
Abstract
Acute respiratory distress syndrome is a life-threatening acute lung injury (ALI) characterized by the destruction of alveoli leading to pulmonary edema. The infiltration and activation of inflammatory cells and production of inflammatory cytokines are both involved in the pathogenesis of ALI. Here, we show that the infiltration of neutrophils, major inflammatory cells causing ALI, into the lung is mediated by sialyl Lewis x (sLex) glycans, which can be efficiently suppressed by a monoclonal antibody (mAb) against these glycans. In fucosyltransferase-IV and -VII double-deficient mice lacking sLex expression, neutrophil infiltration into the lung was significantly suppressed compared with that observed in wild-type mice in a lipopolysaccharide (LPS)-induced ALI model. Administration of a highly specific anti-sLex mAb F2 3 hours after LPS administration significantly suppressed pulmonary neutrophil infiltration, accompanied by the reduced induction of inflammatory cytokines. It was consistently indicated from ex vivo cell rolling assay that mAb F2 blocked the rolling of mouse neutrophils on P-selectin-expressing cells. Overall, these results indicate that the sLex glycan could serve as a therapeutic target against ALI, and also that mAb F2 would be useful for specific targeting of this glycan.
Introduction
Acute respiratory distress syndrome (ARDS) is a life-threatening clinical complication of acute lung injury (ALI), which is characterized by the destruction of alveoli that leads to pulmonary edema. 1 In critically ill patients, ALI is a major cause of respiratory failure, 2 yet, the underlying mechanisms by which it initiates and transmits lung injury are still unclear. 3 Its clinical features include accumulation of neutrophils and the destruction of alveolar epithelial tissue and pulmonary capillaries, 1 resulting in severe pulmonary inflammatory responses.4,5 Currently, there are no effective treatments for this, and thus the morbidity and mortality of the disease remain high. 6 As a result, it is of great importance to develop an effective treatment.
To date, approaches to ALI treatment have been based on two strategies: ventilated 7 and nonventilated. In recent years, the development of physiological research on ALI has led to the continuous development of drug therapy, 8 as opposed to the use of external machinery for treatment. Among these, neutrophils are considered to be significantly involved in the pathogenesis of inflammation in ALI.9,10 Studies have shown that the neutrophil infiltration into the bronchoalveolar space caused pulmonary edema and damaged the inner epithelial tissues. 11
Neutrophils are recruited to tissues by sequential interactions with endothelial cells, that is, rolling mediated by selectins, integrin activation by chemokines, firm adhesion by integrins, and transmigration. 12 During the inflammatory response, stimulation from inflammatory mediators triggers changes in the endothelial surface thereby inducing the E- and P-selectin expression.13–15 Similar roles are played by the two selectins in neutrophil invasion. E- and P-selectin interact with the sialyl Lewis x (sLex) glycans on P-selectin glycoprotein ligand 1 (PSGL-1) expressed on neutrophils, which allows them to be captured in the circulation and roll on the endothelial surfaces. 16 Previous studies have shown that PSGL-1 on neutrophils in E- and P-selectin double-deficient mice could not mediate their rolling on the endothelial surfaces, thereby preventing leukocytes from carrying out the cascade of inflammatory events.17,18
In this study, we determine the role of sLex glycans in neutrophil infiltration into the lungs of ALI mice, by using fucosyltransferase-IV and -VII double-deficient (Fut4/7 DKO) mice lacking sLex expression. To examine the therapeutic effects of blocking sLex glycans in ALI, we also examine the effects of a highly specific anti-sLex monoclonal antibody (mAb) F2 reactive with sLex glycans expressed in both humans and mice here. 19 Based on the findings described in this study, it is expected that the sLex glycan could become a therapeutic target against ALI, and furthermore that mAb F2 would be useful as a therapeutic agent against ALI.
Materials and Methods
Animals
C57BL/6J mice (female, 8–10 weeks old) were obtained from Charles River Laboratories Japan. Fut4/7 DKO mice were bred and housed as previously described. 20 All the animal experimental protocols were approved by the Animal Research Committee of Chiba University.
