
Abstract
Coagulation factor XIII (FXIII) is an enzyme that strengthens hemostatic clots, and its deficiency can cause life-threatening bleeding. We immunized mice with human plasma-derived FXIII to generate monoclonal antibodies (mAbs) against the B subunit (FXIII-B), which stabilizes the A subunit (FXIII-A) of FXIII, and analyzed their properties. The epitopes of the seven mouse antihuman FXIII-B mAbs obtained were found to be the 3rd, 5th, 6th, 9th, and 10th Sushi domains. One of these mAbs, mAb 5–6C, recognized the 10th Sushi domain and inhibited the fibrin cross-linking reaction without affecting the amine incorporation activity of FXIII. We previously reported that the 10th Sushi domain is the site where FXIII-B binds to fibrin and functions to bring FXIII-A closer to the substrate fibrin. Except for mAb 5–6C, mouse mAbs with high yields were used to measure the amount of FXIII-B antigen by an immunochromatography test (ICT), which showed a high correlation with enzyme-linked immunosorbent assay–obtained results. In addition, we developed a prototype ICT to detect anti-FXIII-B autoantibodies using mAb 1–3C, which showed good results in measuring the amount of FXIII-B antigen. Thus, mouse mAbs may be useful for clinical applications. mAb 5–6C targeting the 10th Sushi domain may also be useful for inhibiting thrombosis progression when humanized as antibody medicines.
Introduction
Coagulation factor XIII (FXIII) is a precursor to a fibrin-stabilizing factor, also known as plasma protransglutaminase, composed of an A subunit (FXIII-A) dimer and a B subunit (FXIII-B) dimer, which together form an A2B2 heterotetramer in plasma.1–3 At the final stage of blood clotting, thrombin activates FXIII, converting it into activated FXIII (FXIIIa), a plasma transglutaminase. This enzyme cross-links fibrin monomers, fibrin and α2-plasmin inhibitor (the primary fibrinolytic inhibitor in plasma), thus increasing the mechanical strength and resistance to proteolysis of the clot.
FXIII-A contains the enzyme-active site, while FXIII-B consists of 10 tandem Sushi domains,4,5 which are defined as follows: Protein modules (complement control protein [CCP] modules, short consensus repeats [SCR], ∼60 amino acids) that structurally are said to resemble Japanese sushi. They are involved in protein–protein interactions and are common in many proteins involved in the regulation of the complement system, blood coagulation, cell surface proteins (IL2 and IL15 receptors), and some selectins. 6
FXIII-B stabilizes and protects FXIII-A from rapid clearance from the bloodstream. 7 Consequently, a significant deficiency in either FXIII-A or FXIII-B, whether congenital or acquired, results in life-threatening bleeding. Although rare, with an annual incidence of 0.044 per million people, 8 autoantibodies can develop against FXIII-A and/or FXIII-B, primarily in older individuals, leading to autoimmune-acquired FXIII deficiency (AiF13D). Therefore, rapid assessment of FXIII activity, measurement of the antigen levels of FXIII-A and FXIII-B, and identification of autoantibodies against FXIII-A or FXIII-B are crucial for a definite diagnosis of AiF13D, as recommended by Japanese and international guidelines,9,10 to prevent early hemorrhagic death. 11
This study aimed to develop and characterize monoclonal antibodies (mAbs) against FXIII-B in mice. The developed mAbs were preliminarily utilized in rapid point-of-care and immunochromatographic tests (ICT) to quantify the FXIII-B antigen and detect anti-FXIII-B autoantibodies.
Materials and Methods
Statement of Ethics
This study was approved by the Institutional Review Board of the Yamagata University School of Medicine. All procedures were conducted in accordance with the Declaration of Helsinki. Informed consent was obtained from all individuals.
Materials
Recombinant FXIII-B (rFXIII-B) and its truncated constructs were produced using a baculovirus system and purified as previously described. 12 Anti-FXIII-B polyclonal antibody (pAb) was procured from Calbiochem. Its immunoglobulin G (IgG) was purified via Protein A-Sepharose (GE Healthcare Bioscience AB) and biotinylated using the ECL Protein Biotinylation Module. Horseradish peroxidase (HRP)-conjugated antimouse IgG, HRP-streptavidin, Protein G-Sepharose, and CNBr-activated Sepharose 4B were also sourced from GE Healthcare Bioscience AB. Protein A-coated Staphylococcus aureus (PANSORBIN) was obtained from Merck KGaA. The tetramethylbenzidine (TMB) Peroxidase Substrate Kit was purchased from Bio-Rad Laboratories. Immobilon Western Chemiluminescent HRP Substrate was supplied by Millipore. Ready-to-GlowTM-secreted luciferase was obtained from Clontech. Coagtrol N, a standard normal plasma, was purchased from Sysmex Corporation. N,N-dimethylcasein, monodansylcadaverine (MDC), and bovine plasma thrombin were obtained from Sigma-Aldrich.
