
Abstract
The present work has validated the capability of using diluted Luria broth (10−3) and its amendment with antibiotics to quantify proportions of antibiotic-resistant oligotrophic bacteria contained in river water. The assay conditions for assessing fractions of antibiotic-resistant oligotrophic bacteria, especially the type and concentration of antibiotic exposure by determining LD50 concentration for each antibiotic using sensitive oligotrophic strain, have been standardized. The presence of an array of two gene cassettes including a novel dihydrofolate reductase gene, dfrA28, within the variable region of the class 1 integron in an oligotrophic test strain, MB52, in a population of oligotrophic bacteria isolated from Mahananda River water, has been described. The predicted protein consisting of 172 amino acids exhibited 76.4% amino acid identity with DfrA1. A gene for streptomycin resistance, aminoglycoside adenyl transferase aadA1, was located downstream of dfrA28. Phylogenetic analysis of MB52 identified it as a strain of Acinetobacter johnsonii.
Introduction
Conjugation among the three gene transfer modes plays a significant role in the transfer of resistance genes in Acinetobacter. 48 Acinetobacter develops resistance through acquisition of plasmids, 42 transposons, 11 or integrins. 41 There are more than nine classes of integrons reported till date, of which the most documented and well characterized are the class 1 integrons. The following are the three different segments involved: (i) 5′ conserved segment (CS) containing an intI gene that codes for integrase and an attachment site (attI) recombination site; (ii) the 3′ CS containing a combination of antiseptic resistance gene (qacE), sulfonamide resistance gene (sulI), and one or two additional ORF(s), ORF5, ORF5, and ORF6 that are hypothesized to act as transposition sites of the progenitor ISCR1 element for development of complex class 1 integrons; 47 and (iii) a variable region between 5′ CS and 3′ CS that constitutes a class 1 integron. 15 Very recently, class 1 integrons have been detected and characterized among carbapenem-resistant isolates of Acinetobacter spp. 19
Since our laboratory has been engaged in assessing the microbial risk due to antibiotic-resistant copiotrophic bacteria in river water,25–27 we have recently turned our focus to generate data on antibiotic-resistant oligotrophic river-water bacteria. Since natural environments like river systems host a majority of bacteria in nutritionally deficient (oligotrophic) conditions, we have standardized a new cultivation method for quantifying antibiotic-resistant oligotrophic bacteria from water samples. In this study, we describe the biochemical, phylogenetic, and oligotrophic characteristics of a novel A. johnsonii strain MB52, bearing class 1 integron, from River Mahananda of Siliguri, West Bengal, India. Sequence similarity and phylogenetic analyses of class 1 integron-borne gene cassettes confirmed the discovery of a novel dfr gene coding for trimethoprim resistance in A. johnsonii MB52.
Materials and Methods
Sampling
Composite water samples were collected using standard methodology 3 from a single sampling station on River Mahananda underneath the Mahananda bridge situated at the heart of the Siliguri city, West Bengal, India.
Total aerobic colony-forming units on rich and poor media
Serial dilutions of water samples of each river were made in filtered (water passed through 0.2 μm filter) and autoclaved river water and plated on Luria agar (LA) (Luria broth; HiMedia, Mumbai, India; amended with purified agar [15 g/L] [Merck, Mumbai, India]) (rich) and on 0.001 × LA (poor) to obtain total copiotrophic (eutrophic) and oligotrophic bacterial colony-forming units (CFU), respectively. Colonies on rich and poor media were evident after 24 and 96 hr of incubation, respectively.
Identification of oligotrophs
Discrete colonies on 0.001 × LA plates were numbered serially (1, 2, 3 … n). Random numbers comprising a set of n/4 unique numbers were generated using Research Randomizer software (www.randomizer.org). Colonies corresponding to the numbers from the set of unique numbers were picked up with sterilized toothpicks and imprinted on the master plate made of R2A agar (a standard environmental cultivation medium 35 ). Each master plate was replicated separately on 1 × LA, 0.001 × LA, and R2A agar. After incubation of these plates for 72 hr at 30°C, colonies that had grown on 0.001 × LA and R2A agar but not on 1 × LA were termed as “obligate oligotrophs,” whereas colonies that were able to grow on all three different plates were termed as “facultative oligotrophs.”