Induction of mouse ALI by lipopolysaccharide
After being anesthetized using 3% isoflurane, mice were received nasal inhalation of lipopolysaccharide (LPS) (50 μg per mouse dissolved in 50 μL sterile phosphate-buffered saline [PBS]; Sigma-Aldrich) to induce ALI. In some experiments, 200 μg of mAb F2 in 200 μL sterile PBS was intravenously injected 3 hours after LPS administration, while 200 μL sterile PBS was injected into control mice. Then, 24 hours after LPS administration, the mice were euthanized and the lungs and bronchoalveolar lavage fluid (BALF) were collected. To collect BALF, a 22G indwelling needle was inserted from the tip of the trachea. The lungs were then flushed with 1.5 mL of 2% fetal bovine serum-PBS. For hematoxylin and eosin (H&E) staining experiments, 0.5 mL of optimal cutting temperature (O.C.T) compound (Sakura, Tissue-Tek) was slowly injected into the lungs from the trachea, and the lung tissues were embedded in O.C.T. compound and stored at −80°C until use.
For reverse transcription real-time quantitative polymerase chain reaction (RT-qPCR) experiments, lung tissues were preserved in RNA later reagent at −80°C until further use.
Flow cytometry of leukocytes in the BALF
The BALF was filtered using a 70 μm nylon mesh before being centrifuged at 440 × g for 5 minutes at 4°C. The supernatant was then removed and cells in the pellet were suspended in 0.1% bovine serum albumin (BSA)-PBS and Fc-blocked using anti-mouse CD16/32 antibody (Clone: 2.4G2; BioLegend) and incubated at 4°C for 10 minutes. The cells were then incubated at 4°C for 20 minutes with the following antibodies: APC-Cy7-anti-mouse CD45 (Clone: 30-F11; BioLegend), APC-anti-mouse CD11b (Clone: M1/70; BioLegend), and Alexa488-anti-mouse Gr-1 (Clone: RB60-8C5; BD Biosciences). After washing with 0.1% BSA-PBS, the cells were resuspended in filtered PBS and subjected to flow cytometry using CytoFLEX (Beckman Coulter). Data processing and analysis were done with FlowJo software 10.6.1 (BD Biosciences).
Real-time-qPCR
Using TRIzol reagent (Thermo Fisher Scientific), total RNA was extracted from the upper lobe of the lungs, while complementary DNA (cDNA) was synthesized using ReverTra Ace qPCR RT Master Mix (Toyobo). Using NanoDrop One (Thermo Fisher Scientific), the cDNA concentration was determined. THUNDERBIRD SYBR qPCR Mix (Toyobo) was used for RT-qPCR, and the expression of messenger RNA in each sample was normalized to that of β-actin by the ΔΔCt method using CFX96 Touch Real-Time PCR Detection System (Bio-Rad Laboratories). The following primer sets were used: β-actin (Actb), 5′-CATCCGTAAAGACCTCTATGCCAAC-3′ and 5′-ATGGAGCCACCGATCCACA-3′; IL-1α (Il1a), 5′-ACGTCAACAACGGGAAGAT-3′ and 5′-AAGGTGCTGATCTGGGTTGG-3′; IL-1β (Il1b), 5′-TGCCACCTTTTGACAGTGATG-3′ and 5′-TGATGTGCTGCGAGATT-3′;IL-6 (Il6), 5′-CACTGTTCTCTGGAAATCG-3′ and 5′-TTGTATCTCTGGAAGTTTCAGATTGTT-3′.
H&E staining
Using a cryostat (HM525; Thermo Fisher Scientific), the lung tissues that had been frozen in O.C.T. compound were cut into 7 μm thick sections. The frozen sections were fixed for 5 minutes using 10% buffered formalin solution. The sections were then washed with PBS, and stained with hematoxylin for 5 minutes. After washing with running water, the sections were stained with eosin for 15 minutes. Using graded ethanol and xylene, the sections were then dehydrated. Thereafter, the dehydrated sections were mounted using Mount-Quick (Daido Sangyo) and observed using a BZ-9000 fluorescence microscope (Keyence).
