
Abstract
Porcine transmissible gastroenteritis virus (TGEV) infection results in severe gastrointestinal disease manifesting vomiting, diarrhea in neonatal porcine, with extremely high mortality. Monoclonal antibody (MAb) specific to TGEV nonstructural protein (NSP)14 that contains two functional domains, exonuclease (ExoN) and methyltransferase (MTase) domains, may help elucidate the role of NSP14 in the viral life-cycle. In this study, we developed a murine MAb, designated 12F1, against TGEV NSP14 using traditional cell-fusion technique. It was shown the MAb can exclusively bind to viral NSP14, as evidenced by the results of indirect fluorescent assay and western blotting. Intriguingly, epitope screening assay shown that 12F1 targets a hinge region connecting ExoN and N7-MTase of NSP14.
Introduction
Porcine transmissible gastroenteritis manifests severe vomiting, diarrhea and dehydration in neonatal swine, with a mortality approaching 100%. 1 The etiologic agent, porcine transmissible gastroenteritis virus (TGEV), contains a single-stranded positive-sense genomic RNA, with the largest genome ∼30 kb among RNA viruses. This genome encodes 16 viral nonstructural proteins (NSPs), respectively, involved in viral replication, pathological damage, and immune evasion. 2 Among these NSPs, NSP14 acts as a catalytic subunit when activated by NSP10. The individual NSP14 simultaneously has two distinct functions, dsRNA exonuclease (ExoN) and guanosine 5′ methyltransferase (MTase) activities. 3
Several crucial amino-acid mutations in NSP14, which abolished proofreading function, significantly reduced viral adaption to host, thus suggesting the crucial importance of ExoN in coronavirus transcription and replication.4,5 In addition, reverse genetics study showed that ExoN inhibition enhanced coronavirus sensitivity to chain terminating and mutagenic nucleotides. 6 Meanwhile, the C-terminal (N7-guanine)-MTase domain confers immune evasion from recognition and degradation by host cell sensor. 7 Since such functional significance of NSP14/NSP10 complex, a variety of drug candidates targeting NSP14/NSP10 complex are being intensively developed for preventing various coronavirus infections.3,8
In this study, we developed a TGEV NSP14-specific monoclonal antibody (MAb), which targets a linking region between ExoN and MTase of NSP14. In addition, we mapped the precise epitope recognized by the MAb. This MAb deserves further investigation for antiviral development against TGEV.
Materials and Methods
Preparation of MAb against TGEV NSP14
The preparation of His-tagged TGEV NSP14 and its corresponding murine MAbs were described previously.9–11 In brief, 6-week-old female BALB/c mice were immunized subcutaneously with 100 μg of the TGEV NSP14 at 2-week intervals. Mice splenocytes were harvested and fused with SP2/0 using 50% polyethyleneglycol (Sigma-Aldrich, MO). Antibodies in culture supernatants were screened using enzyme-linked immunosorbent assay (ELISA).
The positive hybridoma cells were repeatedly cloned by a limited dilution, and the stable hybridoma clones were injected into abdominal cavities of BALB/c mice to yield sufficient antibody. Subsequently, the MAbs were harvested from the seroperitoneum and purified with an antibody purification kit according to the manufacturer's specifications (NAb™ Protein A/G Spin Kit, Thermo Scientific). The animal study was approved by the ethics committee of the Wuhan University of Bioengineering, China (permit no. WUB20220517).
Immunofluorescence assay
PK-15 cells were seeded into 24-well tissue culture plates (Costar Corning, Inc., NY) at a density of 1 × 105 cells per well. When the cells reached ∼70% confluence, culture medium was removed and then cells were washed three times with phosphate buffer saline (pH7.4) and incubated with TGEV (WH-01 strain) at a multiplicity of infection (MOI) of 0.01 for 1 hour at 37°C.
After removal of supernatant, fresh medium was added and cultures were incubated at 37°C. At 10 hours of postinfection, the infected cells were fixed with absolute methanol and incubated with MAbs for 2 hours, followed by application of fluorescein isocyanate-conjugated goat anti-mouse IgG. Fluorescent images were examined under a fluorescent microscope.