Selection of antibiotic-sensitive oligotrophic isolates and determination of LD50 of each of 12 antibiotics used in this study
The master plates constructed with oligotrophic strains were replicated on antibiotic plates. For each antibiotic, three plates were used. The concentration of antibiotics in the plates was 10, 25, and 50 mg/L, respectively. The plates were incubated for 96 hr at 30°C. Colonies that failed to appear in all 12 different antibiotic plates of 25 mg/L concentration were tentatively chosen as sensitive isolates (n = 5). For determination of LD50 (LD50 of each antibiotic is the dose required to kill half the members of the tested bacterial population), a duplicate set of 16 culture tubes each containing sterile 5 ml R2A broth was amended with 0, 0.25, 0.5, 0.75, 1.0, 1.25, 1.5, 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, and 10.0 mg/L of antibiotic, respectively. Each tube was inoculated with pregrown sensitive bacterial culture to an initial cell density of 106 CFU/ml. After 72 hr of incubation at 30°C, optical density was recorded at 540 nm. The particular tube (with defined antibiotic concentration) wherein OD540 was recorded to be half the OD value of the tube without the antibiotic was taken as LD50.
Antibiotic resistance determination
Obligate oligotrophic isolates were tested for susceptibility to a panel of 12 antibiotics, representing 7 different classes. The concentrations of the antibiotics were chosen as five times the calculated LD50 values. Antibiotics and their concentrations employed in this investigation were as follows: aminoglycosides (azithromycin, 5 mg/L; kanamycin, 5 mg/L; netilmicin, 3.75 mg/L; and streptomycin, 2.5 mg/L); antifolates (trimethoprim, 5 mg/L; sulfamethoxazole, 15 mg/L); cephalosporins (cefepime, 7.5 mg/L; cefotaxime, 7.5 mg/L); penicillin (ampicillin, 25 mg/L); quinolones (ciprofloxacin, 5.0 mg/L; livofloxacin, 5.0 mg/L); others (chloramphenicol, 30 mg/L; oxytetracycline, 15 mg/L). R2A agar was used as the basal medium. The desired concentrations of the antibiotics were stirred into the melted agar at approximately 45°C and immediately poured into Petri plates to minimize the exposure of elevated temperatures. Replica plating method was employed for determining the antibiotic resistance profile of individual isolates. The premade master plates were replicated onto R2A agar plates, each containing a single antibiotic at the concentration previously mentioned. The final plate replicated was the plain R2A agar plate. The control plate was replicated last to confirm the successful inoculation of the preceding plates. The replicated plates were incubated at 30°C for 48 hr and drug resistance was determined. The isolates were considered resistant to multiple antibiotics, only if their growth in the presence of antibiotics was as well developed as their growth on the control plates. Any sign of inhibition or sensitivity was considered to be indicative of sensitivity. The isolates were considered multiple antibiotic resistant (MAR) if growth on at least two different antibiotic-containing plates was at least equal to that on the growth control without antibiotics.
On the other hand, sensitivity and resistance of facultative oligotrophic isolates were also determined following agar dilution method using Mueller–Hinton (MH) agar, as described in EUCAST definitive document, E. Def 3.1 (2000) [www.escmid.org/fileadmin/src/media/PDFs]. E. coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, and Staphylococcus aureus ATCC 29213 were taken as the quality control strains. Minimum inhibitory concentrations (MICs) of antibiotics (recommended for Acinetobacter in EUCAST [www.srga.org/eucastwt/MICTAB/index.html]) in MH agar were determined for the test strain MB52.
Test strain
From the collection of class 1 integron-bearing MAR oligotrophic bacteria, we selected one representative strain, MB52, for this study. MB52 could resist trimethoprim (30 mg/L) and streptomycin (10 mg/L). It was maintained by periodic transfer to R2A agar slants and stored at −20°C after supplementing with glycerol (20% v/v) for further study.
Characterization of MB52
Diagnostic tests for MB52
Gram-staining, motility, and growth at different pH and aerobic/anaerobic conditions were analyzed according to the standard procedures. Growths at different temperatures were observed in 1× Luria broth, 0.001× Luria broth, 1× R2A broth, 1× nutrient broth, and tryptone soya broth (TSB) at 4, 10, 30, 37, and 42°C. All phenotypic tests were performed in TSB/agar (HiMedia). Tests were carried out at 30°C unless otherwise specified. Gelatin hydrolysis and hemolysis were performed as previously described. 8 A 1% (v/v) filter sterilized ethanol was amended to 3 ml XfD media, 1 and tubes were incubated for 24 to 48 hr to detect ethanol as the sole carbon source for energy. Catalase activity was tested using 3% H2O2. Urea hydrolysis, citrate utilization (Simmons citrate), indole production, oxidase activity, and carbohydrate utilization/fermentation test were performed according to the manufacturer's instructions (HiMedia).