Preparation of neutrophils
Using three-layer Percoll, neutrophils were prepared from the bone marrow. 21 To collect bone marrow cells, all femurs and tibias of mice were harvested, bones were dissected, and bone marrow cells were washed extensively in 2 mL RPMI-1640 (Sigma-Aldrich) using a 26G needle. The collected bone marrow cells were then pelleted by centrifugation at 4°C and 440 × g for 5 minutes, before being suspended in 2 mL Hanks' balanced saline solution (HBSS)-ethylenediaminetetraacetic acid (EDTA) (HBSS containing 15 mM EDTA, and 1% BSA). 22
This cell suspension was then gently placed on top of a three-layer gradient of 78%, 69%, and 52% Percoll (diluted with HBSS, 100% Percoll = 9-parts Percoll +1-part 10 × HBSS, 2 mL each) and centrifuged at room temperature and 1500 × g for 30 minutes without braking. Neutrophils were carefully collected from the layer between 69% and 78% and then suspended in 2 mL HBSS-EDTA. After centrifugation at 4°C, 440 × g for 5 minutes, the purified neutrophils were subjected to flow cytometry and rolling assay.
Neutrophil rolling assay
Human P-selectin (CD62P)-expressing Chinese hamster ovary (CHO)-K1 cells were grown in 35-mm culture dishes (Corning). Neutrophils were labeled using 1 μM carboxyfluorescein succinimidyl ester (CFSE) (Thermo Fisher Scientific) fluorescently for 20 minutes at 37°C and then incubated for 10 minutes at room temperature either with, or without, 10 μg/mL mAb F2. During the incubation period, the culture dishes were set at the bottom of the parallel plate flow chambers (GlycoTech Co.). CFSE-labeled neutrophils that had been incubated with or without mAb F2 were resuspended in rolling assay buffer (0.1% BSA, 20 mM HEPES-NaOH, 150 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, and pH 7.4) at a density of 4 × 106 cells/mL.
Using a Model 11 Plus syringe pump (Harvard Apparatus Co.), the cell suspension was introduced into the flow chamber at a wall shear stress of 1.5 dynes/cm2. Images were taken using an OptiMOS™ scientific complementary metal–oxide–semiconductor camera (QImaging Co.) equipped with an Axiovert S100 inverted microscope (Zeiss) and analyzed using the Fiji ImageJ software.
Statistical analysis
Data are shown as mean ± standard error of the mean. The Student's t-test and Dunnett's multiple comparisons test utilizing one-way analysis of variance were used to assess statistical significance. Statistical significance was set at p < 0.05. All statistical evaluations were carried out utilizing GraphPad Prism 7.0.
Results
Neutrophil infiltration to the lung in ALI was suppressed in Fut4/7 DKO mice
Although the expression of sLex glycans on neutrophils has been well-documented over the decades, their role in neutrophil infiltration into the lungs in ALI has not yet been verified. Thus, here, we induced ALI in wild-type (WT) and Fut4/7 DKO mice lacking sLex and compared the respective levels of infiltration of neutrophils into the lungs. The schedule for the induction of inflammation is shown in Figure 1A.
