
Abstract
The acquisition of β-lactamases, such as class B metallo-β-lactamases, in Pseudomonas aeruginosa is detrimental to antimicrobial therapy in hospitalized patients. In Mexico, metallo-β-lactamase IMP-15 has been found to be encoded on the In95 class 1 integron in a major clone of P. aeruginosa. In this work, we describe the variability of this class 1 integron in an epidemic clone of carbapenem-resistant P. aeruginosa clinical isolates highly related to isolates previously described in Mexico.
A total of 117 nonduplicate P. aeruginosa isolates were obtained at HIM “Eva Sámano de López Mateos,” Michoacán, Mexico, between August, 2004 and July, 2006. All isolates were initially tested for imipenem resistance using the Kirby Bauer method; 31 (26.5%) of them turned out to be resistant to imipenem, according to the Clinical and Laboratory Standards Institute (CLSI) recommendations. 1 The antimicrobial susceptibility test was carried out by agar dilution, and the results were interpreted according to the CLSI guidelines. The resistance profile was as follows: imipenem, 100%; meropenem, 65%; ceftazidime, 74%; aztreonam 45%; amikacin and ciprofloxacin, 59%; gentamicin, 42%; and piperacillin and piperacillin-tazobactam, 6%.
All of the 31 carbapenem-resistant isolates (≥8 μg/ml) were tested for pulsed-field gel electrophoresis and screened for metallo-β-lactamase (MβL) production by double-disk synergy test, 9 Etest® strips (AB BIODISK, Solna, Sweden). A polymerase chain reaction (PCR) amplification mediated specific primers for blaVIM, blaIMP, blaSPM and blaGIM was performed in all isolates.4,5 Seventeen of them corresponded to a major clone (B) with five subtypes (B1–B5), and 18 isolates, including the major clone, were positive for MβL phenotype production and yielded positive amplicons with the blaIMP specific primers. Analysis of the sequence from all amplicons showed the blaIMP-15 gene. To identify the number and sizes of class 1 integron encoded in each isolate, a PCR amplification assay was performed using primers 5′CS and 3′CS described by Levesque. 10 In each clinical isolate, between 1 and 4 different sizes of class 1 integrons were identified with values from 4.8 to 1.5 kb (Table 1). To identify which of the class 1 integrons contained the blaIMP-15 gene, a Southern blot hybridization was performed with all PCR products and a blaIMP-15 probe. 5 The results indicated that all PCR products of 4.8, 2.2, 4.4, and 4.2 kb and one 1.4 kb harbored the blaIMP-15 gene. Moreover, three isolates (3530, 3584, and 3587) that encoded one additional blaIMP-15 copy with a different molecular weight class 1 integron were identified (Table 1 and Supplemental Fig. 1, available online at www.liebertonline.com). All class 1 integrons encoding the blaIMP-15 gene were digested with DraIII restriction endonuclease. An identical restriction profile was obtained from each amplification product, confirming the presence of the same class 1 integron structure in each of them (data not shown).
The amplification product underlines harboring the blaIMP-15 gene, identified by Southern hybridization and was also screened for class 1 integron using specific primers for 5′ and 3′ CSs. 10
4663 and 4703 strains contain two additional plasmids of 1 and 3 kb.
PCR, polymerase chain reaction; CS, conserved sequence; MIC, minimum inhibitory concentration; AMK, amikacin; CAZ, ceftazidime; IMI, imipenem; MER, meropenem; ATM, aztreonam; PIP, piperacillin; PIP/TAZ, piperacillin–tazobactam; GEN, gentamicin; CIP, ciprofloxacin; IUC, insensitive care unit; Qr, Quirurgical; Onc, oncology; Mat, maternal; Urg, urgency, UI, unavailable information.