DNA isolation, amplification, cloning, and sequencing of 16S rDNA coding rRNA
Whole cell DNA was extracted according to the method described by Ling et al. 24 27F and 1492R primers, as described by Lane, 20 were used to amplify the 16S rRNA gene. The amplified PCR product was purified using the Gen Elute™ PCR Clean-up kit (Sigma-Aldrich, St. Louis, MO), subsequently cloned into pGEM-T easy vector system II (Promega, Madison, WI), and then transformed into E. coli JM109. The recombinant plasmid was sequenced with T7 and SP6 primers from both ends, and the middle portion was sequenced with 530F primer. 20 Sequencing was performed at DBT-supported DNA sequencing facility at University of Delhi, South Campus, India.
Phylogenetic affiliation of MB52
The 16S rRNA gene sequence of A. johnsonii MB52 was aligned with the 16S rRNA gene sequences of all the known species of Acinetobacter retrieved from GenBank using clustal X version 1.83. 46 The 16S rRNA gene sequence of E. coli was included as outgroup. The resulted multiply aligned sequence was corrected and edited, and approximately 1334 bp long nucleotide sequence of all the Acinetobacter was selected for further analysis. The rest of the nucleotide sequence from both the ends was omitted due to alignment ambiguities. Phylogenetic trees were inferred by using the neighbor-joining (N-J), 39 maximum likelihood, and Unweighted Pair Group Method with Arithmetic Mean (UPGMA) methods to ensure consistency of the clusters formed. The topology of the phylogenetic trees was evaluated using a bootstrap analysis 12 of N-J method based on 1,000 replications.
Viability and growth of MB52 in diluted (10−3) Luria broth
Inoculum was prepared by transferring a single colony of 24-hr-old culture MB52 into 10 ml sterile 1× Luria broth (pH 7.0) in a 100 ml Erlenmeyer flask. The inoculated medium was incubated at 30°C for 20 hr without agitation. The culture was harvested by centrifuging at 7,000 rpm for 7 min at 4°C and washed twice with sterile saline (0.5% NaCl) water to ensure removal of traces of media. The washed pellet was finally suspended in 3 ml 0.001× Luria broth. Aliquots of 0.5 ml of concentrated (1.75 × 109 to 2.25 × 109 cells/ml) cell suspension(s) were added to 25 ml volume(s) of diluted (0.001×) Luria broth in 250 ml Erlenmeyer flask(s). The flasks were kept at 30°C (without shaking) throughout the period of investigation. Survivability of MB52 cells in 0.001× Luria broth was assessed through dilution-plating of pure culture aliquots at different time intervals on fresh 1× LA.
DNA preparation, detection of class 1 integron, cloning, sequencing, and expression of variable region
Bacterial DNA was extracted from a single colony as described in DNA isolation, amplification, cloning, and sequencing of 16S rDNA coding rRNA section. Gene cassettes from class 1 integron were amplified using CS-PCR method. 23 PCR was performed using the primers 5′ CS (5′-GGCATCCAAGCAGCAAG-3′) and 3′ CS (5′-AAGCAGACTTGACCTGA-3′), as previously described. 23 The amplicon was cloned into pGEM-T easy vector system II (Promega) and then transformed into E. coli JM101. Transformants were selected on LA plates containing ampicillin (50 mg/L). The recombinant plasmid was sequenced with T7 and SP6 primers from both ends and the middle portion was sequenced by primer walking. For studying the expression of gene cassettes, recombinants obtained from ligating pJET1.2/blunt vector (Fermentas Life Sciences, Vilnius, Lithuania) with insert (CS-PCR product) in proper orientation were selected on trimethoprim (5 mg/L) and streptomycin (2.5 mg/L) amended LA plate. One of the clones (pAK01) was used for determining the maximum tolerance of trimethoprim and streptomycin. Plasmidless E. coli JM101 was used as control.
Sequence analysis and phylogenetic consideration of the novel dfrA gene
Nucleotide sequence analysis was performed using BLAST 2 search programs (National Center for Biotechnology Information). A phylogenetic analysis of DfrA at the amino acid level (DfrA protein translated from dfrA gene cassette) was performed by making a phylogenetic tree based on one representative microorganism for each trimethoprim-resistance-mediating DfrA protein. The multiple alignments were made using the program ClustalX (version 1.83) 46 ; an N-J tree was made based on this alignment, which was used as a guide tree for the final alignment used to produce the final bootstrap N-J tree.
Nucleotide accession number
The nucleotide sequence data reported here have been assigned the accession no(s). FN263373 and FN263374 by EMBL.