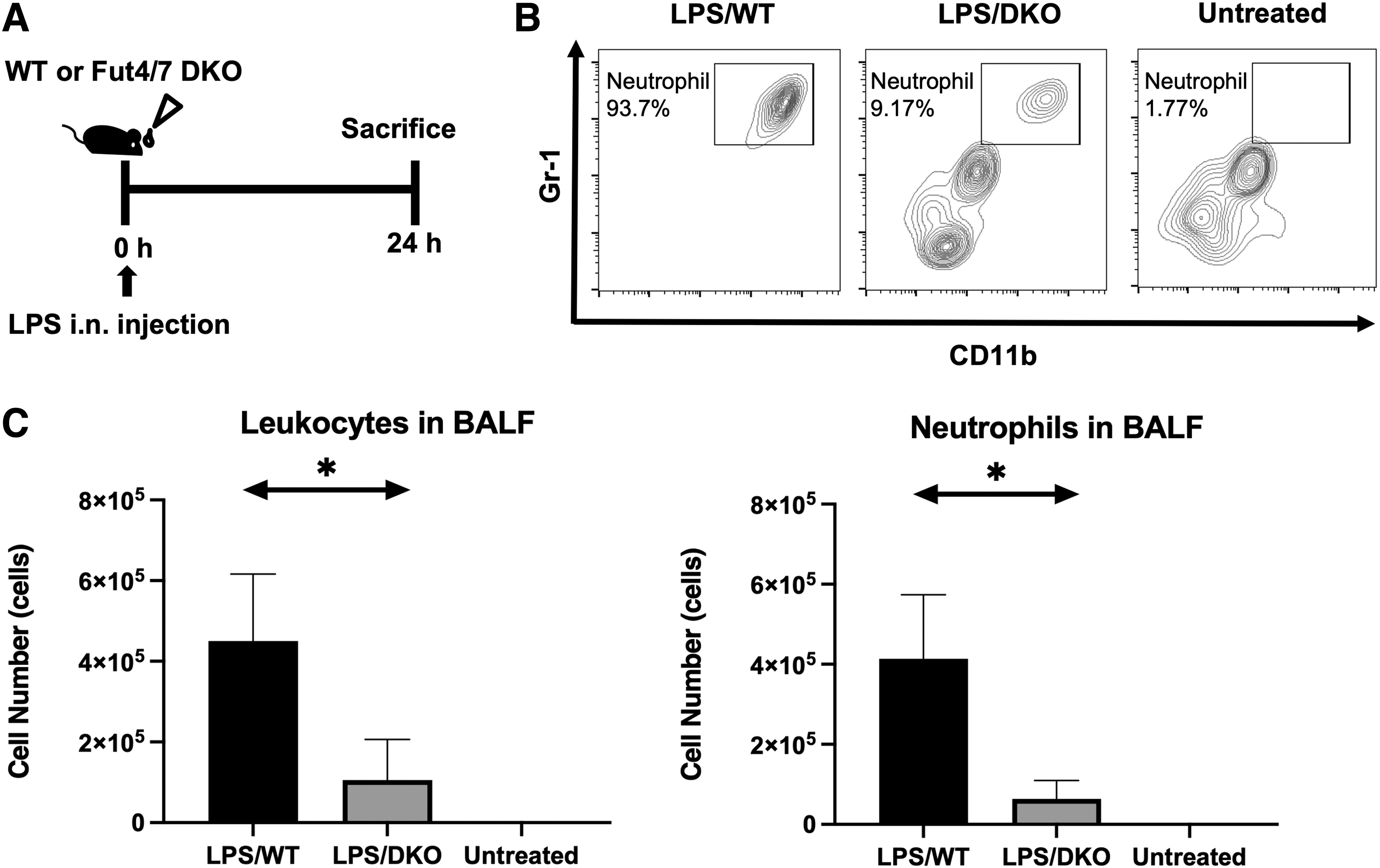
Suppression of neutrophil infiltration to the lung in Fut4/7 DKO mice in ALI.
Then, 24 hours after intranasal administration of LPS, the BALF of mice was subjected to flow cytometry. As shown in Figure 1B, the BALF of LPS-treated WT mice showed a more severe degree of neutrophil infiltration than that of untreated mice. Cells that were doubly positive for CD11b and Gr-1 represented neutrophils. In contrast, the BALF of LPS-treated Fut4/7 DKO mice showed significantly lower neutrophil infiltration. In addition, the total number of leukocytes (Fig. 1C, left) and neutrophils (Fig. 1C, right) in BALF was reduced. These results indicated that neutrophil infiltration into the lungs in ALI was dependent on sLex glycans synthesized by fucosyltransferase-IV and -VII.
