
Abstract
CD300A is a member of the CD300 immunoglobulin (Ig)-like receptor family consisting of eight molecules in humans, all of which contain one Ig-like domain in the extracellular portion. Upon binding its ligand phosphatidylserine or phosphatidylethanolamine, CD300A mediates an inhibitory signal through the immunoreceptor tyrosine-based inhibitory motif in the cytoplasmic portion. The CD300 family molecules are highly homologous to each other. In addition, CD300A has a single nucleotide polymorphism (rs2272111), which is a nonsense mutation encoding glutamine (CD300AQ111) instead of arginine (CD300AR111) at residue 111 in the Ig-like domain of CD300A. In this study, we successfully generated monoclonal antibodies (mAbs) specific to either CD300AR111 or CD300AQ111 or both. These mAbs are useful for the analysis of CD300A genotype by flow cytometry and the development of an antibody drug for the treatment of various diseases.
Introduction
The CD300 family molecules are encoded by 8 genes mapped on chromosome 17 in humans and 12 genes on chromosome 11 in mice. They belong to the immunoglobulin (Ig) gene superfamily and contain one Ig-like domain in the extracellular portion. They are preferentially expressed on myeloid cells and mediate either activating or inhibitory signals through associated adaptor proteins, such as DAP 12 or FcɛRIγ, and immunoreceptor tyrosine-based inhibitory motif (ITIM) in the cytoplasmic region, respectively.1,2
CD300a and CD300A, one of the CD300 family members in mice and in humans, respectively, have a long cytoplasmic region containing ITIM, which provides a docking site for Src homology 2 (SH2)-containing cytoplasmic phosphatases, including SHP1, SHP2, and SHIP1,3 and mediates an inhibitory signal in myeloid cells upon binding to the ligands phosphatidylserine and phosphatidylethanolamine (PE).4,5 We previously reported that CD300a suppresses innate immune responses by mast cells, dendritic cells, and macrophages in mice.6–8 Moreover, blockade of CD300a signaling ameliorated the pathogenesis of allergies, including passive systemic anaphylaxis and airway inflammation, sepsis, enteritis, tumor formation, and acute ischemic stroke in mouse models.9–11 These results suggested that a neutralizing anti-CD300A monoclonal antibody (mAb) may be useful for the treatment of these diseases. However, CD300A has a single nucleotide polymorphism (SNP) (rs2272111), which is a nonsense mutation encoding glutamine (CD300AQ111) instead of arginine (CD300AR111) at residue 111 in the Ig-like domain of CD300A.
The CD300 family molecules show high homology with each other. Among them, CD300A shows the highest homology with CD300C. Thus, the anti-CD300A mAb should specifically recognize CD300A but not other CD300 family molecules. In addition, mAbs specific to either CD300AQ111 or CD300AR111, but not to other CD300 family molecules, would also be useful for the analysis of CD300A genotype by flow cytometry. Therefore, anti-CD300A mAbs that specifically recognize either CD300AQ111 or CD300AR111, but not CD300C, would be useful for the analysis of CD300A genotype by flow cytometry. In addition, anti-CD300A mAbs specific to both CD300AQ111 and CD300AR111, but not CD300C, are required for the blockade of CD300A-mediated signaling as an antibody drug candidate. In this study, we successfully generated antibodies specific to either CD300AR111 or CD300AQ111 or both but not to CD300C.
Materials and Methods
Generation of the Fc proteins and establishment of transfectants
The human CD300AR111 and CD300AQ111 extracellular portions were fused with the human IgG Fc portion at the C-terminus and subcloned into the pcDNA3.4 vector for the generation of CD300A-Fc proteins. For the generation of transfectants, the human CD300AR111 and CD300AQ111 cDNA tagged with IRES-GFP was subcloned into the pcDNA3.4 vector. Then BW5147 expressing CD300AR111-IRES-GFP and CD300AQ111-IRES-GFP were generated as described. 12 Human CD300C-IRES-GFP expressing transfectant was used as previous study. 13
Establishment of hybridomas
Balb/c mice were immunized with the human CD300AR111-Fc or CD300AQ111-Fc proteins emulsified with the complete Freund's adjuvant (Sigma-Aldrich) by intraperitoneal or subcutaneous injection. After 1, 2, and 3 weeks, mice were challenged with the human CD300AR111 and/or CD300AQ111-Fc proteins emulsified with the incomplete Freund's adjuvant (Sigma-Aldrich). Cells from the spleen or draining lymph nodes were harvested 3 days after the challenge, fused with SP2/O myeloma cells, and cultured in HY Medium C (Stem Cell Technologies) overnight. Then cells were cultured in HAT medium in a 96-well plate for 7 days and overlayed by Medium E for antibody production (Stem Cell Technologies). The supernatants were harvested and analyzed by flow cytometry for screening.
