
Abstract
The Pseudomonas aeruginosa isolate TS-832035 was responsible for an outbreak that occurred in an Italian hospital between 1999 and 2002. It exhibited a high-level resistance to carbapenems due to the contemporary presence of two independent mechanisms: the production of a carbapenemase, coded by a blaVIM-1 determinant carried by the chromosomal class 1 integron In70.2 (containing also the aacA4, aphA15, and aadA1 genes in its cassette array), and the lack of the OprD porin. We compared TS-832035 with a strictly related isolate, TS-103, whose resistance to carbapenems was due to the lack of the OprD porin only, as it did not carry In70.2. We evaluated their growth kinetics, in both separate cultures and competition assays, under permissive conditions. These experiments highlighted a significant in vitro fitness cost associated with the integron. On the contrary, none of the resistance determinants other than the blaVIM-1 seemed to confer a real selective advantage to its host. Comparison of these results with the in vivo behavior, showing that the In70.2-carrying isolates largely prevailed over the In70.2-lacking ones, besides the detection of similar integrons in other Italian clinical isolates, evidenced the need to investigate accurately the causes of their large distribution, as possible soft spots could exist in the ability of their hosts to adapt to the hospital settings.
Introduction
An epidemiologic survey performed at the Trieste University Hospital (northeastern Italy) between 1999 and 2002 revealed a remarkable spread of an MDR Pseudomonas aeruginosa strain, named TS-832035, which carried the chromosomal integron In70.2 containing four gene cassettes (blaVIM-1, aacA4, aphA15, and aadA1) in its variable region conferring resistance to β-lactams, including carbapenems, and to several aminoglycosides.14,25 Moreover, some other P. aeruginosa isolates, strictly related to TS-832035 but lacking the integron In70.2, were detected, but they remained a minor component within the cluster during the 3 years of surveillance. They showed an MDR phenotype like that of TS-832035, differing only in the susceptibility level to carbapenems. Their carbapenem minimal inhibitory concentrations (MICs) were lower overall compared with those of the blaVIM-1-positive isolates but not so low to consider them clinically susceptible (imipenem MICs, 16–32 μg/ml; meropenem MICs, 8–32 μg/ml), because of the loss of the OprD protein showed by the entire clonal lineage.13,14 Undoubtedly, the selective pressure exerted by the carbapenem use inside the hospitals confers to the metallo-β-lactamases (MBLs)-producer isolates a great advantage over the carbapenem-susceptible ones. But can we really say that MBL producers get ahead also of strains that express different carbapenem resistance mechanisms and have MICs equal or a little higher than the breakpoint values? We compared the TS-832035 isolate with a clonally related one, TS-103, lacking in the integron In70.2, to evaluate the fitness cost associated with the maintenance of a chromosomal integron with respect to the advantages it may confer to the host.
Preliminary investigations at a molecular level (e.g., multilocus sequence typing [MLST]) were done to confirm the clonality of TS-103 with TS-832035.
Materials and Methods
Bacterial isolates
Two clinical isolates of P. aeruginosa, TS-832035 and TS-103, belonging to a clonal lineage that spread in the Trieste University Hospital at the beginning of the 2000s, 13 were investigated. They belonged to the previously described cluster A, 13 most members of which carried a blaVIM-1 cassette in the variable region of the class 1 chromosomal integron In 70.2 25 ; TS-832035 was the first VIM-1–positive isolate detected during the 3 years of surveillance, whereas TS-103 was one of the few VIM-1 negatives.
Genotyping of isolates
The close genomic relatedness between TS-832035 and TS-103, previously verified by random amplification of polymorphic DNA typing and pulsed-field gel electrophoresis analysis of SpeI-digested genomic DNA, 13 was confirmed by MLST, performed according to the protocol described by Curran et al. 7 and modified by Giske et al. 10 Nucleotide sequences of the seven housekeeping genes were determined for both strands and compared with the sequences in the MLST database (http://pubmlst.org/paeruginosa/) for the assignment of allelic numbers and sequence types.