Neutrophil infiltration to the lung in ALI was suppressed by mAb F2
To determine whether sLex glycans can serve as therapeutic targets against ALI, we examined the effects of anti-sLex mAb F2 on neutrophil infiltration in ALI. As shown in Figure 2A, mice were administered F2 mAb 3 hours after nasal LPS inhalation. The degree of neutrophil infiltration in the BALF of the F2-administered group was observed to be lower than that in the BALF of the control group (Fig. 2B). Leukocyte (Fig. 2C, left) and neutrophil numbers (Fig. 2C, right) in BALF were significantly diminished in the F2-administered group. These results indicated that mAb F2 had suppressive effects on neutrophil infiltration after the onset of LPS-induced ALI.

mAb F2 suppresses neutrophil infiltration.
mAb F2-administered mice showed reduced leukocyte infiltration in the lung tissue in ALI
To further determine leukocyte infiltration into lung tissues, histological examination was performed using lungs from the PBS- or F2-administered mice that had been treated with LPS (Fig. 3). Following cryosectioning of the lungs, tissue sections were subjected to H&E staining. As a result, a higher level of leukocyte infiltration around the bronchioles was observed in the PBS-administered group compared with that observed in the F2-administered group.
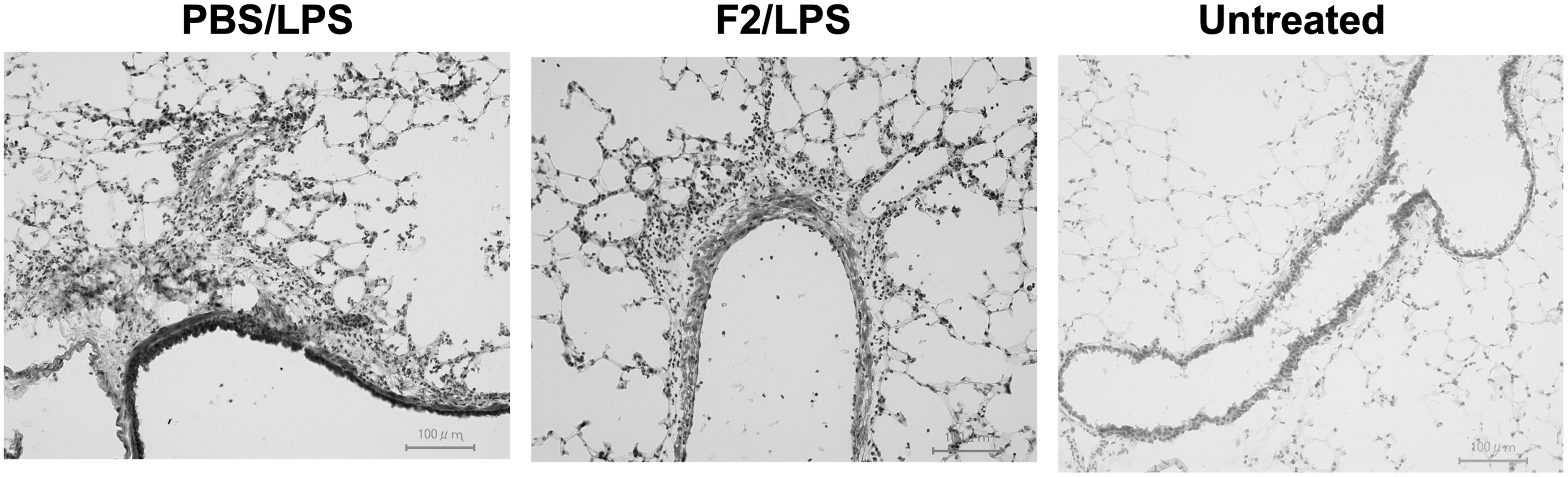
Decreased neutrophil infiltration in mAb F2-administered mice. Frozen sections of the lungs from PBS-injected (LPS/PBS) or F2-injected (LPS/F2) mice that had been treated with or without (Untreated) LPS were stained with H&E. Bars, 100 μm. H&E, hematoxylin and eosin.
mAb F2 suppressed inflammatory cytokine expression in the lungs in ALI
The blockage of sLex suppressed the accumulation of neutrophils in the lungs, which led to further investigation on the inflammatory cytokine expression in the lungs of LPS-administered mice. Total RNA was extracted from the lungs from PBS- and F2-administered mice in ALI and the expression of inflammatory cytokines, including IL-1α, IL-1β, and IL-6, were subsequently examined by RT-qPCR (Fig. 4). All of these cytokines were expressed significantly lower in the F2-administered group than in the PBS-administered group. These results indicated that mAb F2's blockage of neutrophil infiltration caused a considerable reduction in the inflammatory cytokine response in ALI.
