
Abstract
The genus Corynebacterium includes a high number of species that are usually isolated from human skin as saprophytes. However, these microorganisms have also been reported as infectious agents in a broad group of patients and have showed broad-spectrum resistance. We studied the susceptibility profiles against macrolides, clindamycin, and streptogramins of 254 clinical strains belonging to the species Corynebacterium urealyticum (120), Corynebacterium amycolatum (66), Corynebacterium jeikeium (17), Corynebacterium striatum (20), Corynebacterium coyleae (12), Corynebacterium aurimucosum (11), and Corynebacterium afermentans subsp. afermentans (8). The MLSB phenotype was detected in 186 strains and was associated with the presence of methylase enzymes codified by the erm(X) gene in 171 strains. The erm(B) gene was only detected in two C. urealyticum strains. Fourteen strains showed macrolide resistance, but they did not carry erm genes. mef genes were not detected despite eight C. amycolatum strains showed the M phenotype. Also, the presence of hydrolytic enzymes codified by ere(B) was evaluated, but all results were negative. Resistance to macrolide in Corynebacterium sp. is mainly due to the presence of erm(X) methylase, although other resistance mechanisms could be involved.
Introduction
The high level of macrolide resistance is a matter of particular concern. Macrolides are frequently used in infections that can be caused by these microorganisms, and are an alternative to β-lactams and tetracyclines. 33 Macrolide resistance is mainly due to methylase enzymes that are codified by erm genes. 22 Other resistance mechanisms such as mutations in 23S rRNA domain V, modifications of proteins L4 and L22, efflux pump systems, and hydrolytic enzymes have been reported in different genera.18,30 Despite all these data, only a few reports have studied this topic in Corynebacterium sp.23,32 The objective of this study was to evaluate the mechanisms implicated in macrolide resistance among clinical isolates of Corynebacterium species.
Materials and Methods
Bacterial strains
Bacterial strains were isolated from clinical samples of patients with infection and from a skin colonization study in hospitalized patients 28 presented at the Hospital Fundación Jiménez Díaz de Madrid, Spain, and the Complejo Asistencial de León, Spain, between January 1985 and December 2005. In this study, one isolate per patient was used. The strains were identified by the API Coryne System (bioMérieux, Marcy l′Etoile, France) using the instructions provided by the manufacturer. 14 All the strains belong to the species C. amycolatum, C. striatum, Corynebacterium coyleae, Corynebacterium aurimucosum, and Corynebacterium afermentans subsp. afermentans, as well as a randomly selected strains from the species C. urealyticum and C. jeikeium, and were also identified by 16S rDNA sequence analysis using the protocol described by Fernández-Natal et al. 10 Strains were kept frozen in skim milk at −20°C.
Antimicrobial susceptibility
In vitro susceptibility testing against erythromycin, clarithromycin, azithromycin, clindamycin, and quinupristin/dalfopristin was conducted using E-test strips (AB, Biodisk, Solna, Sweden) using the previously described protocol. 11 The susceptibility of spiramycin was studied using commercial disks of 100 μg (bioMérieux). All the studies were performed on Mueller–Hinton agar supplemented with 5% sheep blood in a normal atmosphere at 37°C during 24–48 hours.
Susceptibility to erythromycin, clindamycin, and quinupristin/dalfopristin was assessed using the breakpoints described for Corynebacterium sp. by the Clinical and Laboratory Standards Institute (CLSI). 4 To analyze the susceptibility of clarithromycin, azithromycin, and spiramycin, breakpoints reported for Staphylococcus sp. were used. 5 Staphylococus aureus ATCC 29213 and ATCC 25923 collection strains were used for quality control purposes.
Detection of the resistance genes
The strains were grown on blood agar plates during 24–48 hours at 37°C in normal atmosphere and checked for purity. One loop was resuspended in 200 μl of sterile distilled water. Suspension was boiled during 20 minutes at 100°C and subsequently centrifuged at 12,000 rpm for 15 minutes. Supernatant (150 μl) was used for PCR studies.