Polymerase chain reaction detection of class 1 integrons
The presence of integrons in the two isolates was investigated by using polymerase chain reaction (PCR) with the 5′-CS and 3′-CS 18 primers. The presence of the gene cassettes within the variable region of the integron In70.2 was detected by PCR using the following forward primers: the VIM-DIA/f, 25 ant(3")-I-3′, 18 aphA-f (5′-TATTTCCTGATCGCAATAC) designed on an internal region of the cassette aphA15, and 3′-CS as reverse primer. PCRs were carried out in a 25 μl volume mixture containing 1 U of AmpliTaq Gold enzyme (Applied Biosystems, Foster City, CA), 20 ng template DNA, and 0.5 μM of each primer in the reaction buffer recommended by the enzyme manufacturer. Amplification conditions were as previously described18,25 except for annealing temperature of 53°C for the couple of primers aphA-f and 3′-CS. The PCR products obtained with primers 5′-CS and 3′-CS were subjected to sequence analysis on both strands (BMR Genomics, Padua, Italy).
The maintenance of the integron In70.2 by TS-832035 was verified by PCR with the VIM-DIA/f and 3′-CS primers at the beginning and at the end of each growth kinetic and competition assay.
Amplification and sequencing of blaampC genes
The ampC-type genes were amplified and sequenced as previously described. 26
In vitro susceptibility testing
MICs were determined by using the agar dilution method according to the Clinical and Laboratory Standards and Institute guidelines 5 for the following antimicrobials: the beta-lactams meropenem, imipenem and ceftazidime ± cloxacillin, the aminoglycosides sisomycin, amikacyn, gentamycin, tobramycin, netilmycin, kanamycin, streptomycin, and spectinomycin, and the disinfectants mercuric chloride, chlorhexidine, and cetrimide. P. aeruginosa ATCC 27853 was used as a quality control strain.
Determination of biological fitness
The cost of the integron In70.2 on the fitness of TS-832035 was determined by performing growth kinetics and direct competition assays against the clonal isolate TS-103 in three media differing in nutrient availability: a rich medium (Luria Bertani [LB] broth) and a minimal medium (28 g/L K2HPO4, 12 g/L KH2PO4, 0.4 g/L MgSO4 · 7H2O, 4 g/L (NH4)2SO4) added with a rich carbon source (0.4% [w/v] glucose) or with a poorer carbon source (0.4% [w/v] sodium acetate).
Growth kinetic data were obtained by measuring optical density at 600 nm (OD600). Each of the two isolates was inoculated into 5 ml of LB broth and cultured to the stationary phase. Cultures were then diluted to an OD600 of 0.1 in 10 ml of sterile phosphate-buffered saline; 100 μl of each dilution was separately inoculated in 100 ml of fresh medium and incubated with shaking (200 rpm) at 37°C. The equal number of colony forming unit (CFU)/ml of the two isolates at the beginning of the incubation period was confirmed by plating serial dilutions on LB agar plates. OD600 was monitored for 50 hr. Results are given as mean ± SD of at least six experiments performed with independent cultures.
For competition assays, isolates were grown and diluted as described earlier, mixed into the fresh medium, and counted on LB agar at different times over the course of 50 hr. The contribution of each isolate to the total population was estimated by colony hybridization. Briefly, plates containing suitable dilutions of each culture (50–300 colonies per plate) were replica plated onto a Hybond-N+ membrane (GE Healthcare, Buckinghamshire, United Kingdom). Then the cells were lysed and their DNAs were denaturated and fixed to the membrane according to manufacturer's recommendations (GE Healthcare). Then, they were probed using a digoxigenin-labeled DNA probe specific for the blaVIM-1 gene, as previously described. 13 The number of blaVIM-1-negative colonies was obtained by subtracting from the total the number of the blaVIM-1-positive ones. The experiments were performed in triplicate, with three independent cultures. The number of generations (G) was calculated by the formula (log B − log A)/log 2, where A and B are the numbers of CFU/ml at the beginning of the culture period and at the end of the exponential phase, respectively. 3 The relative fitness of TS-832035 was calculated using the following formula: GTS-832035/GTS-103.