Generation of mouse mAbs against human FXIII-B
Six-week-old female BALB/c mice were immunized subcutaneously every 2 weeks, three times, with purified human plasma-derived FXIII (A2B2 heterotetramer) mixed with complete Freund’s adjuvant. Four days after the final intraperitoneal injection of FXIII, splenic cells from the mice were fused with NS-1 myeloma cells. Hybridomas producing significant amounts of anti-FXIII mAbs were identified based on their binding affinity to purified FXIII using a sandwich enzyme-linked immunosorbent assay (ELISA) with a rabbit antimouse γ-globulin antibody. Selected hybridoma cell lines underwent several rounds of cloning by limiting dilution. The resultant cell lines were cultured, and the mAbs were purified from the culture supernatants via 40% ammonium sulfate precipitation and gel filtration with a Sephacryl S-200 column.
The hybridoma supernatants were further tested for reactivity with rFXIII-B using ELISA. Positive clones were subcloned and re-evaluated by ELISA and immunoblotting.
ELISA for binding of mAbs to rFXIII-B
A sandwich ELISA was performed as previously described. 13 A microtiter plate coated with anti-FXIII-B pAb IgG served as the capture antibody to detect FXIII-B. Three concentrations (1, 5, and 25 ng) of rFXIII-B were added and incubated. This was followed by incubation with seven homemade mouse mAbs against FXIII-B (1-3B, 1-3C, 3-3D, 4-2B, 5-6C, 5-11B, 6-5F; 0.5 mg/mL each) and three homemade mouse mAbs against FXIII-A as negative controls (1TH2-8C4C, 1TH6-2H7F, and 1TH6-10E 14 ; 0.5 mg/mL each). Subsequently, the plates were incubated with HRP-conjugated antimouse IgG as the detection antibody. TMB substrate was then added, and the reaction was stopped by adding 0.5 M sulfuric acid (H2SO4). Absorbance at 450 nm was measured using a Biolumin 960 microtiter plate reader (Molecular Dynamics) and compared with standard curves generated using rFXIII-B. To determine FXIII-B antigen levels in plasma, these levels were converted to equivalent values based on pooled normal plasma (a mixture of plasma derived from the plasma of 27 healthy individuals) used as a standard. Plasma levels are reported as a percentage of normal levels.
Western blotting for binding of mAbs to rFXIII-B
To analyze the binding of anti-FXIII-B to rFXIII-B, both full-length or truncated versions of rFXIII-B containing various Sushi domains were used. These were boiled in a solution containing 1% sodium dodecyl sulfate (SDS), 50 mM Tris-HCl (pH 6.8), 7.5% glycerol, and 0.01% bromophenol blue and then separated using SDS-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were transferred to nitrocellulose membranes, blocked, and incubated with affinity-purified mouse antihuman FXIII-B antibody. FXIII-B antigens were detected using an HRP-conjugated antimouse IgG and chemiluminescent substrate.
Binding of anti-FXIII-B mAbs to FXIII-B’s Sushi domains fused with Metridia luciferase
Metridia luciferase (MetLuc) fused with each FXIII-B Sushi domain (MetLuc-BSs) was prepared as described. 15 The fusion proteins were incubated with mAbs at 37°C for 2 hours, followed by incubation with IgG using protein A-coupled Sepharose at 4°C for 1 hour. The precipitate was washed twice with tris-buffered saline (TBS; 20 mM Tris-HCI, pH 7.5, 150 mM NaCl) with 0.1% Tween® 20 Detergent (TBS-T). MetLuc activity in the immunoprecipitate (IP) was measured using a Ready-to-Glow-secreted luciferase substrate. mAb binding was expressed as the ratio of the IP to the initial MetLuc-BS preparation (total).
Epitope analysis using FXIII-B peptide-coated plates
Microtiter plates were coated with 31 synthesized FXIII-B peptides (2.5 nmol; Supplementary Table S1) in 100 μL of 50 mM carbonate buffer (pH 9.6) and incubated overnight (12 hours) at 4°C. The plates were then incubated with homemade mouse mAbs (100–550 ng) at 4°C for 8 hours, followed by HRP-conjugated antimouse IgG at 37°C for 1 hour. The TMB substrate reaction and the termination were performed as described above.
Five-step dilution cross-mixing test for possible inhibition of FXIII activity using amine incorporation assay
To perform the assay, ∼10–50 μL of pooled normal plasma was mixed with 40–0 μL of culture medium containing mouse mAb clones (total 50 μL), followed by a 2-hour incubation at 37°C. Before starting the assay, a reagent solution was prepared by combining 1.5 mL of TBS, 6 mL of 0.4% N,N-dimethylcasein (final concentration 0.2%), 3 mL 5 mM MDC (final concentration 1.25 mM), 120 μL of 1 M dithiothreitol (final concentration 2 mM), 60 μL of 1 M CaCl2 (final concentration 5 mM), and 60 μL of 1000 U/mL thrombin (final concentration 10 U/mL).