Immunoblotting
The TGEV-infected PK-15 cells were separated by 10% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then electrophoretically transferred to nitrocellulose transfer membrane (GE Healthcare). The membrane was blocked for 1 hour at room temperature with blocking solutions containing 1% bovine serum albumin in tris buffered saline (TBS) (20 mM Tris-HCl [pH7.5], 150 mM NaCl), and then incubated overnight with MAb at concentration of 1 μg/mL at 4°C.
After washing three times with T-TBS (20 mM Tris-HCl [pH7.5], 150 mM NaCl, 0.05% Tween 20), the membrane was incubated for 30 minutes with horseradish peroxidase-conjugated anti-mouse IgG (Jackson ImmunoResearch Laboratories, West Grove, PA). After washing three times with T-TBS, the membrane was developed by treatment with ECL Western Blotting Detection Reagents (GE Healthcare, Darmstadt, Germany).
Enzyme-linked immunosorbent assay
The peptides derived from the 19 amino acids of Hinge I was chemically synthesized (Moonbiochem, Wuhan, China). The epitope on NSP14 recognized by 12F1 was determined by peptide-based ELISA. Generally, 96-well ELISA plates were coated with 100 μL of the indicated antigen (2 μg/mL of peptides) in coating buffer at 4°C overnight.
MAb 12F1 and irrelative MAb 5G10 (a salmonella bacteria flagellin-specific MAb) were detected by using alkaline phosphatase-labeled goat anti-mouse IgG (SouthernBiotech, AL), followed by substrate p-nitrophenyl phosphate (Sigma-Aldrich, MO). The reciprocal values of the last dilution giving an OD405nm reading that was twice the background of the empty wells were determined as positive.
Results and Discussion
Preparation of TGEV NSP14
To obtain murine MAb specific for NSP14, we sought to construct and express full-length TGEV NSP14 in pET28a prokaryotic expression system (Novagen), but failed. Fortunately, a truncated construct harboring entire Exon and partial MTase (NSP14 201-460 amino acid, namely NSP14a) in BL21 was successfully expressed upon the induction of 0.5 mM isopropyl β-
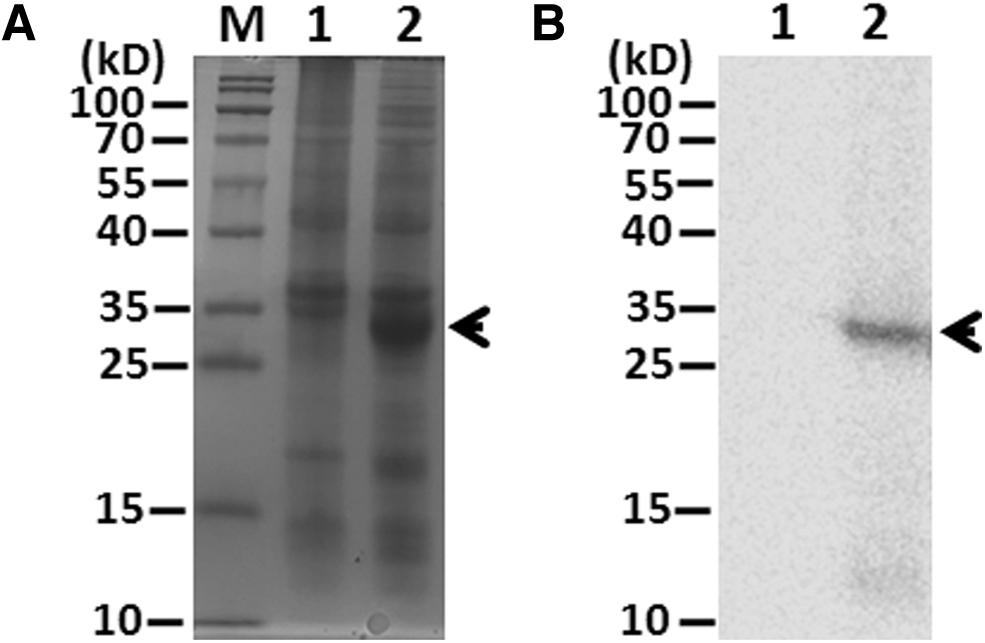
Expression of prokaryotic TGEV-NSP14a.