Statistical analysis
Differences in the growth kinetic data and the number of generations of TS-832035 and TS-103 were analyzed by the Student t test and a p-value of <0.05 was considered statistically significant.
Nucleotide sequence accession number
The nucleotide sequence of the variable region of the class 1 integron In90 carried by both TS-832035 and TS-103 was deposited in the GenBank/EMBL database with the accession number FJ495083.
Results
Genomic relatedness of TS-832035 and TS-103
In previous works, the P. aeruginosa clinical isolate TS-832035 was described as the first isolate of cluster A, a clonal lineage widely spread in an Italian hospital during a 3-year surveillance.13,14 It carried a chromosomal class 1 integron, named In70.2, whose variable region, ∼3,500 bp long, contained four gene cassettes (blaVIM-1, aacA4, aphA15, and aadA1). 25 Most of the isolates belonging to the cluster carried the same integron but few of them did not, although they appeared strictly related to the others after random amplification of polymorphic DNA typing and macrorestriction analysis. At the beginning of the present work, we investigated more accurately one of them, TS-103, in comparison with TS-832035, to support by additional techniques that they belong to the same clone and differ from each other by the presence or absence of the integron In70.2.
Compared by MLST, the two isolates revealed an identical allele profile, corresponding to ST235 (http://pubmlst.org/paeruginosa/). Other investigations confirmed the close relation of TS-103 with TS-832035: (1) no plasmids were ever detected in the total DNAs extracted from either of the two isolates (data not shown); (2) amplification with primers 5′-CS and 3′-CS showed that both isolates carried a class 1 integron, containing a variable region of ∼1,500 bp. After sequencing, this region appeared identical in both isolates and was deposited in GenBank under the accession number FJ495083. The integron was named In90 (Fig. 1) and carried a cassette array (aacA4, catB10-Ib) very similar to that already described in another Italian P. aeruginosa isolate (GenBank accession number AJ878850), differing for a G→A transition in codon 154 of the catB10 cassette, which gave rise to a new variant of this gene. So, although the detection of a common integron cannot be considered a typing method, this finding undoubtedly supports the clonality of the two isolates.
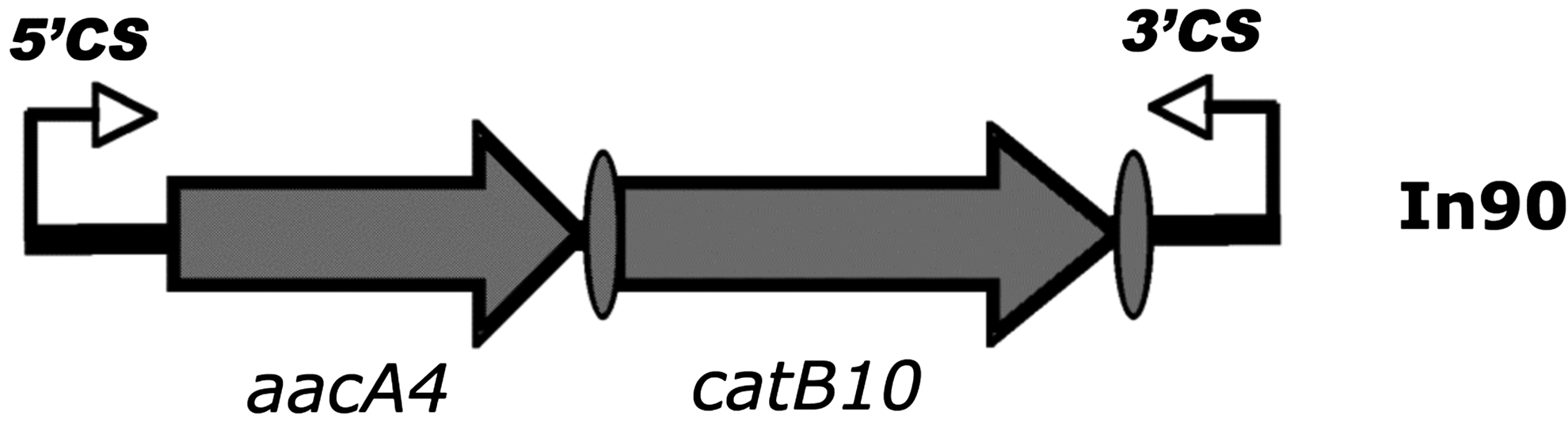
Structure of the variable region of the integron In90 from Pseudomonas aeruginosa TS-832035 and TS-103. Open reading frames are indicated by arrows; the attC recombination sites (59-base-element sequence) of gene cassettes are indicated by ovals.