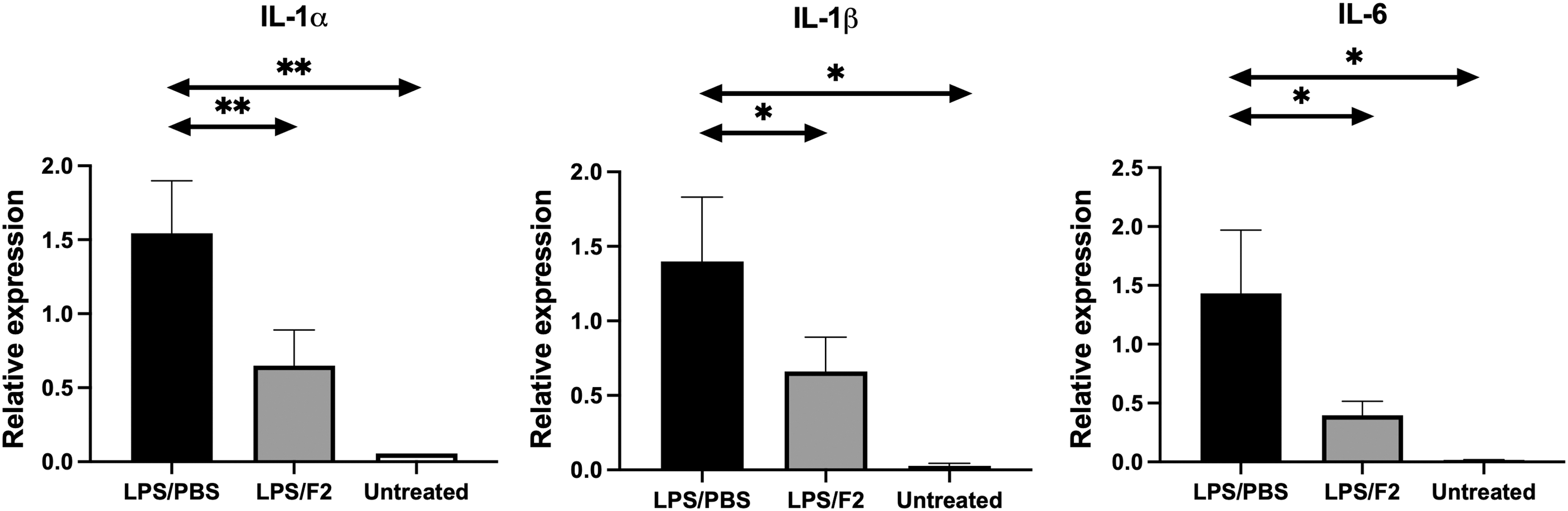
mAb F2 suppresses inflammatory cytokine expression in the lungs in ALI. The messenger RNA expression of IL-1α (Il1a), IL-1β (Il1b), and IL-6 (Il6) was examined in the lungs of PBS-injected (LPS/PBS) or F2-injected (LPS/F2) mice that had been treated with or without (Untreated) LPS by quantitative real-time polymerase chain reaction. n = 4. *p < 0.05, **p < 0.01.
sLex mediates neutrophil rolling by binding to P-selectin
The ex vivo cell-rolling assay is useful for examining the earliest stage of the sequential leukocyte infiltration steps mediated by selectins. We thus prepared neutrophils from mouse bone marrow by Percoll density gradient centrifugation and used them in an ex vivo cell rolling assay (Fig. 5A). These collected neutrophils strongly expressed sLex glycans on their surfaces, as determined by flow cytometric analysis using mAb F2 (Fig. 5B). The rolling assay was then performed using neutrophils that had been treated with or without mAb F2. P-selectin-expressing CHO cells, which mimic P-selectin-expressing endothelial cells at the sites of inflammation, were used as the cell layer here. In Figure 5C, the rolling tracks are shown as white lines. A significantly larger number of rolling neutrophils were found in the PBS-treated group than in the mAb F2-treated group, indicating that sLex was essential for the neutrophil rolling on P-selectin-expressing cells.