Five representative class 1 integrons containing blaIMP-15 were selected for further characterization: They were 4.8 kb (In95-1-like), 4.4 kb (In95-2-like), 4.2 kb (In95-3 like), 2.2 kb (In204), and 1.4 kb (In205) in length, and they corresponded to strains 6805, 3530, 3599, 3534, and 3587, respectively (Table 1 and Fig. 1). To characterize the variable region from each class 1 integron, a primer combination was used, as previously described. 5 All class 1 integron structures were determined by examining overlapping PCR fragments upstream and the blaIMP-15 gene downstream; this yielded a continuous sequence. The PCR products were sequenced by means of the chain termination method with a Big-Dye Terminator kit (Applied Biosystems, Foster City, CA) and analyzed on an ABIPRISMA 3100 (Applied Biosystems); the nucleotide sequences were assembled using the VectorNTI software. The structure of each one of these class 1 integrons is shown in Fig. 1. The class 1 integrons with molecular weight of 4.8, 4.4, and 4.2 showed a highly similar gene array to that of the In95 previously identified in P. aeruginosa isolates 4677 and 1750J from the HCG in Mexico and the University of Kentucky HealthCare (UKUC) in the United States; consequently, they were denoted as In95-1, In95-2, and In95-3-like class 1 integrons, respectively (Fig. 1). The In95-1-like (4.8 kb) class 1 integron was identified in 12 (66%) out of 18 isolates, and it contained six gene cassettes within its variable region. The In95-2-like (4.4 kb) and In95-3 like (4.2 kb) class 1 integrons were identified in 1 (5%) out of 18 isolates, and both had a similar gene array to that of the In95-1-like integron, with the exception of the qacH and blaOXA-2 genes. Regarding the other two class 1 integrons, In204 (2.2 kb) and In205 (1.4 kb) included the blaIMP-15 in the first position followed by aadA6, ORFD, and qacH (Fig. 1). These class 1 integrons were identified in 6 (33%) and 1 (5%) out of 18 strains, respectively. The 1.4 kb class 1 integron that did not encode the blaIMP-15 gene was identified in 10 out of 18 isolates. It was analyzed using the DraIII restriction enzyme. The results showed an identical restriction profile to the one previously identified in HCG isolates (data not shown), and its structure corresponded to that of In51, which has been described elsewhere. 