On the other side, we confirmed the absence of the integron In70.2 in TS-103 by using PCR with couple of primers specific for the blaVIM-1, aphA15, and aadA1 gene cassettes. All the expected amplicons were obtained from DNA of TS-832035 but not from that of TS-103, and so we verified that TS-103 was lacking in the entire integron In70.2, not only in the blaVIM-1 cassette.
All these findings confirmed that TS-832035 and TS-103 really belong to the same clonal lineage but differ by the presence of the integron In70.2. Therefore, they were used to evaluate the fitness cost of this integron.
Antimicrobial agent susceptibility
TS-832035 confirmed the previously described higher resistance level to carbapenems compared with TS-103 13 (Table 1). The lack of the OprD porin observed in both isolates 8 explained the reason why TS-103 showed MICs equal to the resistance breakpoints, despite the absence of the carbapenemase activity. However, OprD loss does not confer resistance to extended-spectrum cephalosporins such as ceftazidime.
P. aeruginosa ATCC 27853 was included as a control.
CLOXA was added at 250 μg/ml.
IPM, imipenem; MEM, meropenem; CAZ, ceftazidime; CAZ + CLOXA, ceftazidime plus cloxacillin; AMK, amikacin; GEN, gentamicin; KAN, kanamycin; NET, netilmicin; SIS, sisomicin; SPT, spectinomycin; STR, streptomycin; TOB, tobramycin; MIC, minimal inhibitory concentration.
To verify if additional selective advantages were conferred to TS-832035 by the integron In70.2 or by other unidentified determinants, we investigated the susceptibility level of the two isolates to different antimicrobials: ceftazidime, in which resistance is conferred by the blaVIM-1 cassette; eight aminoglycosides, whose resistance determinants are carried in the cassette array of In70.2 25 ; three disinfectants: mercuric chloride, whose resistance determinants are frequently carried by mobile DNA sequences such as transposons, 12 to which In70.2 is associated 25 ; chlorhexidin and cetrimide, commonly used in the hospital wards.
As expected, TS-103 exhibited an MIC of ceftazidime much lower than TS-832035 but, as in the case of carbapenems, not so low to be considered clinically resistant. Moreover, its susceptibility increased fourfold in presence of cloxacillin (Table 1), suggesting that its resistance to ceftazidime might be due to the overexpression of some AmpC-type beta-lactamases. So this hypothesis was investigated (see below).
In the case of all the other antimicrobials tested, the MIC values of the two isolates were comparable (Tables 1 and 2). The resistance of TS-103 to so many aminoglycosides can be explained by the presence of the aacA4 determinant in the cassette array of the integron In90, carried by both isolates. It codes for an AAC(6′)-Ib variant enzyme, capable of acetylating kanamycin, 16 tobramycin, netylmicin, amikacin, and sisomicin. 29 Moreover, it shows a leucine-to-serine substitution in the catalytic site, which increases its affinity for gentamycin. 29 The molecular mechanisms leading to resistances toward the remaining aminoglycosides and the disinfectants were not further investigated. Actually, these resistances are not unusual in clinical isolates of P. aeruginosa.21,31 So, for the purpose of this work, the most important finding is that we could not find any phenotypic difference that could account for a real advantage to TS-832035 other than the higher resistance levels to carbapenems and ceftazidime, against which TS-103 still shows MICs equal to the clinical breakpoints.