For the test, 10 μL of normal plasma and mAbs mixture was combined with 90 μL of the premixed reagent solution. After 30 minutes of incubation at 37°C, the reaction was stopped with 0.1 mL of 10% trichloroacetic acid. The resulting precipitate was centrifuged at 12,000 rpm for 5 minutes, washed three times with 0.3 mL of a 1:1 mixture, and dissolved in 0.3 mL of 8 M urea, 1% SDS, and 50 mM Tris (pH 8.0). Fluorescence was then measured at 520 nm with an excitation wavelength of 360 nm.
Effect on fibrin cross-linking reaction in normal plasma
A total of 10 μL of pooled plasma was incubated with 10 μL of culture medium containing one of seven mouse mAbs at 37°C for 2 hours. Prior to the assay, a reagent solution was prepared by mixing 120 μL of TBS, 2.5 μL of 1 M CaCl2 (final concentration 6.7 mM), and 2.5 μL of 1000 U/mL thrombin (final concentration 6.7 U/mL). For the reaction, 10 μL of normal plasma and mAb mixture was combined with 10 μL of the reagent solution and incubated at 37°C for varying durations (0.5, 2, 5, or 10 minutes). The resulting clot was then harvested and analyzed using SDS-PAGE under reducing conditions with 5% β-mercaptoethanol.
ICT for quantitation of FXIII-B antigen
Each of the seven homemade mouse monoclonal antibodies was applied to a nitrocellulose strip (test line). rFXIII-B samples at concentrations of 1, 10, and 100 ng were added to a 96-well microtiter plate. The strip was then inserted into a well for 5 minutes at 37°C and washed three times. Subsequently, the strip was immersed in a solution containing a second homemade mouse antihuman FXIII-B mAb-gold conjugate for 15 minutes at 37°C. The results obtained were read using a reader device (FactScan, Denken Co., Ltd.) and expressed as a percentage of the pooled normal plasma standard.
Anti-FXIII-B autoantibodies detection using ICT
ICT was performed as previously described. 14 A homemade mouse monoclonal antibody was applied to a nitrocellulose strip to serve as the test line. Equal volumes of patient plasma and commercially available normal plasma were combined and incubated at 37°C for 5 minutes before performing ICT. Patient or control plasma premixed in a 1:5 dilution was added to a 96-well microtiter plate. The strip was then inserted into the well at 37°C for 5 minutes. After washing, the strip was placed into a solution containing another mouse mAb-gold conjugate at 37°C for 10 minutes. The results were read using a FactScan reader (Denken Co., Ltd.) and expressed as absorbance units relative to the positive control. The positive control, designated as 1.0 arbitrary unit, was prepared by mixing patient plasma with that of a congenital FXIII-B deficiency, which contained anti-FXIII-B alloantibodies 16 with Coagtrol N at 1:29 ratio and incubated at 37°C for 2 hours.
Statistical analysis
ELISA and FXIII amine incorporation (AI) assay were performed in triplicate for each sample. Statistical analyses were performed using JMP version 12.2.0 software (SAS Institute), and data were presented as mean ± standard deviation. Differences were considered significant at p-value < 0.05.
Results and Discussion
Characterization of mouse mAbs against human FXIII-B
We began by determining the concentration of eight homemade mouse mAbs in the culture medium through dot blot analysis (Supplementary Fig. S1). Using mouse IgG as a standard protein (at concentrations of 1, 10, and 100 ng), the concentration of each mouse mAb was estimated as follows: 1-3B (107 ng/μL), 1-3C (50 ng/μL), 3-3D (16 ng/μL), 4-2B (55 ng/μL), 4-8B (<1 ng/μL), 5-6C (196 ng/μL), 5-11B (48 ng/μL), and 6-5F (217 ng/μL). Due to its low yield, mAb 4-8B was excluded from further analysis.
Next, we characterized the remaining seven mAbs to aid the future development of an ELISA (or ICT or latex agglutination test) for quantifying the FXIII-B antigen. All seven mAbs bound to rFXIII-B in a dose-dependent manner (Supplementary Fig. S2). In contrast, none of the three anti-FXIII-A mouse mAbs 14 bound to rFXIII-B.
Epitope mapping of mouse mAbs against human FXIII-B using truncated rFXIII-B
To identify the regions recognized by the mAbs, Western blot analyses were conducted on full-length or truncated rFXIII-B (Supplementary Fig. S3, 1st round). Coomassie staining revealed one or two bands corresponding to constructs missing various Sushi domains (Fig. 1). For instance, mAb 1-3B bound to Lane 1; full-length rS1–10 (rFXIII-B include all 10 Sushi domains) + rS4–10 (7 Sushi domains without S1–3), Lane 2; rS2–10 (9 Sushi domains without S1) + rS1–6 (6 Sushi domains without S7–10), Lane 3; rS1–9 (9 Sushi domains without S10), Lane 4; rS1–8 (8 Sushi domains without S9 and S10), but not Lane 3; rS1–5 (5 Sushi domains without S6–10) and Lane 4; rS1–4 (4 Sushi domains without S5–10), suggesting that mAb 1-3B recognizes the 6th Sushi domain. All constructs were recognized by a control anti-FXIII-B antibody, indicating recognition by this “polyclonal” antibody.