12 The GenBank/EMBL accession numbers for the sequence data reported in this paper are In95-1, GQ856538; In95-2, GQ856539; In95-3, GQ856540; In204, GQ856541; and In205, GQ856542.
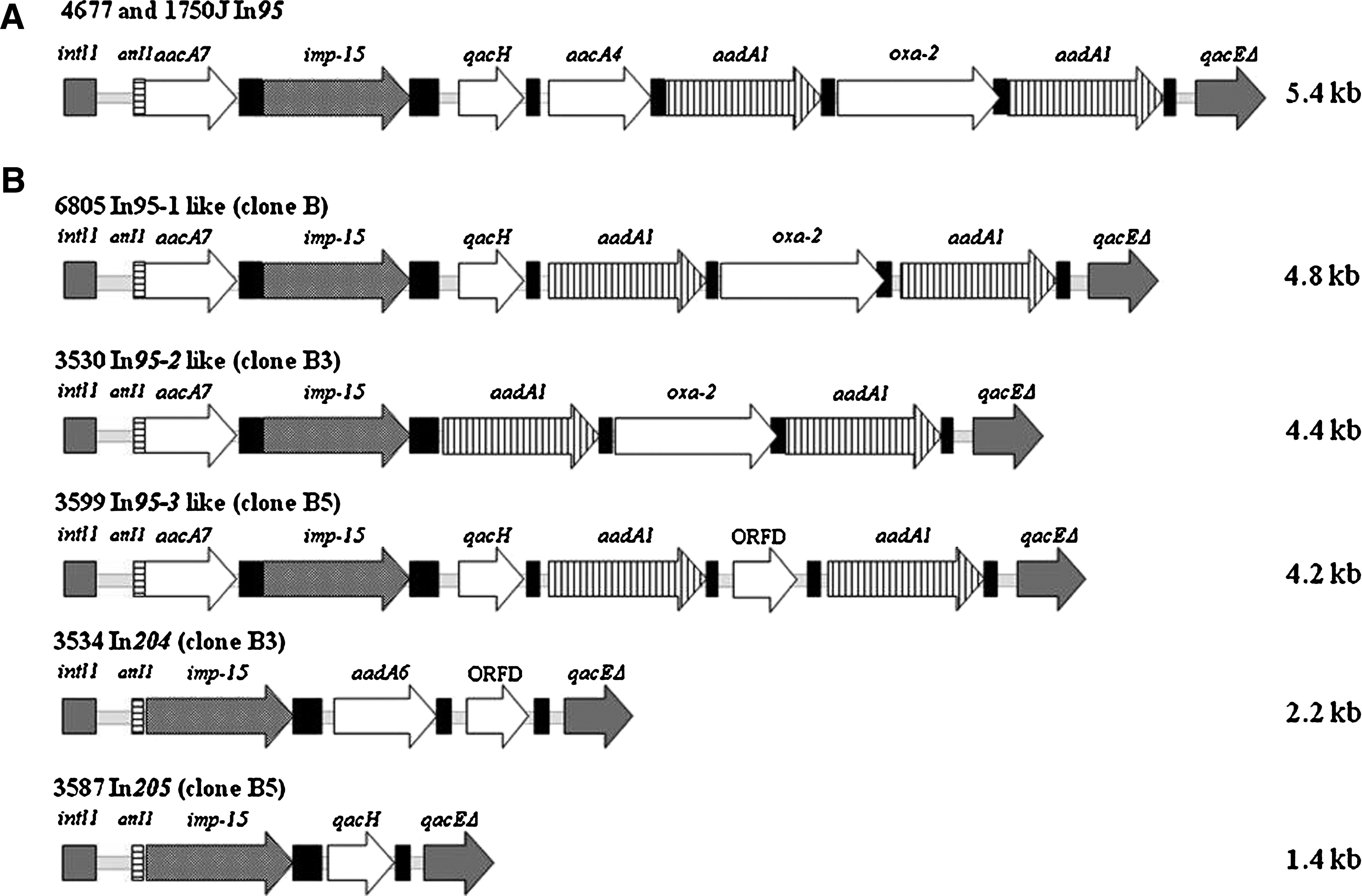
Schematic representation of the blaIMP-15-containing class 1 integrons. (
To determine if the plasmid-borne class 1 integrons from the P. aeruginosa clinical isolates contained the blaIMP-15 gene, a Southern hybridization was performed using the blaIMP-15 as a probe. The results indicated a positive signal with the 30 kb plasmid in all the strains from HIM, including isolate 4677 from HCG. In addition, two isolates (6805 and 3584) harbored a second plasmid, of 80 and <40 kb, respectively, which did not give a positive signal (Table 1). To identify a possible genetic relation between the 30 kb plasmids from the P. aeruginosa clinical isolates from HIM and HCG, a plasmid restriction fragment length polymorphism (RFLP) was performed using DraI enzyme. The restriction pattern was analyzed using the GelCompar II software (Applied Math, Kortrijk, Belgium). The similarity percentage was determined by means of a dendrogram derived from Unweighted Pair Group Method with Arithmetic Mean (UPGMA) and Dice coefficients. The results indicated a high similarity (>80%) between the blaIMP-15-bearing plasmids collected at both hospitals (data not shown). The attempts to transfer resistance by conjugation or transformation of HIM plasmid DNA into E. coli J52 and P. aeruginosa PU21 as recipient strains were unsuccessful under the conditions assayed, as previously reported in the case of the P. aeruginosa isolate from HCG. 5 This suggests that these plasmids have a similar molecular structure, which indicates the restriction pattern obtained by RFLP.