P. aeruginosa ATCC 27853 was included as a control.
Identification of AmpC variants
As it is known that in P. aeruginosa the overexpression of some AmpC variants is associated with a decreased susceptibility to extended-spectrum cephalosporins such as ceftazidime, 26 we investigated the ampC gene of TS-103 to explain its resistance to ceftazidime. Sequencing analysis revealed that the ampC gene of TS-103 coded for the PDC-2 variant, known to confer reduced susceptibility to ceftazidime. 26
The ampC gene of TS-832035 was also investigated. It showed 100% homology to that of TS-103, further confirming the clonality of the two isolates.
Growth kinetics
The comparison between the growth kinetics of TS-832035 and TS-103 showed a different effect of the growth medium on the fitness of the two isolates (Fig. 2). In a rich broth such as LB (Fig. 2A), the two isolates exhibited an identical short lag phase and reached the same cell densities at the end of the log phase; so no differences were observed in their growth curves.
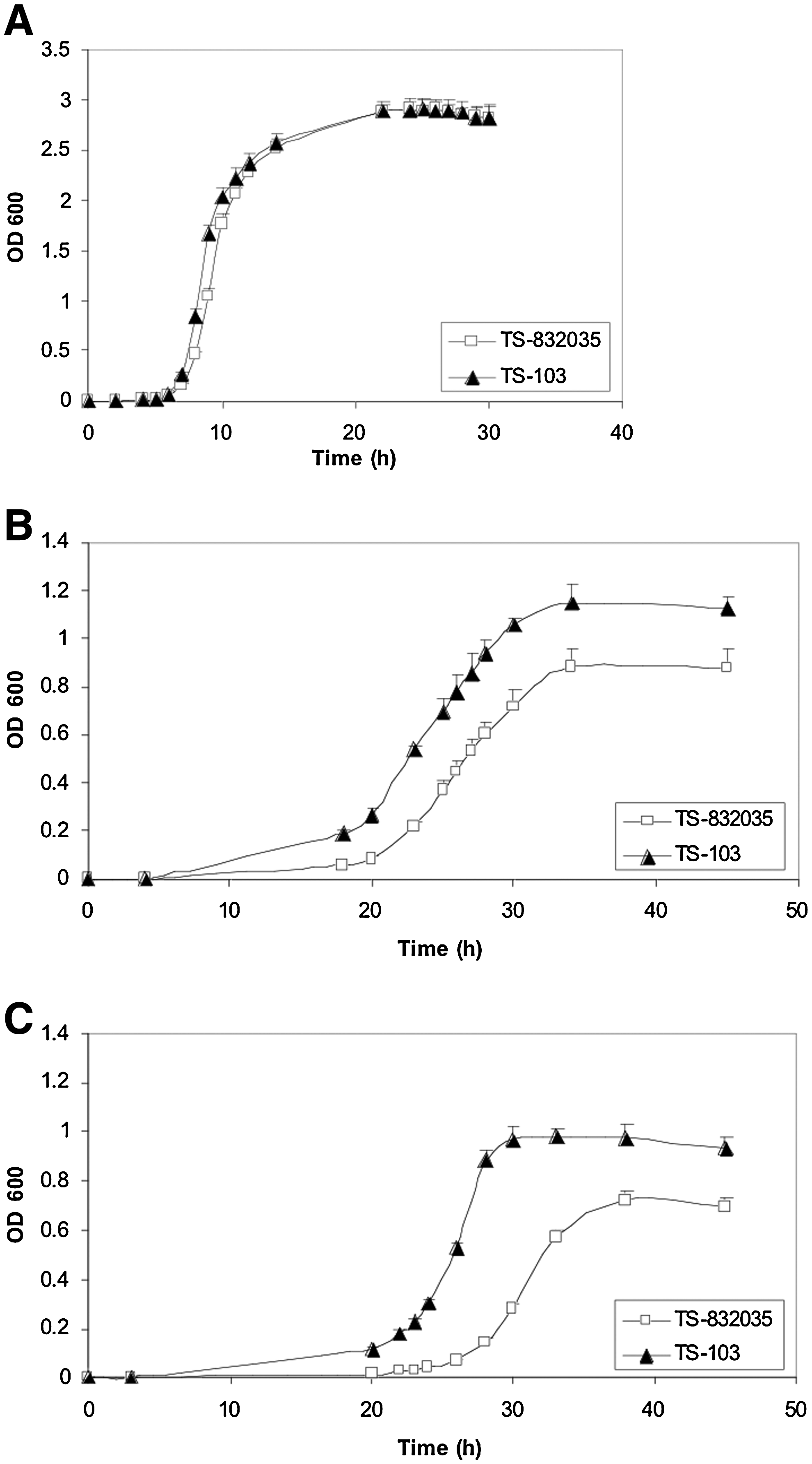
Growth kinetics of TS-832035 and TS-103 in LB broth (
In a minimal medium containing a rich carbon source such as glucose, both isolates showed a marked delay in the growth when compared with LB broth. However, TS-832035 exhibited a more extended lag phase and an overall reduction of the final cell density in stationary phase compared with TS-103. Moreover, the first showed OD600 measurements even two or three times lower than the latter and their differences were significant during the whole culture period (p always <0.05) (Fig. 2B). This effect was more evident in a poorer broth such as minimal medium supplemented with sodium acetate, where TS-832035 showed the longest lag phase, exhibited the lowest cell density in stationary phase, and achieved the highest reductions in growth values compared with TS-103, with very significant differences (p < 0.01) at any time point (Fig. 2C). In Table 3, the number of hours necessary to the