Results
Oligotrophic bacterial load in Mahananda river water and strain selection
Cultivable oligotrophic bacterial load from Mahananda river water samples varied from 3.5 × 103 to 5.7 × 104 CFU/ml (from February 2008 to June 2008). Fifty-five percentage of the total oligotrophs were facultative oligotrophs. About 37.23% of the facultatively oligotrophic bacteria were sensitive to all the antibiotics used in this study. Obligate oligotrophs were found refractory to antibiotic sensitivity testing by EUCAST method, as they did not grow in MH agar. Only facultative oligotrophs could be tested for sensitivity and resistance using MH agar. Antibiotic resistance parameters, derived from facultative oligotrophs, determined in R2A agar (as described in Antibiotic resistance determination section) were well comparable with EUCAST clinical MIC breakpoints. MAR oligotrophic bacteria were at least 1.5 times more abundant than singly resistant ones. From the collection of antibiotic-resistant isolates, one representative facultative oligotrophic strain, MB52, sensitive to growth at 37°C and resistant to trimethoprim and streptomycin, was used for this study.
Phenotypic and 16S rRNA gene characterization of MB52
MB52 cells were strictly aerobic, nonfermentative, facultatively oligotrophic, gram-negative, nonmotile, coccobacilli. The colonies were circular, convex, smooth, and slightly opaque on TSB; colony diameter ranged from 0.5 to 1.0 mm after 23 hr, 1.5 to 2.0 mm after 40 hr, and 2.0 to 3.0 mm after 60 hr of incubation. The cells could grow in a temperature range of 10°C to 30°C within 24 hr in TSB medium (optimum temperature 28°C to 30°C) but failed to grow at 37°C. The cells were positive to catalase and growth on citrate (Simmons) plate. MB52 cells were negative to tests of oxidase, gelatinase, urease, and hemolysis. These cells could not reduce nitrates to nitrites. The cells did not produce indole, γ-glutamyltransferase, and β-xylosidase. They could utilize L-arginine, L-aspartate, D-L-lactate, ethanol, and malonate. MB52 cells could not utilize trans-aconitate, β-alanine, DL-4-aminobutyrate, azelate, 2,3-butanediol, glutarate, histamine, L-histidine, malonate, L-leucine, D-malate, phenylacetate, and