In addition, we wanted to identify a possible genetic relation between the P. aeruginosa clinical isolates obtained at both hospitals. A pulsed-field gel electrophoresis was performed including representative P. aeruginosa clones and their respective subtypes from HIM (clone B and five subtypes) and from HCG (clone A and two subtypes). Electrophoretic patterns were analyzed using the GelCompar II software. The similarity percentages were identified by means of a dendrogram derived from UPGMA and Dice coefficients (Band position tolerance and optimization were set at 1.8% and 0.9%, respectively). The representative HIM isolates subtype B5 (isolate 3599) clustered with the representative HCG isolates subtypes A1 and A2 (isolates 4659 and 4658) with a similarity coefficient of 100%, and all of these isolates displayed a similarity coefficient of 94.1% with regard to clone A (strain 4677) from HCG. In addition, clone B isolates and the three subtypes (isolates 3530, 3584, 3583, 3588, 3585, and 6805) obtained at HIM clustered with clone A from HCG and the two subtypes with a similarity coefficient of 82.4% (Fig. 2). These results indicate a clonal dissemination of P. aeruginosa at both medical centers. On the other hand, clone C (isolate 3587) from HIM was not related to these isolates.
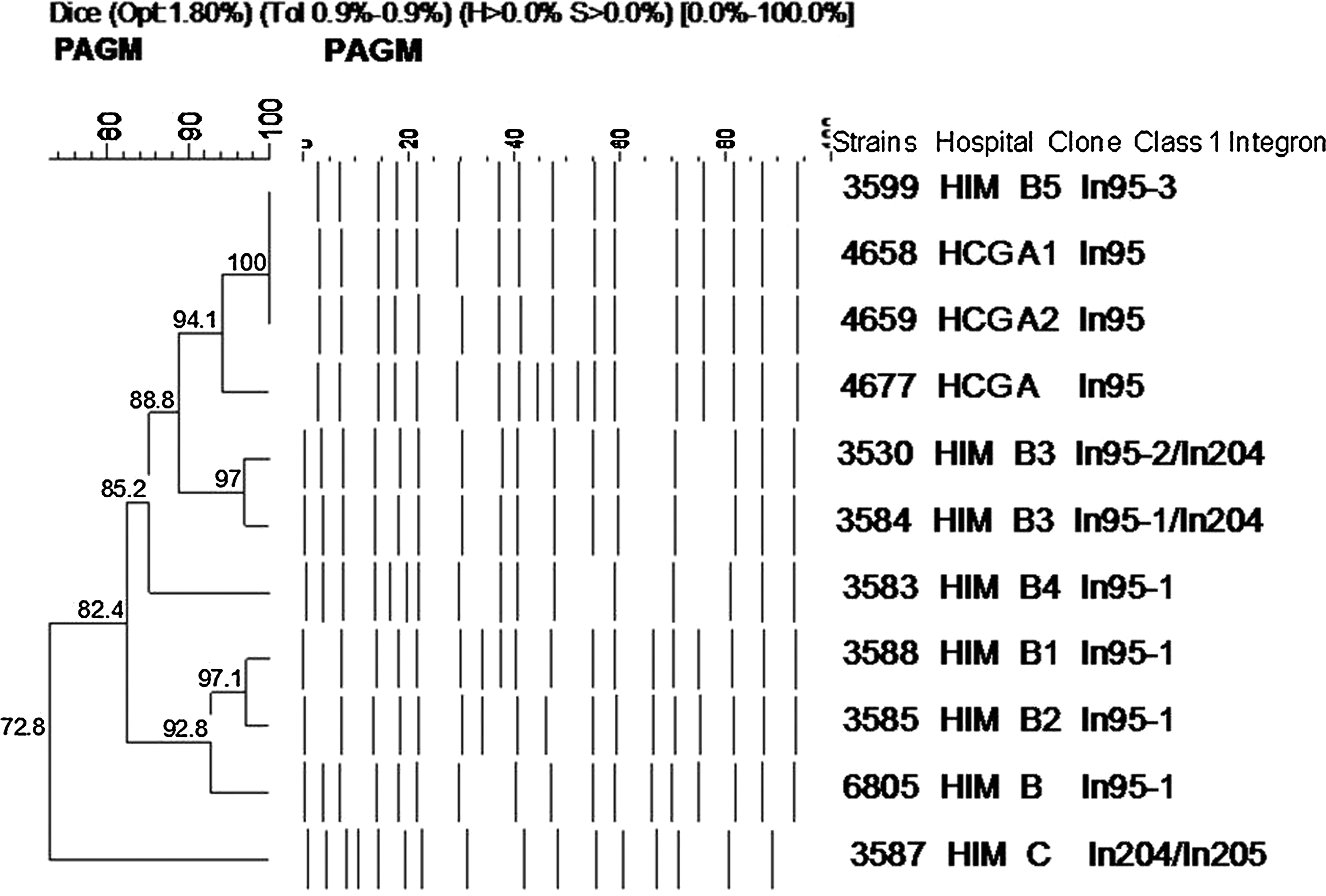
Dendrogram of pulsed-field gel electrophoresis using SmaI macrorestriction fragment of representative P. aeruginosa isolates belonging to the major clones from HIM and HCG. For this study, isolates showing a similarity coefficient of >80% were considered as genetically related. Isolate number, medical center, clone designation, and class 1 integron(s) contained are indicated. HIM, Hospital Infantil de Morelia; HCG, Hospital Civil de Guadalajara; PAGM, P. aeruginosa Guadalajara/Morelia.