PCR was performed using commercial puRe Taq Ready-To-Go PCR Beads® (Amersham Biosciences, Buckinghamshire, United Kingdom). The primers used for detection of the erm(X) gene were Cerm 1 (5′-GAC ACG GCC GTC ACG AGC AT-3′) and Cerm 2 (5′-GGC GGC GAG CGA CTT CC-3′); for the erm(A) gene, Erm A-1 (5′-TCT AAA AAG CAT GTA AAA GAA-3′) and Erm A-2 (5′-CTT CGA TAG TTT ATT AAT AGT-3′); for the erm(B) gene, Erm B-1 (5′-GAA AAG GTA CTC AAC CAA ATA-3′) and Erm B-2 (5′-AGT AAC GGT ACT TAA ATT GTT TAC-3′); and for the mef(A-E) gene, Mef A-E-1 (5′-AGT ATC ATT ATT CAC TAG TGC-3′) and Mef A-E-2 (5′-TTC TTC TGG TAC TAA AAG TGG-3′). The erm(X) gene amplification was performed according to the protocol described by Rosato et al. 23 For all other genes we applied PCR conditions previously reported for Streptococcus sp. 29 The amplification products were detected by electrophoresis in 3% agarose. The size of the amplicons was estimated using Biogen software (Vilber-Lourmat, Marne-la-Vallée, France) compared with a commercial molecular weight ladder (Bio Rad Laboratories, Hercules, CA). The erm(X) and erm(B) amplicons were subsequently purified and sequenced with the ABI Prism 377 Big Dye Terminator kit using the primers Cerm 1, Cerm 2, Erm B-1, and Erm B-2. 10 Nucleotide sequences were analyzed with BLAST software and compared with previously reported sequences (GenBank, NCBI).
Erythromycin enzymatic hydrolysis
To determine the presence of the ere(B) gene and its hydrolytic effect over erythromycin, we have used the protocol described by Wondrack et al. 31 The growth of the erythromycin-sensitive S. aureus ATCC 25923 strain was evaluated in blood-agar plates after 24 hours at 37°C in a normal atmosphere with erythromycin disks of 40 and 400 mg/L. Previously, the disks and a suspension of different Corynebacterium strains were incubated together to evaluate the hydrolysis of this antibiotic. The Escherichia coli ClaNal/pAT172 (pUC8ΩereB) strain was used as a positive control. This strain has a plasmid that inactives the erythromycin and was provided by Dr. Courvalin (Pasteur Institute, Paris, France). Distilled water was used as a negative control.
Statistical analysis
Erythromycin susceptibility and resistance rates among Corynebacterium species were analyzed by means of exact Fisher test. All statistical calculations were performed using SPSS 10.0 Software (SPSS, Inc., Chicago, IL).
Results
Two hundred fifty-four Corynebacterium spp. strains were included in this study: 120 C. urealyticum, 66 C. amycolatum, 20 C. striatum, 17 C. jeikeium, 12 C. coyleae, 11 C. aurimucosum, and 8 C. afermentans subsp. afermentans. The results of the in vitro susceptibility study are shown in Table 1. Among the macrolide-resistant strains, 186 showed an MLSB phenotype, with a high level of resistance against macrolides and clindamycin, while 8 strains showed the M phenotype, with low level resistance against 14- and 15-membered macrolides but susceptibility to 16-membered macrolide, clindamycin, and quinupristin-dalfopristin.
S, susceptible; I, intermediate; R, resistant.
The amplification product of the erm(X) gene was detected in 175 strains: 171 showed MLSB resistance, 2 strains (one C. striatum and one C. aurimucosum strains) were susceptible against all studied antibiotics, and 2 C. amycolatum strains were resistant to erythromycin, claritromycin, and azithromycin but susceptible against spiramycin, clindamycin, and quinupristin/dalfopristin (M phenotype). No amplification products were detected in 14 strains, although they showed resistance to macrolide and clindamycin. There were also six C. amycolatum strains showing the M phenotype and without any of the erm amplicons studied. The amplification product of erm(B) gene was detected in two C. urealyticum strains, and in one of them both erm(X) and erm(B) amplicons were detected. The erm(A) and mef(A-E) amplification products were not detected in any of the studied Corynebacterium strains (Table 2).
One strain carried the genes erm(X) and erm(B) simultaneously.
Two strains showed the M phenotype despite carrying the erm(X).
One strain was susceptible despite being erm(X) gene positive.
The C. striatum strains were more susceptible against erythromycin compared with all other species, although this difference was only statistically significant against C. amycolatum strains (p = 0.011, Fisher's exact test), but not against C. urealyticum strains (p = 0.103) or C. jeikeium strains (p = 0.3). The number of C. coyleae, C. aurimucosum, and C. afermentans strains was too low for a proper statistical analysis.