L-tyrosine. The phenotypic features of MB52 showed a high degree of similarity (∼100%) with the published characteristics of A. johnsonii ATCC 17909T. 29 MB52 was found sensitive to all antibiotics recommended by EUCAST for Acinetobacter except cotrimoxazole (MIC, 20 mg/L in correspondence to EUCAST MIC breakpoint [S ≤/R> is 2/4]) and streptomycin (MIC, 10 mg/L; streptomycin was absent in the list of EUCAST). BlastN analysis of the 16S rRNA gene sequence of the isolate MB52 (GenBank accession no. FN263374) produced maximum identity (99%) with the 16S rRNA gene sequence of A. johnsonii ATCC 17909T (GenBank accession no. Z93440). The isolate MB52 branched deeply with the Acinetobacter cluster consisting of A. johnsonii ATCC 17909T (Z93440), Acinetobacter bouvetti 4B02T (AF509827), and A. schindleri LUH5832T (AJ278311) in the phylogenetic tree (N-J) constructed with 16S rRNA sequences of Acinetobacter spp (Fig. 1). The same output was obtained with maximum likelihood and UPGMA clustering (data not shown).
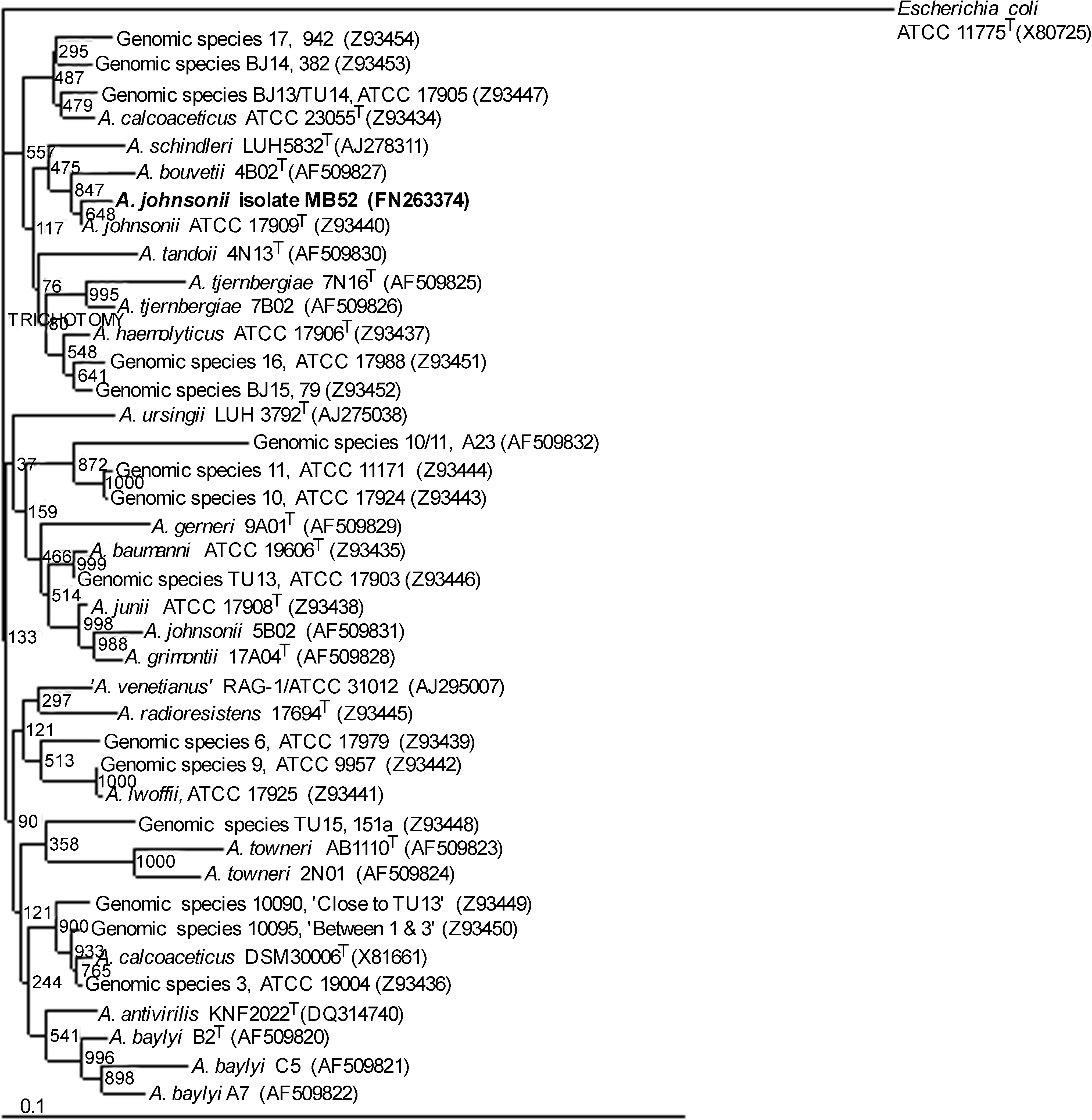
Rooted 16S rRNA sequence–based tree showing the position of isolate MB52 (bold letters) with the other species from genus Acinetobacter and related taxa. Escherichia coli is used as outgroup. The tree was constructed using neighbor-joining method. The numbers at the branching points are the proportion of 1,000 bootstrap resamplings, supporting the tree topology. EMBL/GenBank accession numbers are given in parentheses. Scale bar shows a 10% sequence divergence.
Growth of A. johnsonii MB52 in diluted (10−3 times) Luria broth
MB52 cells were able to grow in 0.001× Luria broths without supplementation of any other carbon or nitrogen source or growth factor (Fig. 2). An increase of 2.5 times the initial cell number was noted in a span of 8 days (under the experimental condition elaborated in Viability and growth of MB52 in diluted (10−3) Luria broth section). The ability of A. johnsonii MB52 to survive (without reduction in viable cell number since inoculation) and grow in a low nutrient medium explains the oligotrophic nature of the strain.

Viability and growth of MB52 in 0.001 ×-Luria broth. Error bars show the standard errors.