To further confirm whether the protein about 27 kDa mentioned earlier was NSP14a, we utilized anti-Histag (Proteintech, CN) to immunoblot with induced BL21 upon IPTG by western blotting (WB). As seen in Figure 1B, the induced BL21 containing pET28a-NSP14a plasmid generated a single band (∼27 kDa) (Lane 2), which was in agreement with the result of SDS-PAGE.
Subsequently, the induced target protein (NSP14a) was then purified using affinity chromatography Ni resin (GE) after mass culture of BL21 containing pET28a-NSP14a. Collectively, a truncated NSP14 (NSP14a) was produced with high purity. In turn, the NSP14a was used to immune five BALB/c mice.
Characterization of the MAb toward TGEV NSP14
The five BALB/c mice were immunized with purified NSP14a as described previously.9–11 The fusion hybridoma cell secreting specific antibody against NSP14a were selected by using ELISA and limited dilution approach. Finally, a stable positive hybridoma cell line, designated MAb 12F1, was established. The characteristics of 12F1 were determined by indirect fluorescent assay (IFA) and WB. PK-15 was infected by TGEV (WH-01 strain) at MOI 0.01 for 10 hours. The TGEV-infected cells were subject to IFA. As shown in Figure 2A, fluorescent signal can be seen when 12F1, but not 5G10 (a MAb against bacterial flagellin), 9 was applied as detection antibody.
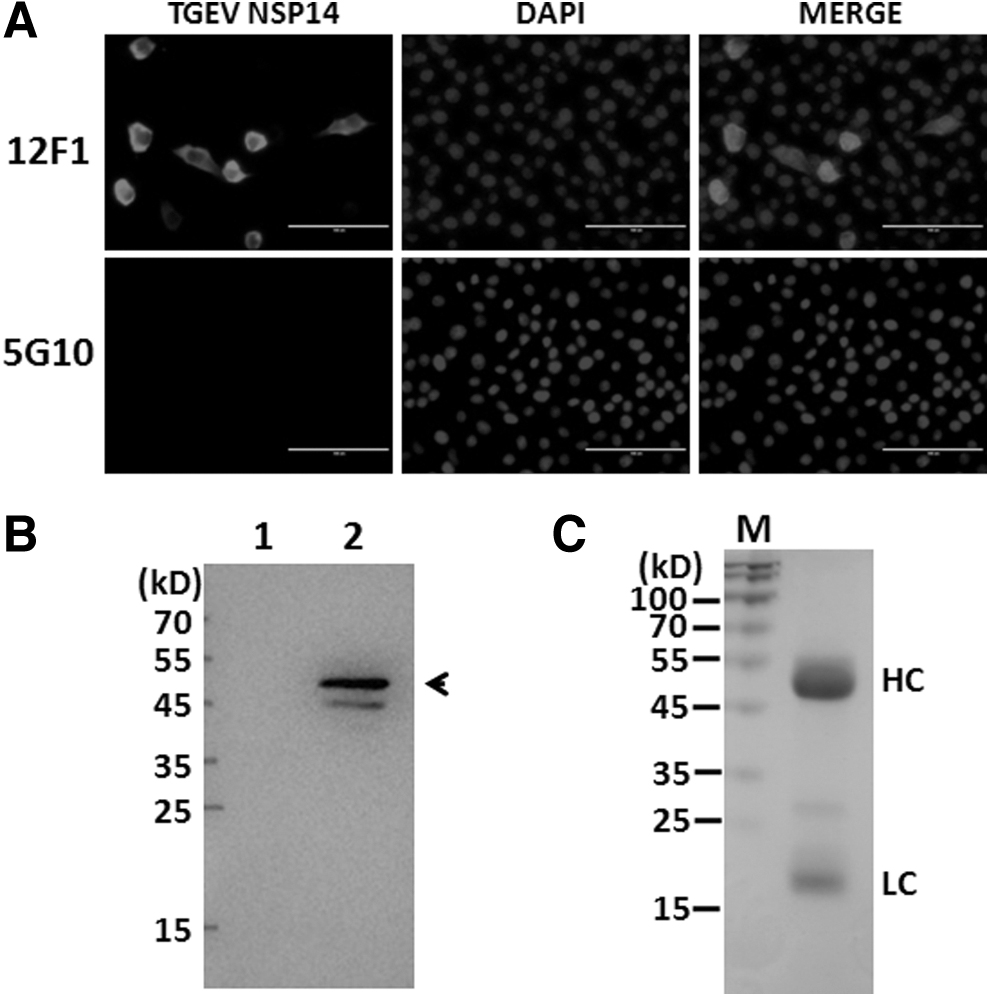
MAb 12F1 recognizes eukaryotic TGEV NSP14.
