Figure 1A, all 2 × Ala mutants and WT mCXCR6 containing the N-terminal PA tag were recognized by NZ-1 (an anti-PA tag mAb), indicating that the expression level of each construct was sufficient. Cx6Mab-1 recognized D2A-D3A, G4A-H5A, Q6A-E7A, E16A-G17A, D18A-F19A, and W20A-L21A, but not S8A-A9G, L10A-Y11A, D12A-G13A, and H14A-Y15A (Fig. 1B).
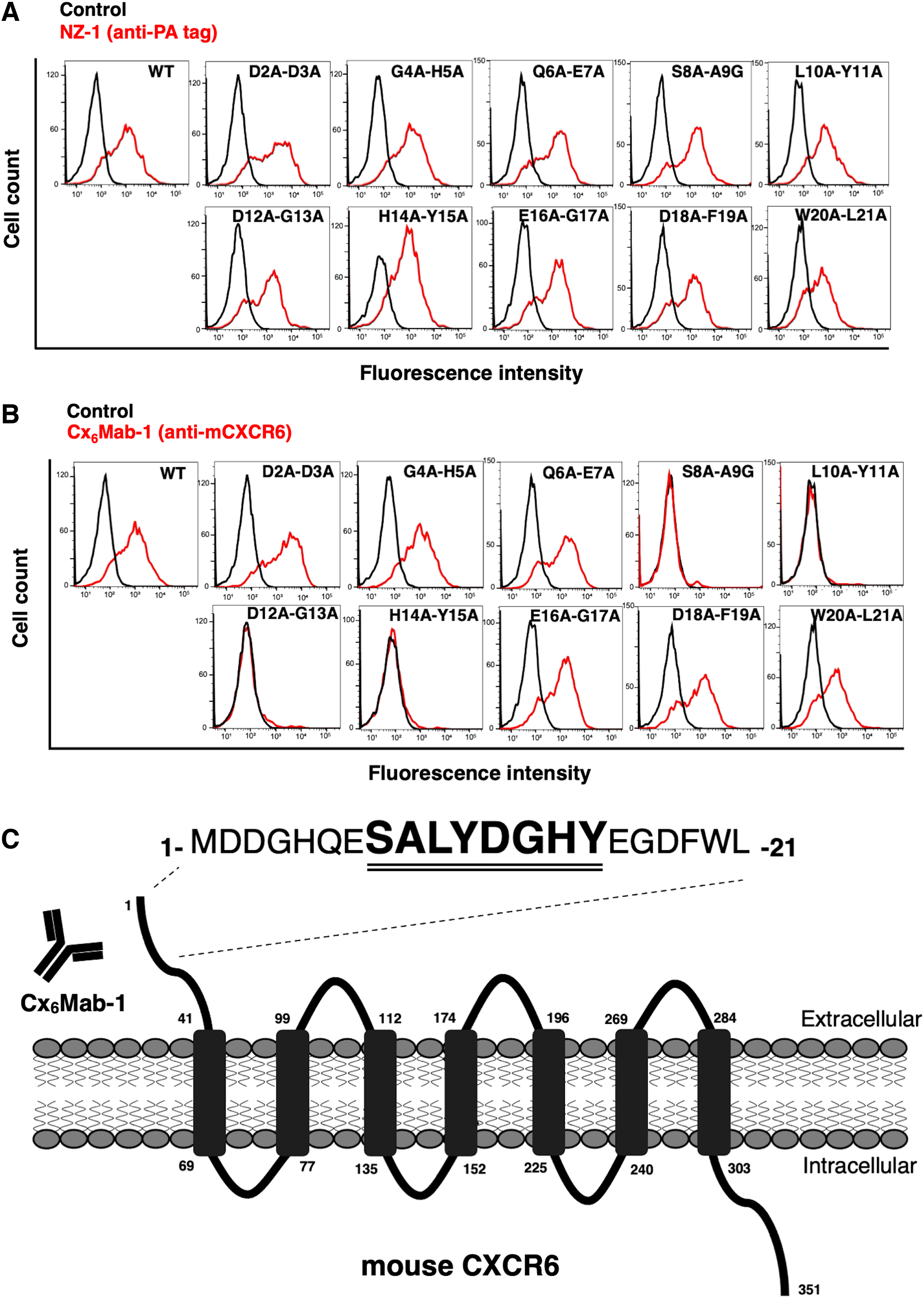
Epitope determination of Cx6Mab-1 using 2 × Ala (or Gly) substituted mutants by flow cytometry.
These results suggest that Ser8, Ala9, Leu10, Tyr11, Asp12, Gly13, His14, and Tyr15 are included in the critical epitope of Cx6Mab-1. The results are shown as a schematic illustration in Figure 1C. Previous results of ELISA experiments indicated that Ser8, Ala9, Leu10, Tyr11, Gly13, and His14 were the critical epitopes of Cx6Mab-1, we speculate that this discrepancy is due to the structural differences between peptides and proteins. This 2 × Ala-scanning strategy could be another option to identify the epitope of mAbs even if the conventional alanine scanning is not effective.
Several GPCRs, including CCR2, CCR3, CCR5, CCR9, and CXCR1, interact with ligands at the N-terminal region.34,35 A molecular dynamics simulation about docking of CXCL16–CXCR6 was reported. 36 Cx6Mab-1 may block CXCL16 binding, although further approach is needed. CXCL16 is highly expressed by tumor cells, including pancreatic cancer, and tumor-infiltrating immune cells. Since CXCR6-armed T cells enhance the recognition and lysis of tumor, 37 Cx6Mab-1 is expected to be one of the immunotherapy strategies by applying modalities such as bispecific antibodies against tumor antigen.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers JP22ama121008 (to Y.K.), JP22am0401013 (to Y.K.), JP22bm1004001 (to Y.K.), JP22ck0106730 (to Y.K.), and JP21am0101078 (to Y.K.).