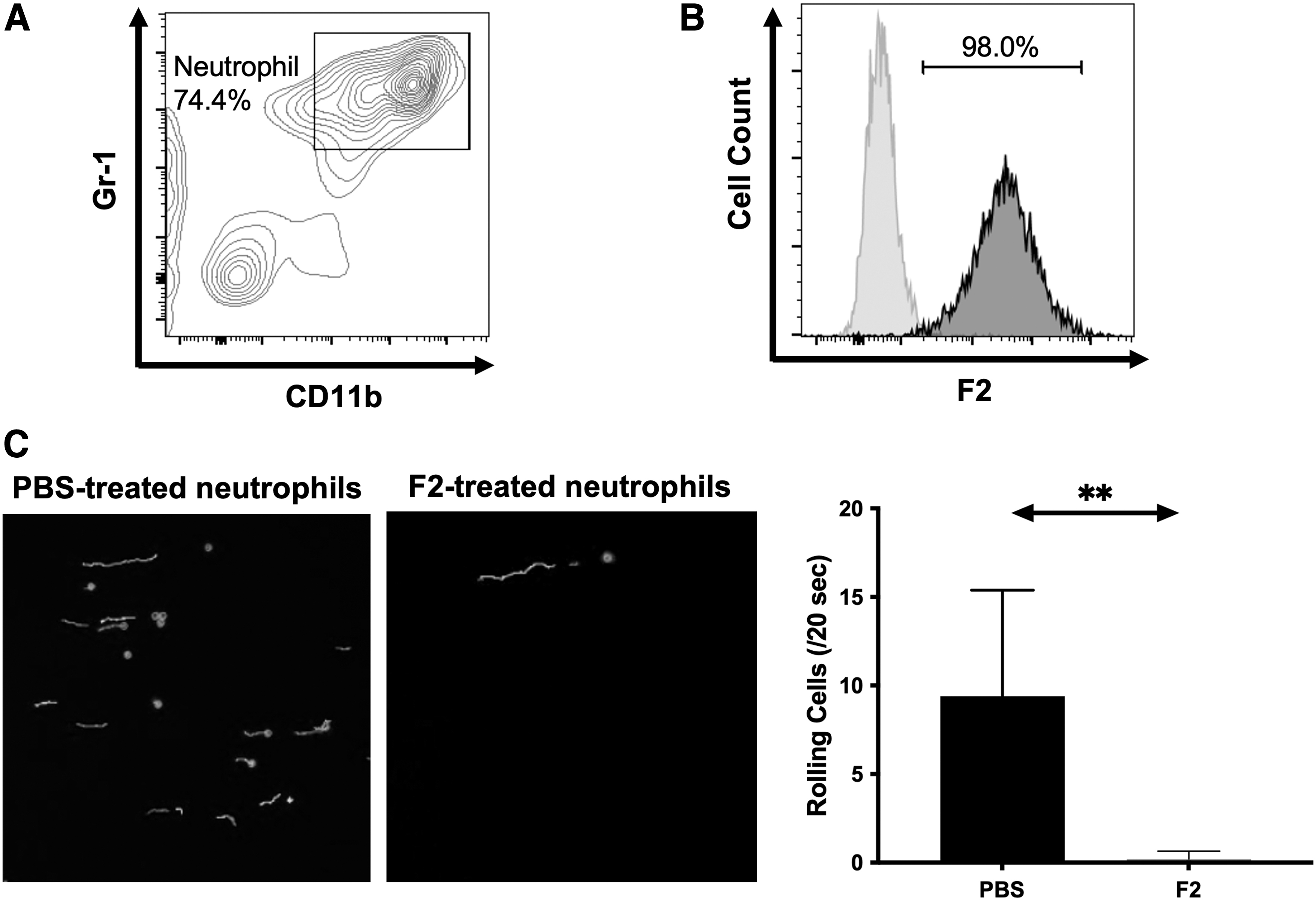
sLex mediates rolling of neutrophils on P-selectin-expressing cells.