Detection of class 1 integron and characterization of novel dfrA28 gene
An amplicon of 1694 bp was obtained from MB52 by CS-PCR. PCR with a primer combination of int2F and 3′ CS resulted in an amplicon larger by an approximate size of 600 bp than the amplicon derived from 5′ CS and 3′ CS primers (data not shown). Purified CS-PCR product of MB52 was cloned in a pGEM-T Easy Vector and was subjected to DNA sequencing, and gene cassettes were identified by sequence analysis. The nucleotide sequence showed the presence of two genes, the novel dihydrofolate reductase gene (dfrA28) and aminoglycoside adenyl transferase aadA1 gene. Both the genes were present as gene cassettes inserted into the variable region between 5′ CS and 3′ CS of a class 1 integron, as they were flanked by integrase recombination core sites and had conserved features at the 3′ ends of the genes with a 59 base element. The trimethoprim resistance gene, dfrA28, was flanked by a core element (GTTAACC) that located 29 nucleotides 5′ to the start of the reading frame and differed from the consensus sequence of the core element (GTTRRRY) by a single nucleotide. Characterization of two gene cassettes revealed a significant association between the nature of the gene cassettes and the corresponding antibiotic resistance phenotype of the isolate, MB52. Thirty-five nucleotides downstream of the attI1 site, a 519-bp ORF, potentially encoding a polypeptide of 172 amino acids, were identified. The start of the ORF began with an ATG start at positions 134 to 136 and terminated with a TAA stop codon at positions 650 to 652. The start codon was preceded by a plausible SD sequence (TGAGGAAGA). The translated polypeptide sequence was compared with other amino acid sequences of the SwissProt data base. The sequences with the best scores were all Dfr sequences. The amino acid sequence of the novel Dfr shared the highest degree of amino acid identity with DfrA1 (76.4%) (GenBank accession. no. AJ419168). The degrees of identity between the DfrA28 (novel Dfr sequence of MB52) and the sequences of other Dfr ranged between 12.9% and 70.8%, thus placing DfrA1 and DfrA28 in an indisputable monophyletic group in the phylogenetic analysis (Fig. 3). The nucleotide sequence immediately 3′ of the end of dfrA28 encoded a structure of 133 nucleotides, which was recognizable as 59-base element (nt 653 to 785). The PROSITE motif search has revealed the presence of dihydrofolate reductase signature sequence, VIGngpdIPWsakg.EqllFkaiT, in the sequence. The phylogenetic tree confirmed DfrA28 to be most closely related to DfrA1. DfrA28 clustered together with DfrA1 and DfrA15 (Fig. 3). The second gene cassette contained an ORF of 792 nucleotides starting at an ATG codon and ending with a TAA stop codon (nt positions 789 to 1580; Fig. 4). This coding sequence, designated aadA1, shared 100% identity with the same gene (EMBL accession no. DQ388124).
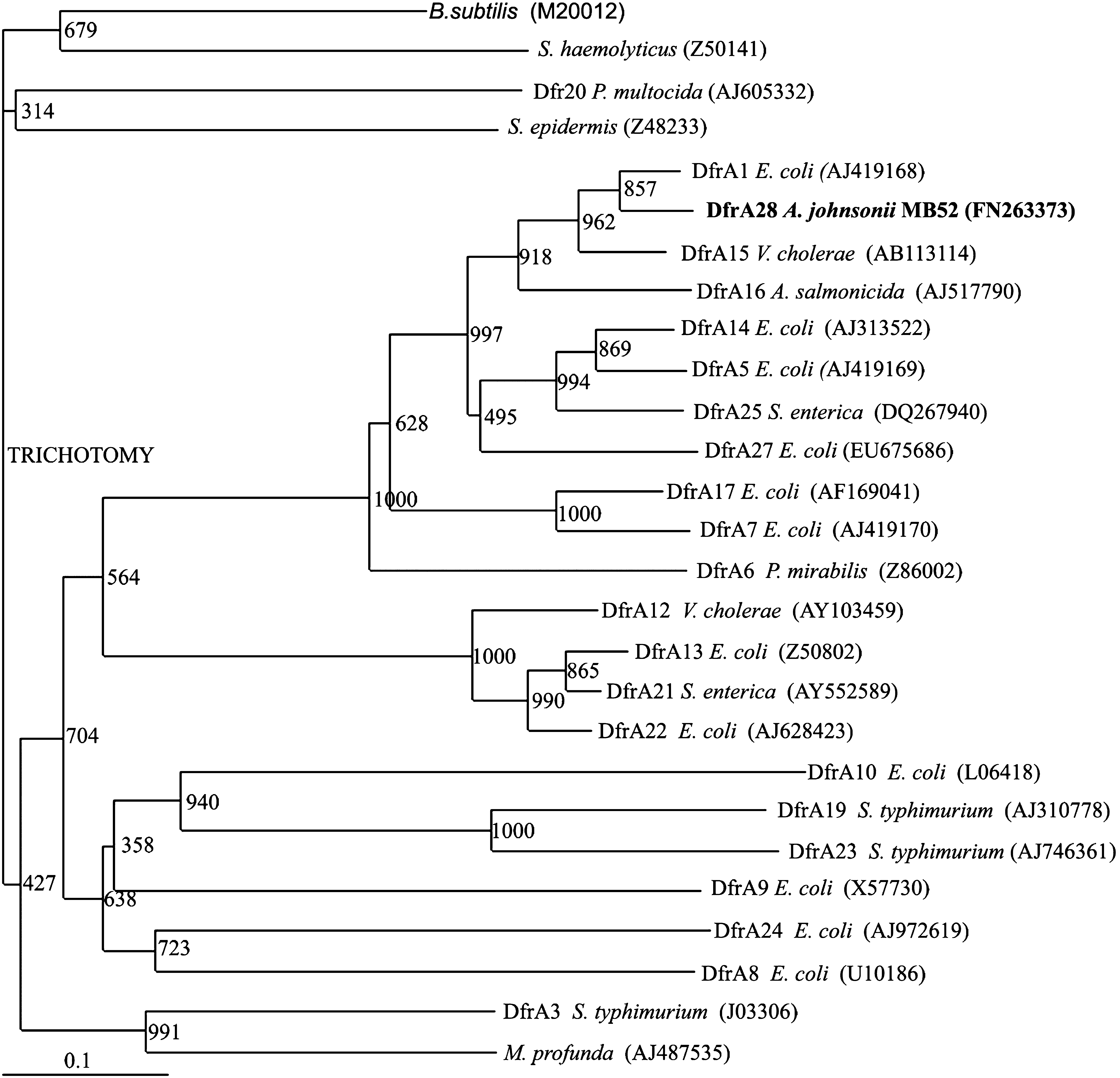
Phylogenetic tree of the DfrA proteins involved in trimethoprim resistance. The number at each major branch point refers to the percentage of times that a particular node was found in 1,000 bootstrap replications. Source and the GenBank accession (in parentheses) numbers are given for each DfrA protein. The placement of DfrA28 in the tree is indicated in bold letters. DfrB proteins, which differ distinctly from DfrA proteins in size and structure, have been excluded from this phylogenetic analysis.