The erm(X) amplicon sequence was over 390 pair of bases and showed a nucleotide identity between 90% and 99% with the previously described sequence (Fig. 1). The erm(B) amplicon sequence of C. urealyticum was 639 pair of bases, showing a nucleotide identity of 99% with respect to other erm(B) amplicon sequences from Streptococcus and Enterococcus species (Fig. 2).
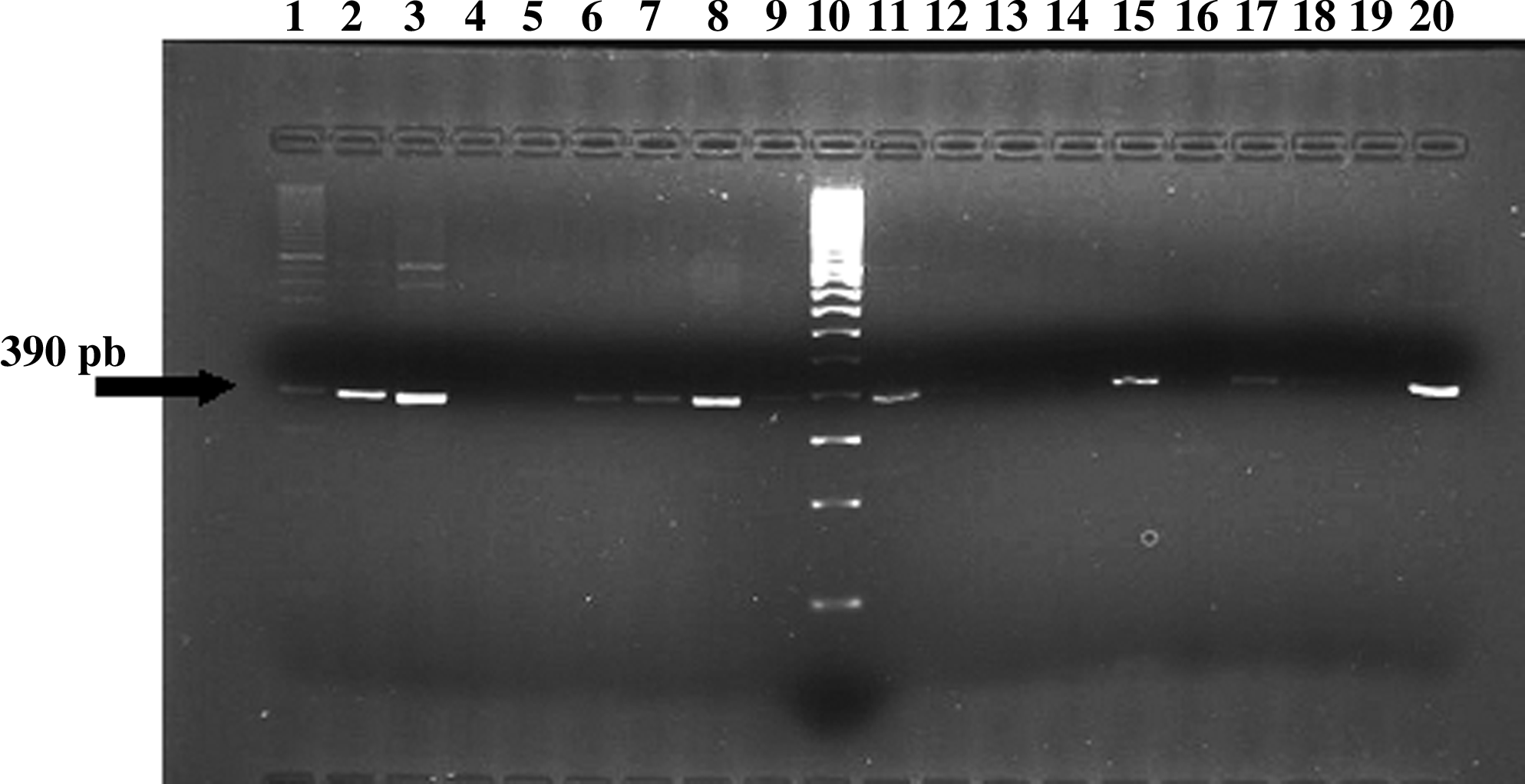
Analysis of erm(X) determinant in Corynebacterium urealyticum. PCR products of amplification using primers Cerm 1 and Cerm 2. Lane 1, strain A-6; lane 2, strain A-7; lane 3, strain A-8; lane 4, strain A-9; lane 5, strain A-10; lane 6, strain A-11; lane 7; strain A-12; lane 8, strain A-13; lane 9, strain A-14; lane 10, DNA ladder; lane 11, strain A-15; lane 12, strain A-16; lane 13, strain A-17; lane 14, strain A-18; lane 15, strain A-20; lane 16, strain A-21; lane 17, strain A-22; lane 18, strain A-23; lane 19, distilled water (negative control); lane 20, Corynebacterium jeikeium CB-396 (positive control). pb, pair of bases.