TGEV NSP14 is dominantly located in cytoplasm of PK-15 at 10 hours of postinfection. Moreover, the binding activity of 12F1 to NSP14 was evaluated by immunoblotting. As seen in Figure 2B, 12F1 can specifically interact with viral NSP14 (about 52 kDa). The purified MAb 12F1 was subject to SDS-PAGE, the result showed 12F1 have both typical heavy chain and light chain (Fig. 2C). ELISA result revealed that MAb 12F1 belongs to IgG1 subtype. (Data not shown) Therefore, the MAb 12F1 we developed here can specifically bind to viral NSP14.
Epitope determination of the MAb 12F1
The entire TGEV NSP14 consists of an Exon domain, two hinge regions, and two N7-MTase domains. (Fig. 3A) Next we further clarify the precise epitope recognized by 12F1. Three constructs derived from NSP14a with various truncations as indicated in Figure 3A. The BL21s containing various recombinant plasmids were induced by IPTG, subsequently the cell samples were subject to immunoblotting analysis by the aforementioned method.

12F1 recognizes a hydrophilic epitope of TGEV-NSP14.
As shown in Figure 3B, three truncates, along with NSP14a, show obvious signal, demonstrating the binding capacity of MAb 12F1 with all four various constructs derived from NSP14. Thus, the four NSP14 truncates shared the epitope of 12F1. Based on these results, the epitope recognized by MAb 12F1, therefore, should be set in the first hinge region (hinge I, 282IHDCFVKRVDWSIVYPFID300) of NSP14.
To accurately identify the epitope of 12F1, the four overlapping peptides derived from hinge I region were synthesized as indicated in Figure 3C. Each peptide was coated on the 96-well ELISA plate, respectively, to assess the binding activity of 12F1 by ELISA. The irrelative MAb 5G10 could not associate with any indicated peptide. As observed in Figure 3D, 12F1 had the strongest binding capacity with peptide 3 (P3, 292WSIVYPFID300), followed by P3 (286FVKRVDWSI294). Therefore, the P3 (292WSIVYPFID300) was the core epitope recognized by 12F1.
To further determine the spatial position of the epitope of 12F1 on NSP14, the peptide was localized on the model using PyMOL. Specifically, the peptide (292WSIVYPFID300) was fixed in the PyMOL molecule model (PDB: 7R2V). 12 As shown in Figure 3E, the epitope of 12F1 was a hinge region between N7-MTase and ExoN, and exposed on the surface of the molecule model, suggesting its hydrophilic property.
A proper interface between MTase and polymerase of flavivirus plays important role in RNA initiation and elongation. 13 The MAb 12F1 developed in this study exclusively targets a hinge region between N7-MTase and ExoN, which orchestrate the TGEV genome RNA replication. Therefore, the application of 12F1 may perturb in the interface of N7-MTase and ExoN and disturb the TGEV replication. In addition, 12F1 associating with NSP14 probably interfere the interaction of NSP10 and NSP14 through steric hindrance effect, thereby inhibiting virus RNA replication. Therefore, a genetically modified 12F1 warrants further investigation for its potential application.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Natural Science Foundation of China (No. 31972692), Hubei Provincial Natural Science Foundation of China (2020CFB520) and High-level Scientific Research Foundation for the introduction of talent of Wuhan University of Bioengineering (2017KQ01).