Clonal dissemination of P. aeruginosa isolates resistant to most commercially available antibiotics has been described in several countries, and it represents a public health problem.7,8 Studies on carbapenem resistance clinical isolates in Mexico are limited. Nevertheless, the National Institute of Respiratory Diseases (NIRD) in Mexico City reported a 20% prevalence of carbapenem-resistant P. aeruginosa isolates and a 3.4% prevalence of MßL-producing strains, including types VIM-2 and IMP-15. 13 Therefore, MβL screening methods should be implemented, especially in hospitals where P. aeruginosa is an endemic organism. 14
The blaIMP-15 enzyme is a broad-spectrum MβL. It was identified in P. aeruginosa isolates from HCG, UKUC, and NIRD in 2003, 2005, and 2004, respectively. In all these cases, the blaIMP-15 gene is encoded on a class 1 integron. Nevertheless, a class 1 integron with a different gene array was previously described for blaIMP-15 in Thailand, 11 as is the case for In204 and In205 class 1 integrons in the present study. Up to now, the blaIMP gene has been identified on integron-borne gene cassettes, which display a variable region. This allows to consider the presence of an integron recombination system that could account for its rapid dissemination in clinical settings under the selective pressure generated by antimicrobial agents.2,3 The finding of variability in the case of the In95 class 1 integron harboring the blaIMP-15 gene cassette in highly related clones underscores the potential for gene-cassette recombination (acquired and release genes) within this class 1 integron. This is supported by the high similarity among the plasmids, identified by the RFLP pattern of the nonconjugative plasmids harbored in clinical isolates from HIM and HCG.
Footnotes
Acknowledgments
This work was supported by grants 37195-M from CONACYT and SALUD-2003-C01-009. The authors want to thank Fatima Alvarez-Guillen for reviewing the manuscript. This work was presented in part at the 48th Interscience Conference on Antimicrobial Agents and Chemotherapy held in Washington, DC, from October 24 to October 28, 2008. The sequence data reported in this paper appear in the GenBank/EMBL nucleotide database under accession numbers In95-1, GQ856538; In95-2, GQ856539; In95-3, GQ856540; In204, GQ856541; and In205, GQ856542.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.