two isolates to reach the end of the exponential phase in each of the three media and the respective OD600 values are reported. These data show that in LB the time employed by the two isolates to reach the end of the exponential phase was about half compared with the minimal medium, and the growth of TS-832035 was particularly limited in the poorest medium, where the end of the exponential phase was even delayed compared with TS-103. Moreover, the lower population density reached by TS-832035 in the stationary phase of a poor medium culture is particularly noteworthy, as it suggests that this isolate needs more nutrients for replicating itself compared with TS-103.
Results are expressed as mean OD600 ± SD of six experiments.
p < 0.04.
p < 0.03.
MM, minimal medium; LB, Luria Bertani.
Competition assays
To determine if the integron In70.2 affected the bacterial fitness, competition assays of TS-832035 against TS-103 were performed. A lower number of CFU/ml of TS-832035 (p always <0.05) was detected during the whole experimental period and in each culture, even in that grown in LB (Fig. 3 and Supplemental Table 1 [available online at www.liebertonline.com]). So, at the end of the exponential phase, this resulted in a lower number of generations formed by TS-832035; therefore, the relative fitness was lower than 1 in all the media used (Table 4). In agreement with the results of the growth kinetics, both isolates grew faster in LB than in the other two media and the cost of the integron appeared different depending on the nutrient availability. It was maximal when TS-832035 had to compete with TS-103 in a poor medium (where the final cell concentration of the first resulted 1 log10 lower than that of the latter) (Fig. 3C), whereas it was alleviated in richer media (0.6 log10 lower, only) (Fig. 3A, B). However, in each of the competition assays, TS-832035 showed a persistent fitness cost associated to the integron that limited significantly its ability to compete with TS-103 (Table 4).