Schematic representation of the CS-PCR product of A. johnsonii MB52. CS, conserved segment; attI1, attachment site; dfr, dihydrofolate reductase; aad, aminoglycoside adenyl transferase; be, base element. Numbers above the integron structure correspond to sequence positions in EMBL accession no. FN263373.
Expression of dfrA28 (coding novel dihydrofolate reductase for trimethoprim resistance) and aadA1 (coding amino glycoside adenyl transferase for streptomycin resistance) was demonstrated in E. coli JM101 bearing the recombinant plasmid, pAK01. The plasmid-bearing strain tolerated trimethoprim and streptomycin up to 1,000 and 10 mg/L, respectively. The MIC of the control strain (plasmidless) was restricted to 2 and 1 mg/L for trimethoprim and streptomycin, respectively.
Discussion
There are only few reports on enumeration and culture of oligotrophic bacteria from water sources. In an earlier study, it was found that R2A medium as a spread plate provided the highest colony counts, and the mean oligotrophic bacterial density (from potable water samples) was found to be 4.3 × 103 CFU/ml. 35 Very recently, almost simultaneously with our work, diluted Luria-Bertani broth with antibiotics was used to detect oligotrophic and antibiotic-resistant bacteria in drinking water and swine manure samples. 30 Our results have also validated the capability of using diluted Luria broth (10−3) and its amendment with antibiotics to quantify proportions of antibiotic-resistant oligotrophic bacteria contained in river water. In fact, standardization of the assay conditions for assessing fractions of antibiotic-resistant oligotrophic bacteria, especially the type and concentration of antibiotic exposure by determining LD50 concentration for each antibiotic using sensitive oligotrophic strains, marked a major advancement in this area of water research. In the present study, the Mahananda river water samples displayed oligotrophic bacteria at a range of 3.5 × 103 to 5.7 × 104 CFU/ml. River Mahananda is the principal river flowing through Siliguri, the fastest growing city situated at the foothills of the Himalayas of West Bengal, India, with a population of over 500,000. A huge quantity of community waste water and other kinds of wastes, including animal wastes from disorganized cattle farms situated by the side of the river and hospital wastes, are ultimately discharged into the main River Mahananda. 25 An earlier study from our laboratory revealed the presence of a fairly high percentage of antibiotic-resistant copiotrophic bacteria in Mahananda river water. 25 The present study has shown that MAR oligotrophic bacteria were at least 1.5 times more abundant than singly resistant ones (details to be published elsewhere). The standard medium used for purification of oligotrophic bacteria was R2A agar, which was found to contain dissolved protein to the extent of 830 mg/L. The average dissolved protein in River Mahananda was estimated to be 130 mg/L (data not shown). The mean protein content of the medium (0.001× Luria broth) used in this study as selective plates for isolation of oligotrophs was found to be 7.8 mg/L (approximate 17 times lower than the content of River Mahananda). A previous study reported the presence of the intI1 gene among 3.6% of the nonselected gram-negative bacteria from an estuarine environment. 37 The percent occurrence of integron-positive isolates among environmental samples including fish farms, 40 irrigation water sources, 36 and other aquatic environments was much lower7,31,32 compared with that recovered from clinical samples. 4 In all these studies, the incidence of class 1 integrons was scored among copiotrophic isolates that grow on rich nutrient medium. But the test strain, MB52, of this study is one representative of the antibiotic-resistant population, comprised of only oligotrophic bacteria recovered on 0.001× LA from River Mahananda. Oligotrophic bacteria have also been isolated from clinical materials. 45 Quite a number of oligotrophic bacteria exhibit antibiotic resistance and can, therefore, be a potential reservoir of antibiotic resistance genes that can be acquired by pathogens through diverse gene transfer mechanisms. Due to their potential clinical importance, oligotrophic bacteria merit attention. The facultatively oligotrophic strain used in this study was characterized as A. johnsonii MB52 (ascertained from phenotype as well as from 16S rRNA phylogeny) (Fig. 1). Actually, all bacteria under the genus Acinetobacter are strictly aerobic, oxidase-negative, and catalase-positive. 