Epitope mapping of mouse mAbs against human FXIII-B by Western blotting (1st round). To determine which Sushi domain(s) of human FXIII-B each mouse mAb interacts with, Western blotting analysis was conducted using two forms of recombinant FXIII-B, each containing various Sushi domains. The lanes were as follows: Lane 1; full-length rS1–10 (rFXIII-B containing all 10 Sushi domains) + rS4–10 (7 Sushi domains without S1–3), Lane 2; rS2–10 (9 Sushi domains without S1) + rS1–6 (6 Sushi domains without S7–10), Lane 3; rS1–9 (9 Sushi domains without S10) + rS1-5 (5 Sushi domains without S6–10), Lane 4; rS1–8 (8 Sushi domains without S9 and S10), rS1–4 (4 Sushi domains without S5–10). The table at the bottom summarizes the possible epitope Sushi domain for the mAbs against FXIII-B. FXIII-B, coagulation factor XIII B subunit; mAb, monoclonal antibody; NC, negative control.
To further pinpoint the specific Sushi domain(s) of human FXIII-B that each mouse mAb binds, immunoprecipitation analyses were conducted using truncated rFXIII-B (BSs) containing one or two consecutive Sushi domains fused with MetLuc (Supplementary Fig. S3, 2nd round). For example, mAb 1-3B is bound to BS5-6 and BS6 but not BS5, indicating that mAb 1-3B recognizes the 6th Sushi domain (Fig. 2). The FXIII molecule is illustrated based on the “FXIII-A dimer-two monomeric FXIII-B (A2[2B])” model. 17

Epitope mapping of mouse mAbs against human FXIII-B by immunoprecipitation (IP) assay (2nd round). To determine the specific Sushi domain(s) of human FXIII-B that each mouse mAb interacts with, binding analysis was conducted using truncated rFXIII-Bs (BSs) containing individual (black bars) or pair of consecutive Sushi domains (gray bars) fused with MetLuc. Bound mAbs were collected by precipitation with protein A-coupled Sepharose (PANSORBIN). The MetLuc activity of the IP was calculated as a ratio to that of the initial MetLuc-BS preparation (total). The binding sites of the mAbs against FXIII-B are illustrated in the schematic diagram at the bottom. The FXIII molecule is illustrated based on the “FXIII-A dimer-two monomeric FXIII-B (A2[2B])” model. 17 FXIII-B, coagulation factor XIII B subunit; mAb, monoclonal antibody.
Epitope analysis of each mouse mAb using 31 FXIII-B synthesized peptides
To pinpoint the epitopes of each mouse mAb, we used 31 synthesized peptides representing 83% (530/641 amino acids 4 ) of the FXIII-B molecule (Supplementary Fig. S4, Supplementary Table S1). Each mAb binds to one or two FXIII-B peptides on the plates, producing strong ELISA signals over 0.05 (absorbance at 450 nm, Fig. 3). Specifically, mAb 1-3C interacted with synthetic peptide B7 of the 3rd Sushi domain; mAbs 4-2B and 5-11B bound to peptides B11 and B12 of the 5th Sushi domain; mAb 1-3B recognized peptides B14 and B17 of the 6th Sushi domain; mAb 6-5F interacted with peptides B22 and B23 of the 9th Sushi domain; and mAb 5-6C bound to peptides B27 and B28 of the 10th Sushi domain. These results align with the Sushi domain epitopes identified through Western blot analyses, except for mAb 1-3C. The reaction with synthetic peptides B11 and B12 was significantly weaker than that with B7, suggesting that mAb 1-3C’s epitope might be in the 3rd Sushi domain rather than the 5th. Alternatively, the epitopes of both Sushi domains might share similar local immunogenic structures, or B7 of the 3rd Sushi domain and B11 and B12 of the 5th Sushi domain could be closely located and form a single “structural/conformational” epitope.