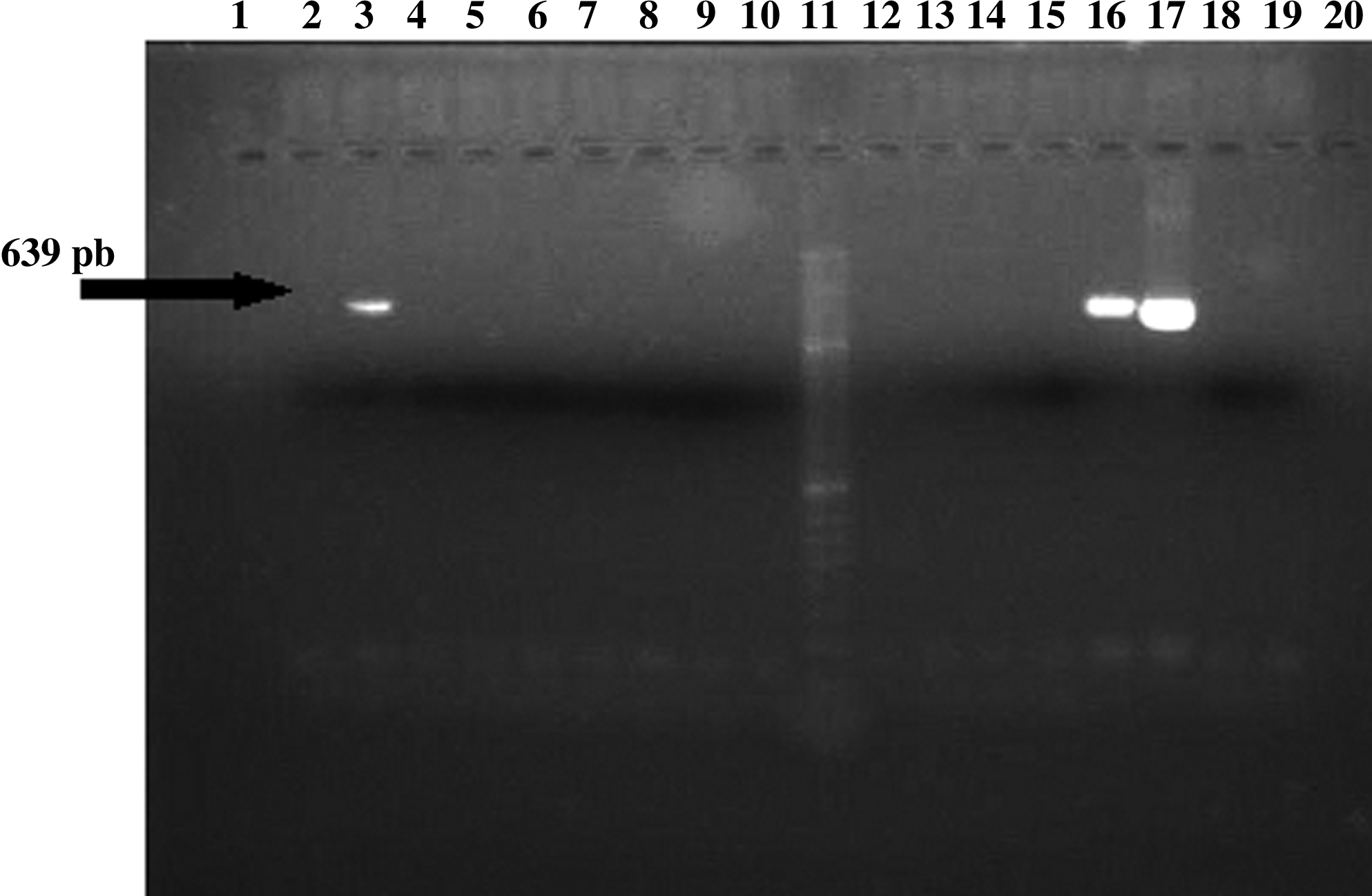
Analysis of erm(B) determinant. PCR products of amplification using primers Erm B-1 and Erm B-2. Lane 3, Corynebacterium urealyticum CB-257; lane 11, DNA ladder; lanes 16 and 17, Streptococcus agalactiae 637 (positive control); lane 18, distilled water (negative control); Other lanes, negative strains. pb, pair of bases.