Competition assays between TS-832035 and TS-103 in LB broth (
The number of generations (G) was calculated as described in the Materials and Methods section and expressed as mean ± SD of three independent experiments.
p < 0.04.
p < 0.02.
p < 0.03.
To investigate the influence of subinhibitory drug concentrations on the growth ability of TS-832035 and TS-103, a competition assay in LB broth with imipenem 2, 4, and 8 μg/ml was performed. As expected, growth of TS-832035 was not affected at all by addition of imipenem concentrations much lower than its MIC. On the contrary, TS-103 was partially influenced by drug concentrations very close to its MIC. At the end of the log phase, its cell density was halved at any doubling of imipenem concentration (2 × 109 CFU/ml, 109 CFU/ml, 5 × 108 CFU/ml, and 2.5 × 108 CFU/ml at 0, 2, 4, and 8 μg/ml imipenem, respectively). This result suggests that TS-832035, because of its higher resistance level, might fit better than its less-resistant counterpart in the presence of subinhibitory concentrations of the drug.
Discussion
Since 1999, a rapid spread of MDR P. aeruginosa strains producing VIM-type MBLs had occurred at the Trieste University Hospital (northeastern Italy). The characterization of 128 carbapenem-resistant P. aeruginosa isolates, collected from patients admitted to that hospital between 1999 and 2002, 13 showed that most of them produced the VIM-1 enzyme and belonged to a common clonal lineage, widely distributed in that hospital and even outside of it. 14 Molecular characterization of TS-832035, the first VIM-1–producing isolate detected in 1999, showed that the blaVIM-1 cassette was carried by a chromosomal class 1 integron, named In70.2. 25 Few VIM-negative carbapenem-resistant P. aeruginosa isolates, belonging to the same cluster, were detected in the hospital during the same period, but epidemiologic investigations revealed the constant prevalence of the VIM-1–positive strains during the 3 considered years. 13 The reasons of such a massive diffusion of the VIM-1–producing isolates are unknown.
The presence of an integron requires the bacteria to synthesize additional nucleic acids and proteins; this causes an increase of the metabolic burden on the organism, 17 which should result in a fitness cost easily detectable in competition assays, under nonselective conditions, between an integron-containing isolate and its integron-negative counterpart. Yet, an evaluation of this type has never been performed, probably because of the difficulties to obtain an isogenic couple of strains differing for a chromosomal integron. In fact, a couple of isogenic strains differing for an integron carried by a plasmid would probably be easier to construct, but the evaluation of its fitness cost would be affected by additional variables (e.g., loss of the plasmid in the absence of selective pressure, copy number, etc.). With respect to this, the couple of isolates described above was compared by different typing techniques, really highlighting a lot of common features except for the presence of In70.2. For this reason, although it is obvious that only the whole-genome sequencing could confirm a perfect isogenicity, we considered the two isolates sufficiently closely related, to ascribe their different fitness to the presence of the integron In70.2. Our in vitro experiments, carried out under permissive conditions, showed that (1) in separate cultures, the fitness cost of TS-832035 was significantly higher than TS-103 in minimal medium containing either of two different carbon sources and the disadvantage of TS-832035 increased in the presence of the poorer carbon source; by contrast, no differences in biological fitness of the two isolates were observed in a rich medium, such as LB; (2) in competition assays, the biological fitness of TS-832035 was more impaired than that of TS-103 even when they were cultured in the rich medium, although the difference remained more evident as the isolates were turned to a medium poorer in nutrients. Therefore, the fitness loss of the integron In70.2-positive P. aeruginosa isolate, TS-832035, resulting in an impaired ability to compete with its integron In70.2-negative counterpart, TS-103, particularly under carbon source limitations, demonstrated a significant disadvantage of the former compared with the latter, mainly under stressful in vitro conditions. To evaluate the significance of these results, we cannot ignore that it is still debated how well the growth ability measured in laboratory settings correlates to fitness in real life. 1 The overall fitness of a resistant bacterium is influenced by a lot of different factors, such as impact of resistance on transmission, 1 competition with the normal human habitat, and exposition to natural environmental factors. 