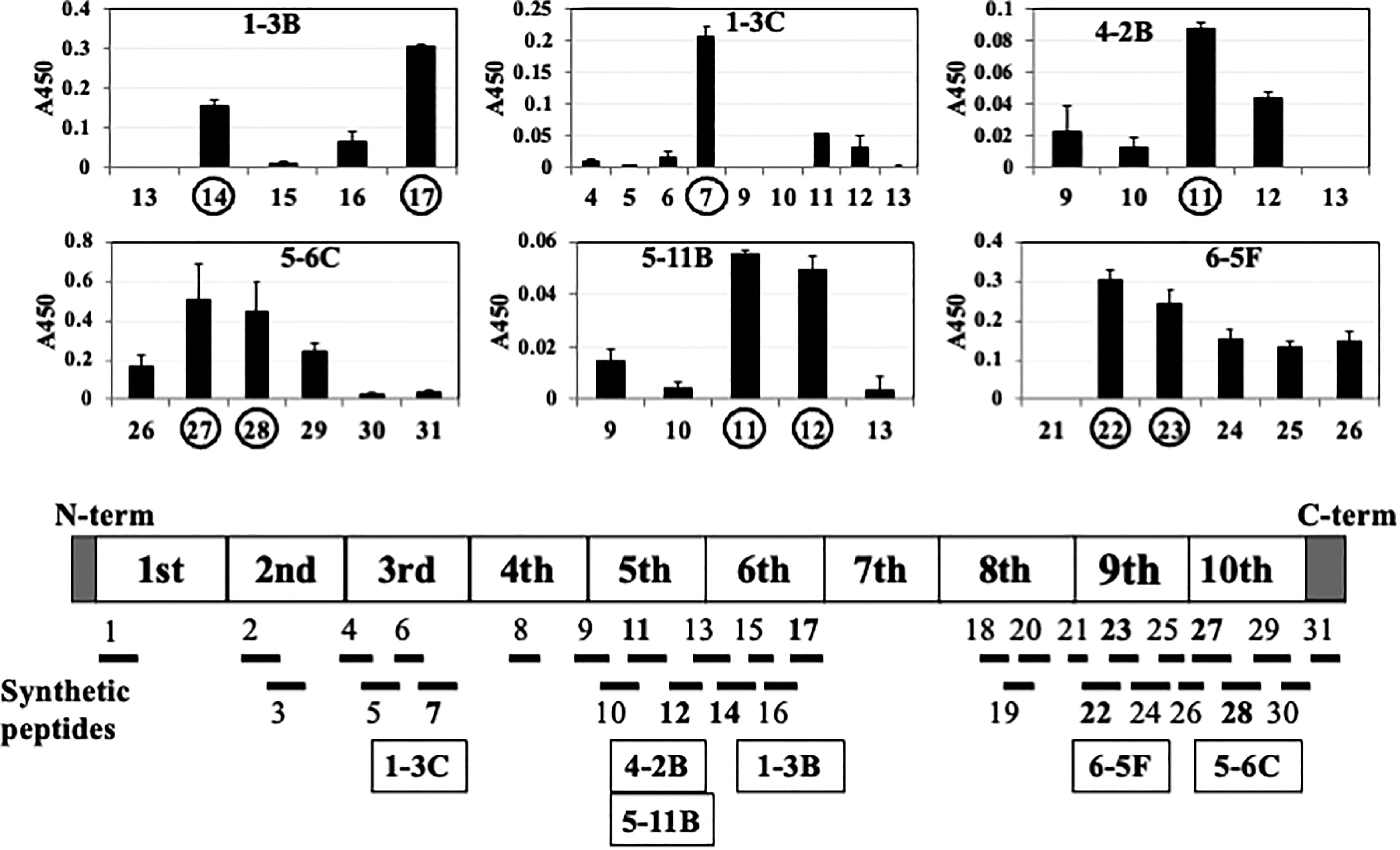
Epitope analysis using synthetic FXIII-B peptide-coated plates by ELISA. Overall, 31 synthetic peptides (bottom) were designed based on the primary amino acid sequence of FXIII-B (Supplementary Table S1). Each synthetic peptide (2.5 nmol/100 μL) was coated onto a microtiter plate and incubated with a homemade monoclonal antibody (mAb). The mAb 1-3C reacted with synthetic peptide B7, 4-2B, and 5-11B with B11 and B12; mAb 1-3B with B14 and B17; mAb 6-5F with B22 and B23; and mAb 5-6C with B27 and B28. However, the yield of mAb 3-3D was too low for the used experiment. ELISA, enzyme-linked immunosorbent assay; FXIII-B, coagulation factor XIII B subunit.
Variations in peptide binding intensities among these mAbs potentially indicate differences in their affinities (i.e., dissociation constants). However, mAb 5-6C bound to B27 and B28, mAb 5-11B bound to B11 and B12, and mAb 6-5F bound to B22 and B23 with nearly equal affinity. These results suggest that the epitopes of the mAbs are arranged as epitopic mosaics within individual Sushi domains of the FXIII-B molecule, likely representing discontinuous “structural/conformational” epitopes, as proposed by three-dimensional models of the Sushi domains of homologous human complement factor H18,19 (Supplementary Fig. S5, middle and bottom, Supplementary Table S2).
Effect of mAbs on FXIII activity and fibrin cross-linking reaction
When these mAbs were preincubated with normal plasma before thrombin activation, the AI activities of FXIII generated by thrombin–calcium activation in the presence of mAbs were generally not inhibited in the five-step dilution cross-mixing test (Supplementary Fig. S6).