Discussion
In this study, using Fut4/7 DKO mice lacking fucosyltransferase-IV and -VII, which are critical for the expression of sLex, it was found that sLex glycan had an important role in neutrophil infiltration in ALI. The therapeutic effects of a highly specific anti-sLex mAb F2 were also observed in the ALI model, which suggested that sLex glycans could serve as a novel therapeutic target to block ALI.
ALI has high morbidity and mortality rates worldwide, 23 and at present, for the majority of clinical treatments, an external ventilator is used. 24 In addition to ventilation therapy, nonventilation therapy options are available, and can be divided into agonist therapy, 25 reductase inhibitors, 26 and bone marrow mesenchymal stem cells. 27 However, the efficacy of these treatments remains controversial.
Neutrophils play an important role in the pathogenesis of ALI. In this study, using the sLex-specific mAb F2, the function of the sLex glycan on the surface of neutrophils was specifically blocked. Previous studies have shown that neutrophil elastase (NE) was influential on neutrophils as the pathogenic cells in ALI. 28 A major inflammatory protease known as NE is released by neutrophils. 29 NE inhibitors have been used to treat ALI in clinical trials, although the results of this have not been effective enough. 30 Considering that the mAb F2 significantly suppressed neutrophil infiltration in murine models of ALI, we believe that this mAb could serve as an alternative or additional therapeutic agent for blocking the rapid progression of ALI.
However, this notion cannot be applied directly in patients, since F2 is a mouse IgG1 mAb. The generation of humanized F2 would be the next important step in translating the results of the present study to clinics.
It is well known that COVID-19, a new type of pneumonia that has widely spread around the world since 2019, causes respiratory infections in humans. 31 This new type of respiratory disease has clinical manifestations similar to those of ARDS, 32 a life-threatening clinical complication of ALI. In this study, we found that the blocking sLex significantly suppressed neutrophil infiltration into the lungs following a cascade reaction with E- and P-selectin in ALI. We also found that inflammatory cytokine production in the lungs was suppressed through administration of mAb F2 targeting sLex in mice as an ALI model. Previous studies have demonstrated that neutrophils played an important role in the pathogenesis of COVID-19. 33
In clinical trials, the treatment of COVID-19 has been divided into antiviral drugs, continuous injection of plasma during the recovery period, and vaccine prophylaxis. 34 Since the infiltration of neutrophils into the lung is dependent on sLex, we believe that sLex could be a new therapeutic target against COVID-19 and furthermore, that mAb F2 would be useful for specific targeting of this glycan.
Conclusions
In this study, we show that the infiltration of neutrophils, the major inflammatory cells causing ALI, to the lung is mediated by sLex using Fut4/7 DKO mice lacking sLex expression. We also show that administration of an anti-sLex mAb F2 3 hours after LPS administration significantly suppressed pulmonary neutrophil infiltration, accompanied by the reduced induction of inflammatory cytokines. Consistent with this result, ex vivo cell-rolling assay indicated that mAb F2 efficiently blocked rolling of murine neutrophils on P-selectin-expressing cells. Collectively, the results of this study indicate that the sLex glycan could serve as a therapeutic target against ALI, and also that mAb F2 would be useful as a therapeutic agent against ALI.
Footnotes
Acknowledgments
We appreciate Drs. Jotaro Hirakawa and Hiroko Nakatsukasa for their technical support and helpful discussions.
Authors' Contributions
W.L.: conceptualization, methodology, investigation, and writing-original draft. W.X., W.L., and Z.W.: investigation. H.A.: supervision and writing-review and editing. H.K.: conceptualization, supervision, writing-review and editing, and funding acquisition.
Author Disclosure Statement
The authors declare that they have no conflicts of interest regarding the contents of this article.
Funding Information
This study was partly supported by AMED (22ae0121017h0002 and 22ym0126065h0001), and JSPS KAKENHI (20H03379).