The presence of the ere(B) gene was evaluated in the 14 strains with the MLSB phenotype but without erm genes, and also in 35 randomly selected strains positive for the erm(X) gene (5 strains of each studied species). However, the presence of the ere(B) gene was not detected in any of the studied Corynebacterium strains.
Discussion
In recent years, macrolide resistance mechanisms have been described in detail in the genus Corynebacterium. 23 In our study, the resistance of the different strains to macrolides was very high (76.38%), a fact that confirms the previously reported high prevalence of resistance among Corynebacterium sp.25,27 Erythromycin showed poor activity against C. urealyticum, C. jeikeium, and C. amycolatum, all of them usually considered as multiresistant species.12,25 Similar results were found in all other studied species. The activity of all other macrolides and clindamycin was almost identical compared to that of erythromycin, and the combination quinupristin–dalfopristin showed better activity than all other studied antibiotics. 15
Among Corynebacterium sp., the most frequently described macrolide resistance mechanism is the presence of methylase enzymes codified by erm(X) class genes. 22 This gene has also been previously detected in C. jeikeium and C. amycolatum.23,32 The results obtained in our study confirm the high prevalence of the erm(X) gene among Corynebacterium species. A high percentage (68.89%) of all strains showed the amplification product of the erm(X) gene, and the correlation between the presence of this amplicon and the MLSB phenotype was 91.93%. Of interest, two strains (one C. striatum and one C. aurimucosum) that were susceptible against all antibiotics and two C. amycolatum strains with the M phenotype carried the amplicon erm(X) gene. It is difficult to find an appropriate explanation for this phenomenon. It has been showed in Staphylococcus strains with an MLSB-inducible resistance phenotype that alterations of the attenuator region would potentially lead to a constitutive expression of the methylase. 7 In an analogous way, deletions or mutations in the codified region could make the erm(X) an inoperative gene.
Fourteen isolates showed an MLSB resistance phenotype despite being negative for all erm genes. This must be due to the presence of other resistance mechanisms. In this sense, mutations in 23S rRNA domain V and alterations of proteins L4 and L22 have originated an MLSB resistance phenotype in Streptococcus pneumoniae.3,8,30 The presence of these mutations in Corynebacterium sp. could be an explanation for the resistance to macrolides, although more studies would be necessary to evaluate this possibility.
The amplification products of the erm(B) gene were detected in two C. urealyticum strains. The presence of this gene has already been described in group A Corynebacterium strains. 17 The prevalence of this gene was very low and highly contrasts with the high percentage of erm(X)-gene-positive strains. Besides, there was a strain that carried the erm(X) and erm(B) genes simultaneously, a characteristic previously reported in Staphylococcus, where the presence of several erm genes is very common. 1
Eight C. amycolatum strains showed the M phenotype. This phenotype is usually mediated by efflux systems that are usually present in Streptococcus and are codified mainly by the mef genes. 6 However, in our study, no amplification products of the mef gene were detected, although this gene has been previously reported in the genus Corynebacterium. 17 Other efflux pump genes such as msr have also been recently reported in this genus, 19 but they were not studied in our report.
The effect of the ere(B) gene, related with hydrolysis of antibiotics, was studied in some strains. The presence of this gene is probably unusual among Corynebacterium sp., because it has been mainly described for Gram-negative bacteria. 22
Conclusions
According to our data erm(X) is the most important gene implicated in the macrolide resistance of all Corynebacterium species analyzed in our study. Other resistance genes like erm(B) must also be implicated in this resistance, although its presence seems to be unusual.
Footnotes
Acknowledgments
The authors acknowledge Instituto de Salud Carlos III for sequence analysis of the strains. This study was funded by grants from FIS (PI030220), and from the CONSOLIDER-INGENIO 2010 Program (FUNCOAT-CSD2008-00023). Nieves Z. Martín-de-Hijas was funded by the Fundación Conchita Rábago de Jiménez Díaz (Madrid, Spain).
Disclosure Statement
No conflicts of interest exist for any authors.