We also assessed whether the mAbs could inhibit the fibrin cross-linking reaction mediated by FXIII. Out of the seven mAbs tested, only mAbs 5-6C (epitope; the 10th Sushi domain) slowed down γ-dimerization and showed the presence of the remaining γ-chain monomer (Fig. 4, top). The inhibitory effect of mAb 5–6C on γ-dimerization and α-polymerization was confirmed through a time-course study of the fibrin cross-linking reaction (Fig. 4, middle). This result is consistent with that of our previous study involving rat mAbs against FXIII-B, which also recognizes the 10th Sushi domain (all 11 of 19 rat mAbs), inhibit γ-dimerization. 20 As mouse mAb 5-6C did not inhibit FXIII AI activity, its inhibitory effect on fibrin cross-linking likely stems from interfering with the binding between FXIII-B and fibrin/fibrinogen. 20
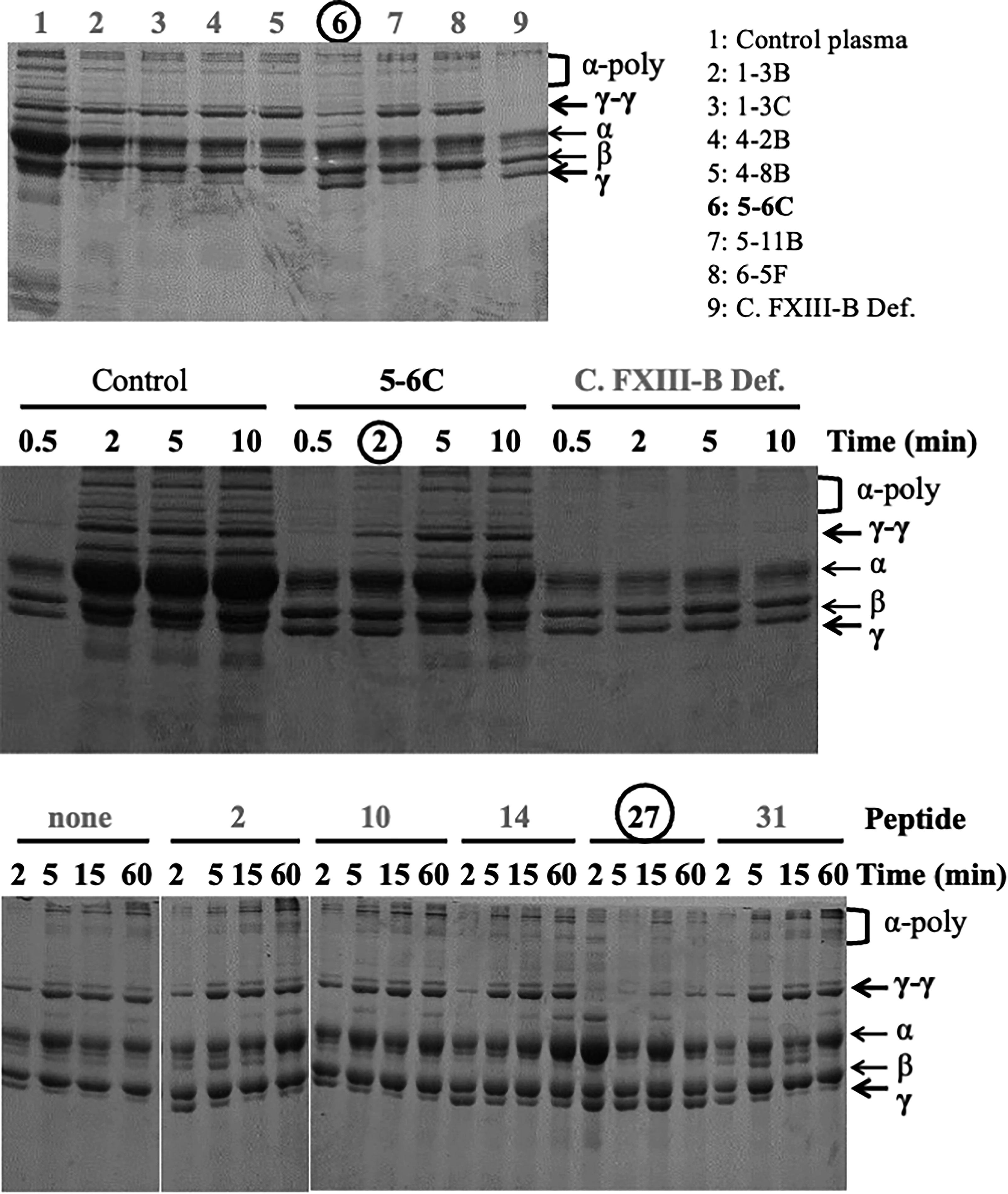
Inhibitory effect of mAbs on fibrin cross-linking activity of FXIII. Pooled normal plasma was combined with the seven mouse monoclonal antibodies (mAbs) at a 1:1 ratio and incubated at 37°C for 2 hours. (Top) FXIII in plasma was activated by adding a thrombin-calcium reagent solution, and the fibrin clot was collected 2 minutes after cross-linking reactions, such as γ-dimerization (γ-γ) and α-polymerization (α-poly). Only mAbs 5-6C (circle) demonstrated a clear delay in γ-dimerization and the persistent γ-chain monomer (γ). (Middle) Pooled normal plasma (control), normal plasma mixed with mouse mAbs 5-6C at a 1:1 ratio, and patient plasma with severe congenital FXIII-B deficiency (C. FXIII-B Def.) with alloantibodies were incubated for the specific times (0.5, 2, 5, or 10 minutes). Once again, mAbs 5-6C showed a clear delay in γ-dimerization and the presence of the γ-chain monomer at 2 minutes (circle). (Bottom) Normal plasma (10 μL) was mixed with a synthesized peptide (1 nmol/10 μL), incubated at 4°C for 1 hours, and reacted by thrombin–calcium activation for specific durations (2, 5, 15, and 60 minutes). Peptide B27 of the 10th Sushi domain (circle) showed virtually no dimerization or polymerization and the presence of the γ-chain monomer at all times. FXIII-B, coagulation factor XIII B subunit.