11 Moreover, different obstacles have been postulated to affect the translation machinery under in vitro or in vivo conditions.2,20 However, recent comparisons among in vitro and in vivo assays suggested that the measurements obtained in vitro reflected adequately the ones obtained in vivo, and that it is reasonable to assume that if a cost is seen in vitro it will also exist in vivo, where conditions are typically more stressful.11,27 As a reduction of fitness of TS-832035 compared with TS-103 was actually observed in our in vitro assays, the reasons for the prevalence of the blaVIM-1-positive over the blaVIM-1-negative isolates in the Trieste University Hospital during the 3 years of surveillance remain difficult to explain. The hypothesis that the carriage of the integron could not be due to a possible disuse of carbapenem antibiotics that made unnecessary the expression of the resistance, as previously described for tetracycline, 23 is not reliable as the expression of the integron cassettes are usually constitutive. 19 Besides, our in vitro assays, showing a loss of the fitness of the isolate carrying the integron, were performed under permissive conditions. Trying to explain this lack of correlation between the epidemiological data and the results of the in vitro experiments, we evaluated the hypothesis that at least one of the resistance determinants carried by or linked to integron In70.2 confers a selective advantage, making its host more suitable to spread in a hospital setting than its In70.2-negative counterpart. The attempt to show that the advantage of these isolates was connected to a higher resistance to the aminoglycosides, whose resistance determinants are present in the In70.2 cassette array, 25 or to a higher tolerance to common disinfectant compounds failed. In fact, both TS-832035 and TS-103 isolates showed the same MIC values. The only phenotypic differences we were able to detect were the higher resistance levels of TS-832035 to carbapenems and ceftazidime compared with that of TS-103, although both of them appeared resistant to the doses of drugs used in clinical praxis, because of the lack of OprD porin 8 and the presence of the PDC-2 variant of the AmpC beta lactamase, 26 respectively. Negri et al. 22 showed that within a short range of subinhibitory concentrations of an antibiotic, which they call “selective windows,” the isolate with higher MIC fits better than its hardly less-resistant counterpart and suggested that, during antibiotic therapy, gradients of drug concentrations are formed in human body creating compartments where “selective windows” are present. So, even if we cannot exclude the existence of any compensatory mutation responsible for a better in vivo fitness of TS-832035, the evidence that TS-103 growth was impaired by subinhibitory imipenem concentrations supports the hypothesis that in a hospital setting, where bacteria are constantly exposed to antibiotics, even sometimes to those they are resistant (e.g., for multiple infections therapy), the isolates with a higher resistance level prevail over those with MIC equal or just a little higher to the breakpoints.
The spreading ability of this MDR clonal lineage was evident since its first detections. In fact, VIM-1–positive P. aeruginosa isolates, PPV-108 and VR-143/97, strictly related to TS-832035, were detected in other Italian hospitals in the same period, either as sporadical isolates or causing small outbreaks. 25 This relatedness has been also supported by our MLST results. The isolates TS-832035 and TS-103 belong to ST235, which differs from ST227, to which belong PPV-108 and VR-143/97, 10 for only one point mutation. Moreover, a blaVIM-1-carrying integron named In71, nearly identical to In70.2, was recently detected in an Enterobacter cloacae isolated from a blood culture of a patient hospitalized in Genoa. 4 This finding is particularly worrying, for the risk that integron-carrying isolates belonging to different bacterial families might be able to get ahead even of strains exhibiting MIC values equal or a little higher than the breakpoint, selecting MDR strains with higher and higher resistance levels. So, the significant fitness cost associated with the integron found in our assays might be considered a possible soft spot that should be taken into account to contrast the spreading ability of integron-carrying strains.
Footnotes
Acknowledgments
This work was supported by grants from Italian M.I.U.R. (PRIN 2007, nos. 20077HCK77_003).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.