Antibodies and flow cytometry
Cells were incubated with a human FcR blocking reagent (Miltenyi) in a staining medium (2% fetal bovine serum and 0.01% sodium azide in phosphate buffered saline) for 15 minutes to mask the Fcγ receptors. Cells were then stained with indicated antibodies for 30 minutes on ice. All samples were analyzed by using the LSRFortessa flow cytometer (Becton Dickinson), followed by analyses using FlowJo software (Tree Star). Fluorochrome-conjugated mAbs specific for, CD16-PE (3G8), CD56-V450 (B159), CD19-FITC (HIB19), CD4-FITC (L200), CD3-PE (HIT3a), and CD4-V500 (RPA-T4) were purchased from BD Biosciences. Anti-human CD14-FITC (MHCD1401) was purchased from Invitrogen and CD8-FITC (RPA-T8) was purchased from BioLegend. Mouse IgG2b (MPC-11) and APC-conjugated anti-mouse IgG2b (RMG2b-1) mAbs were purchased from abcam and APC-conjugated anti-human CD14 (Tuk4) was purchased from Thermo Fisher. Dead cells were excluded by using propidium iodide. Doublets were excluded by gating out the population defined by forward and side scatters by the flow cytometry in all the experiments.
Cell preparation
Human and Macaca fascicularis mononuclear cells were isolated from the peripheral blood by density separation using Lymphoprep (Serumwerk) and Lympholyte-Mammal (Cedarlane), respectively. IRB approval for CD300A analysis in human peripheral blood was undertaken.
Results and Discussion
We found that the extracellular portions of CD300AR111 and CD300AQ111 showed 81% and 80% homologies with that of CD300C, respectively (Fig. 1A). We established the BW5147 transfectants stably expressing CD300AR111, CD300AQ111, or CD300C (CD300AR/BW, CD300AQ/BW, or CD300C/BW, respectively). We found that TX49 clone, which was previously generated in our laboratory, 4 recognizes all CD300AR111, CD300AQ111, and CD300C (Fig. 1B). To establish mAbs specific to either human CD300AR111 or CD300AQ111 or both, but not to CD300C, we generated chimeric fusion proteins of the extracellular portion of CD300AR111 or CD300AQ111 with the Fc portion of human IgG (CD300AR111–Fc and CD300AQ111–Fc, respectively). We immunized mice with either CD300AR111-Fc, CD300AQ111-Fc, or both and obtained hybridomas secreting antibodies against CD300A. We successfully obtained mAbs TX112 and TX113, TX114, and EX67, which specifically recognize both CD300AR111 and CD300AQ111, CD300AR111 alone, and CD300AQ111 alone, respectively. However, all these clones did not bind to CD300C (Fig. 1B).