8 They constitute a unique group of gram-negative bacteria that may be natural residents of human skin, 21 with carriage rates of 42.5% and ∼75% in healthy individuals and patients who are hospitalized, respectively. 43 Viability assay and growth assessment of A. johnsonii MB52 cell concentrate in 0.001× Luria broth for more than a week by taking viable cell count of the cell suspension in 1× LA at different times (Fig. 2) demonstrated its adaptive ease to switch from oligotrophy (ability to survive and grow in extremely poor nutrient conditions) to copiotrophy (ability to form colonies in a rich medium). Such facultative nature of oligotrophy, as shown by the A. johnsonii strain MB52, may contribute to the reported adaptation of remaining viable on both moist and dry surfaces such as polyvinyl chloride, rubber, ceramics, and various types of medical equipment51,52 and cause nosocomial infection in immunocompromised patients. 6 Many studies have demonstrated the presence of antibiotic resistance genes in Acinetobacter spp. on plasmids, transposons, and integrons and, more recently, on a resistance island.11,13,38,42 Most recently in Malaysia, class 1 integrons were detected in 31 of 39 carbapenem-resistant strains of Acinetobacter spp., of which two A. calcoaceticus strains harbored an integron-borne blaIMP-4 metallo-β-lactamase, one of which was located on a 36-kb plasmid. 19 The test strain, MB52, harbored a class 1 integron that carried two gene cassettes, dfrA28 and aadA1, respectively. Several hundreds (>300) of different complete cassette arrays flanked by the 5′-CS and 3′-CS have been identified in GenBank and majority arrays had two or three gene cassettes (within the limitation of CS-PCR conditions employed). 33 Most cassette array sequences in class 1 integrons with the 5′-CS and 3′-CS from gram-negative bacteria deposited in GenBank were from E. coli (21%), P. aeruginosa (19%), Salmonella spp. (14%), A. baumannii, Klebsiella pneumoniae, or Vibrio cholerae (each ∼6%). 33 Cassette arrays flanked by 5′-CS and 3′-CS had two gene cassettes in the test strain MB52 of this study (Fig. 4). The first gene cassette, dfrA28 gene, of the array is—to the best of our knowledge—the first novel trimethoprim resistance gene detected in A. johnsonii. Trimethoprim on its own or in combination with sulfamethoxazole is an antibacterial drug for the control of pathogenic gram-negative organisms. Trimethoprim prevents the reduction of dihydrofolate to tetrahydrofolate selectively by inhibiting the bacterial dihydrofolate reductase (Dfr).18,44 Production of an additional plasmid, transposon, or cassette-borne Dfr is the most widespread mechanism of resistance to trimethoprim in bacteria, which in contrast to the chromosomal enzyme is less susceptible to inhibition by trimethoprim. 18 More than 30 different trimethoprim-resistance-mediating dihydrofolate reductase (dfr) genes, subdivided into two major types, 1 and 2 (referred to as dfrA and dfrB), have been observed till date. 53 A novel trimethoprim resistance gene is claimed when the translated dihydrofolate reductase protein (Dfr) encoded by the gene has <95% identity at the amino acid level compared with known Dfr proteins. 44 The phylogenetic tree confirmed DfrA28 to be most closely related to DfrA1. DfrA28 clustered together with DfrA1 and DfrA15 (Fig. 3). Taking into consideration the trimethoprim resistance phenotype and the phylogenetic relationship with other Dfr type A proteins, we predict this gene to encode a novel trimethoprim-resistance-mediating dihydrofolate reductase named dfrA28 according to the guidelines 14 for naming new trimethoprim-resistance genes.
Footnotes
Acknowledgments
The research was supported by a major research grant from Department of Biotechnology, Govt. of India (BT/PR-7164/BCE/08/448/2006). One of the authors, A.K., was supported by the DBT-BET, Ph.D. studentship from the Department of Biotechnology, Govt. of India (DBT-JRF/05-06/71/29.08.05); S.M. was supported by CSIR Research Associateship grant (ACK. No. 313165/2K7/1).
Disclosure Statement
No competing financial interests exist.