Application to measurement of FXIII-B antigen levels and detection of anti-FXIII-B autoantibodies
The seven mouse antihuman FXIII-B mAbs have potential applications in clinical settings, 21 including measuring plasma FXIII-B antigen levels and detecting anti-FXIII-B autoantibodies. An ICT was developed for these purposes, excluding mAbs 5-6C due to residual amounts and interference with FXIII-fibrinogen binding.
A preliminary ICT identified optimal mAB combinations (one on the strip and another in the gold-conjugate) for quantifying FXIII-B antigen (Supplementary Table S3). As a result, mAb clones 1-3B, 1-3C, 5-11B, and 6-5F were selected for further quantitation. When mAb 1-3B was conjugated to gold particles, and mAb clones 1-3C (blue circles), 5-11B (brown circles), and 6-5F (green circles) were coated on the ICT strip, the sensitivity and linearity were satisfactory (Supplementary Fig. S7, top) with potential for further extension, except for 6-5F. However, linearity was lost above ∼200% of the FXIII-B antigen at higher concentrations (Supplementary Fig. S7, bottom).
All three combinations of mAbs as the capture (strip) and the detection (gold-conjugate) antibodies resulted in exactly the same high correlation coefficient (R = 0.94) between ICT and ELISA methods (Fig. 5). The combination of mAbs 1-3C as the capture and 5-11B as the detection antibodies seemed best, because it gave nearly identical values at 50%, 100%, and 150% of FXIII-B antigen between ICT and ELISA methods. Based on these findings, we plan to develop a latex agglutination method for measuring FXIII-B antigens for laboratories and researchers without ICT strip readers.
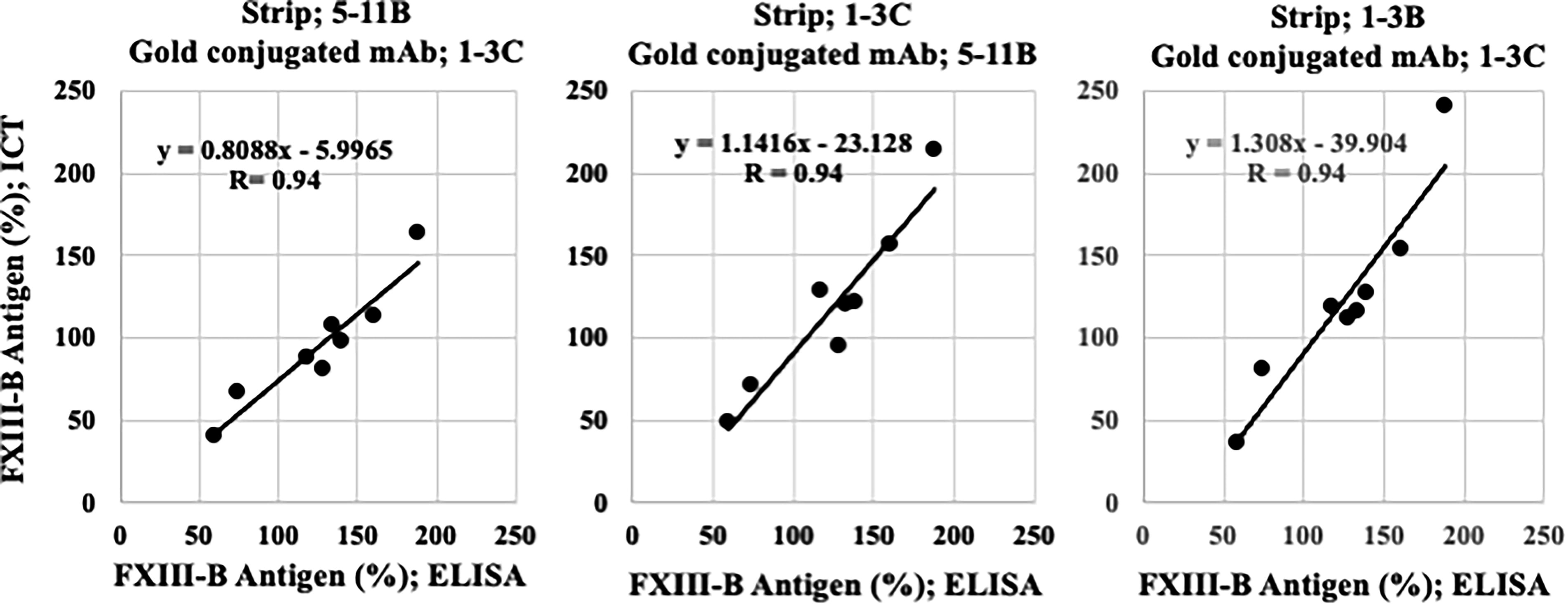
Correlation of ICT and ELISA using mouse anti-FXIII-B mAbs for quantitation of FXIII-B antigen. When either one of the three mAbs against FXIII-B was used as the capture antibody on the strip and another mAb was used as the detection antibody in the gold conjugate, there was a strong correlation between the values obtained by ICT and ELISA in the quantification of FXIII-B antigen in plasma. The correlation coefficients for the three combinations were identical (R = 0.94), but ICT occasionally resulted in slightly lower FXIII-B values than ELISA and vice versa. ELISA, enzyme-linked immunosorbent assay; FXIII-B, coagulation factor XIII B subunit; ICT, immunochromatographic test; mAbs, monoclonal antibodies.
A preliminary ICT method was also developed to detect anti-FXIII-B autoantibodies using six mouse mAbs (Supplementary Table S4). Coating mAb 1-3C on the test line (red arrow) of a strip yielded positive results in 5 out of 10 plasma samples from patients with AiF13D (red circle), while three samples remained ambiguous (orange circle, Supplementary Fig. S8), underscoring the need for enhanced ICT sensitivity.