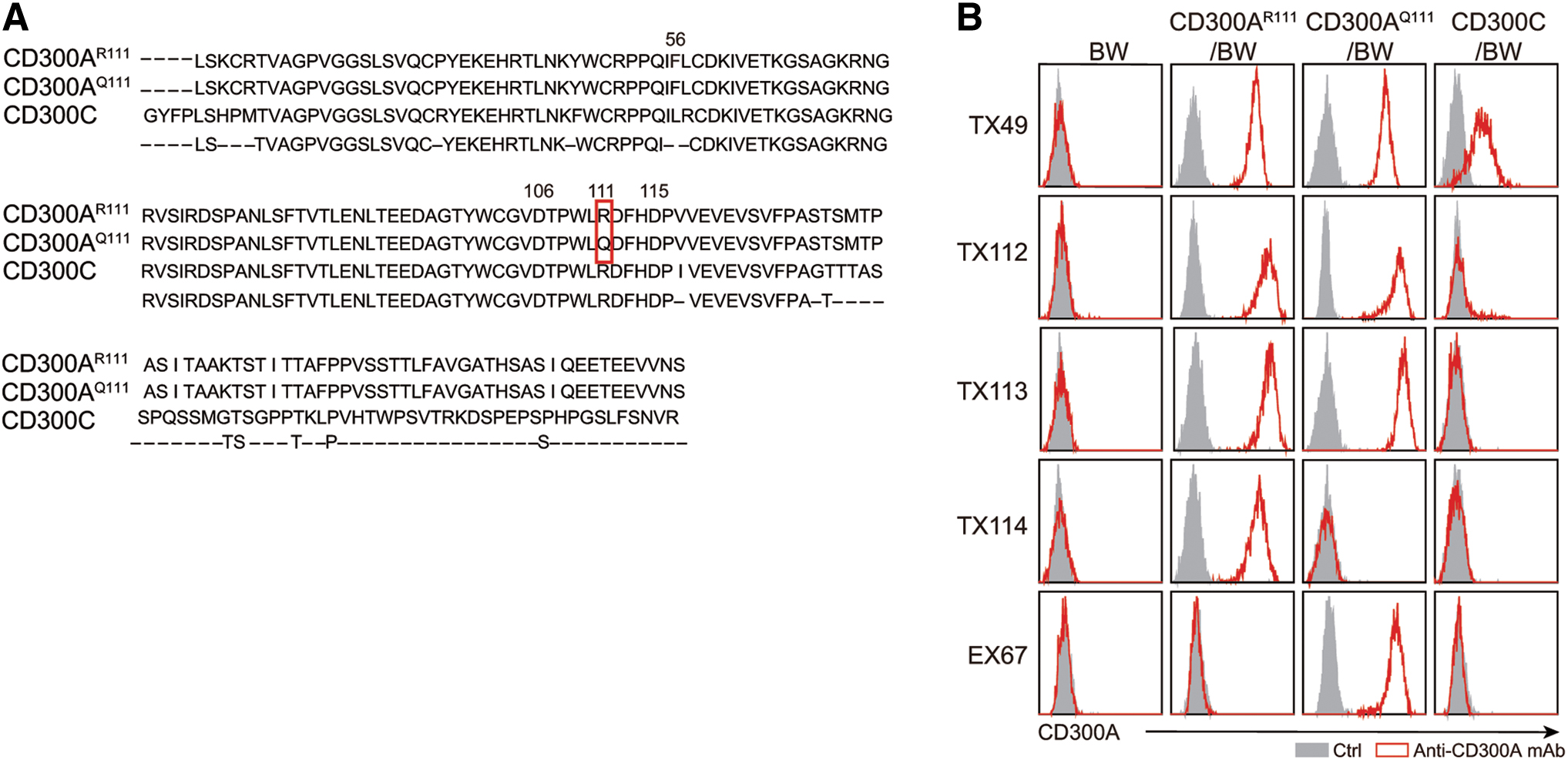
Generation of anti-CD300A mAbs.
Flow cytometry analysis using TX112 mAb, which recognizes both CD300AR111 and CD300AQ111, but not CD300C, demonstrated that CD300A was expressed on all the subpopulations of monocytes in the human peripheral blood. CD300a expression was also detected on a subpopulation of B cells and CD16-CD56hi NK cell population (Fig. 2A–C), consistent with previous reports.12–15
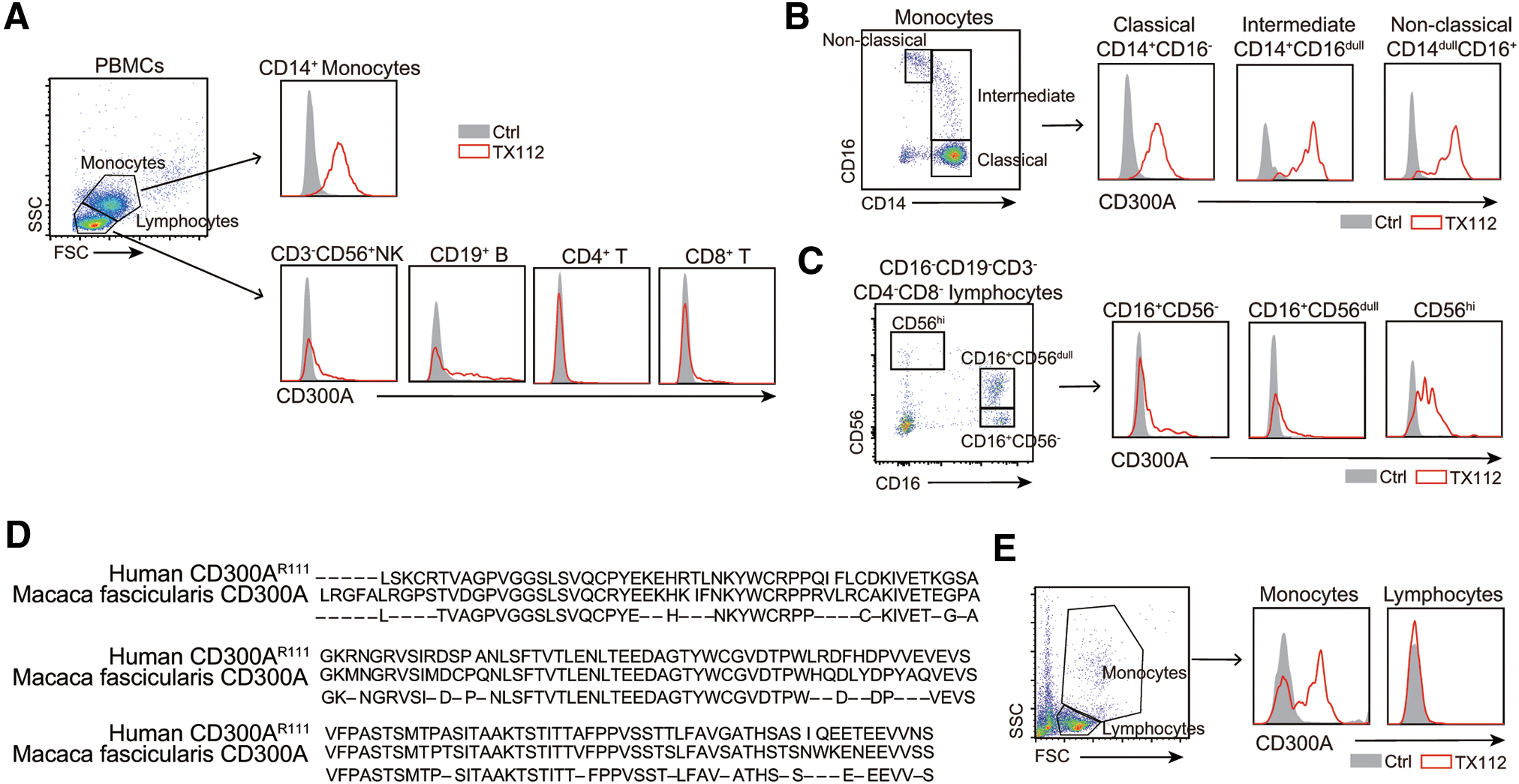
Expression of CD300A in humans and Macaca fascicularis.
The extracellular portion of CD300A of Macaca fascicularis shows 78% homology to that of human CD300A (Fig. 2D). We found that TX112 mAb cross-reacted to Macaca fascicularis CD300A (Fig. 2E).
A genetic linkage study demonstrated that the SNP in the Ig domain of CD300A (rs2272111), which revealed 20%–30% frequency of the population, may link to psoriasis susceptibility. 16 In this study, we successfully generated TX114 and EX67 mAbs that distinguish CD300AR111 from CD300AQ111 and vice versa. These mAbs may be useful for the analysis of CD300A genotype by flow cytometry. Moreover, TX112 and TX113 recognize both CD300AR111 to CD300AQ111, but not CD300C. These mAbs may be useful for the development of antibody drugs because these mAbs can bind CD300A of any genotypes of patients, although further functional analyses of these mAbs should be required.
Footnotes
Acknowledgments
We thank H. Furugen and M. Kaneko for their secretarial assistance.
Authors' Contributions
C.N-O., H.K., W.L. and Y.Y-K. designed and performed research and analyzed data. C.N-O., H.K., W.L. and A.S. wrote the manuscript. K.T., K.S. and A.S. validated and supervised the overall project. All authors critically reviewed the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by grants provided by the JSPS KAKENHI grant numbers 18H05022 and 21H04836 (to A.S.) and 22H03170 (to C.N-.O.) and Takeda Science Foundation (to C.N-O).