Our goal is to integrate a new ICT system for detecting anti-FXIII-B autoantibodies using mouse mAbs, enabling simultaneous detection of both anti-FXIII-B autoantibodies and anti-FXIII-A autoantibodies with antimouse IgG as the detection antibody. However, we currently possess ICT kits for anti-FXIII-B autoantibodies 22 using “rat” mAbs and for anti-FXIII-A autoantibodies14,22 using mouse mAbs.
mAb 5-6C, binding to the 10th Sushi domain of FXIII-B, inhibits γ-chain dimerization of fibrin, critical for FXIII-B binding to fibrinogen. 20 This was demonstrated using 11 rat antihuman FXIII-B mAbs and polyclonal IgG 20 from a patient with severe congenital FXIII-B deficiency. 16 Our findings show that peptide B27 (the head of the 10th Sushi domain) inhibited both γ-dimerization and α-polymerization, whereas peptide B31(the tail of the FXIII-B) showed no such effect. Thus, mAbs 5-6C targeting the 10th Sushi domain hold promise for inhibiting thrombosis progression when humanized 23 as antibody medicines. Alternatively, developing human mAbs against FXIII-B similar to those against FXIII-A 24 or using peptide B27 as a cost-effective option for antibody-based therapies are viable strategies.
In conclusion, advancing laboratory test methods remains crucial despite perceived redundancies. Accurate diagnosis of type B AiF13D is imperative to prevent life-threatening bleeding. 25 AiF13D can develop in any elderly individuals without direct genetic abnormalities in FXIII-B or FXIII-A, highlighting the unpredictability of autoimmune acquired coagulation factor deficiencies as multifactorial diseases.26–28
Footnotes
Acknowledgments
The authors are indebted to Dr. Daisuke Sugiyama of Q-may Laboratory Corporation for performing preliminary experiments on ELISA as well as providing ICT kits, Dr. Arijit Biswas of the University of Bonn for his invaluable discussion on “structural/conformational” epitope, and all members of the Japanese Collaborative Research Group (JCRG) on autoimmune coagulation factor deficiencies (AiCFDs), as well as physicians in charge of AiCFD cases for their help in collecting patients’ clinical information and plasma samples and for participation in the nationwide survey of the JCRG.
Authors’ Contributions
T.O. conducted experimental examinations, statistical analyses, drafted figures and tables, and proofread the article. Y.M. developed and provided ICTs and proofread the article. M.S. assisted experimental examinations and proofread the article. A.I. initiated and designed the study, extracted data, and wrote, edited, and proofread the article.
Author Disclosure Statement
No authors declared competing interest.
Funding Information
This study has been supported by research aids to A.I. from
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.