
Abstract
Conjugative IncHI2 plasmids carrying tetracycline, trimethoprim, and sulphonamide resistance genes were recovered from two multiply antibiotic resistant Salmonella enterica serovar Typhimurium isolates from Australian food-producing animals. Transposons related to the mercury resistance transposon Tn1696 were identified in both IncHI2 plasmids. These transposons contained an In4-type class 1 integron that carried a dfrA5 trimethoprim resistance gene cassette and the sul1 sulfonamide resistance gene. These integrons were located in the same position as In4 in Tn1696. The integron from one isolate includes a large transposon-like structure containing four IS26 and the strAB, sul2, blaTEM, and aphA1 genes conferring resistance to streptomycin, sulphonamides, ampicillin, kanamycin, and neomycin, respectively. This structure is flanked by an 8-bp duplication, but it includes both the aphA1-containing transposon Tn4352 and a transposon, Tn6029, carrying genes derived from RSF1010 and from Tn2. However, Tn4352 and Tn6029 overlap, sharing one IS26 copy. This suggests that they do not move by a standard transpositional mechanism. A circular intermediate, carrying only the region containing the resistance gene(s) and one of the IS26 bounding it, is proposed as an intermediate.
Introduction
Tn1696 is also comprised of a class 1 integron carrying antibiotic resistance genes located in a mercury (II) resistance transposon. 10 However, in this case, the mercury resistance transposon is equivalent to Tn5036, 27 which is only 82% identical to the Tn21 backbone, and the integron is in a different position. 10 In4, the class 1 integron in Tn1696, is found within the res site of the backbone transposon (Fig. 1A) and is the exemplar for one of the main types of class 1 integron backbone structures. 14 Tn1696 was found located on an IncP1-α plasmid, R1033, that was isolated in 1975 in Spain, from a clinical Pseudomonas aeruginosa isolate.5,16,19 The complete sequence of Tn1696 can be found in GenBank accession no. U12338.
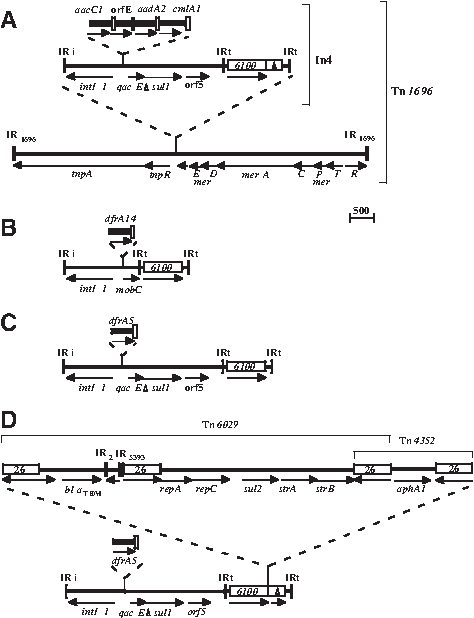
Structure of Tn1696 and integrons in Tn1696 relatives. (
To date, only one further example of a transposon with an integron found in exactly the same location as In4 has been reported. 23 This integron (Fig. 1B) is related to In4 but has different cassettes and has lost the 3′-conserved segment (3′-CS). 14 The transposon is located on an IncHI1 plasmid, pHCM1, from a Salmonella enterica serovar Typhi strain isolated in Vietnam in 1993, 23 suggesting that transposons related to Tn1696 may be more widespread or even globally distributed. However, the transposon in the IncHI1 plasmid carries a copy of the insertion sequence IS4321 in the 38 bp inverted repeat (IR) at the tnp end of the transposon, 11 and this interruption of the transposase binding site should render it unable to move.
Here, we have identified further Tn1696 relatives in S. enterica serovar Typhimurium isolates from Australian cattle. We also found another complex IS26-containing transposon within one of them.
Materials and Methods
Bacterial strains
Two multiply antibiotic resistant S. enterica serovar Typhimurium isolates were studied. They were recovered at the Microbiological Diagnostic Unit (MDU) at Melbourne University from diseased Australian cattle in 1999 and 2000 during routine screening and part of a collection of multiply antibiotic resistant isolates previously described. 7 The serovar, antibiotic resistance profile and phage type were determined at the MDU, as previously described. 7 Isolate SRC26 was phage type PT1 var1 and SRC125 was PT44. The antibiotic resistance phenotypes were confirmed and extended using a disk diffusion assay, the calibrated dichotomous sensitivity (CDS) test with antibiotic disks (Oxoid) containing ampicillin (25 μg), chloramphenicol (30 μg), florfenicol (30 μg), gentamicin (10 μg), kanamycin (50 μg), neomycin (30 μg), streptomycin (25 μg), spectinomycin (25 μg), sulfafurazole (300 μg), tetracycline (30 μg), tobramicin (10 μg), and trimethoprim (5 μg), as previously described. 4 Resistance was defined according to CDS guidelines (http://web.med.unsw.edu.au/cdstest/).
Plasmid isolation and analysis
Polymerase chain reaction (PCR)-based replicon typing was used to identify plasmid types. 2 Plasmids were recovered using conjugal transfer from S. enterica to an Escherichia coli recipient strain, E294 (RifR). 24 One hundred microliters of overnight Luria broth (LB) cultures of donor and recipient were mixed on an LB agar plate and incubated overnight at 26°C or 37°C. Cells were recovered by resuspending in 0.9% (w/v) saline, diluted, and plated onto LB agar containing rifampicin (100 μg/ml) and kanamycin (50 μg/ml) to select for transconjugants and LB agar containing kanamycin to enumerate the donor cells. The transfer frequency was calculated as number of transconjugants per donor. At least 10 transconjugant colonies from each cross were screened for resistance to antibiotics to which the donor strain was resistant, using LB agar containing ampicillin (100 μg/ml), kanamycin (50 μg/ml), neomycin (50 μg/ml), streptomycin (25 μg/ml), or tetracycline (10 μg/ml) and Muller Hinton agar containing sulfonamides (100 μg/ml) or trimethoprim (25 μg/ml). Transconjugants were also screened as described below for the resistance genes found in the donor and for plasmid types.
DNA extraction
Whole cell DNA was made using a boiling lysis method. A single bacterial colony was resuspended in 200 μl of water, incubated at 100°C for 5 min, and centrifuged for 10 min at 20,000 rpm. The resulting supernatant was used as a template for PCR. Plasmid DNA was extracted from 1.5 ml LB cultures of S. enterica or E. coli transconjugants, using an alkaline lysis miniprep method 17 and resuspended at a concentration of ∼1 μg/μl. This DNA was diluted to 10 ng/μl before use as a template for PCR. PCR products used as sequencing templates were purified from gels using the QIAquick Gel Extraction Kit (Qiagen).
Polymerase chain reaction
PCR reactions contained 200 μM of each dNTP, 1 U Taq Polymerase, and 50 pmol of primer in 25 μl of 1 × PCR reaction buffer (supplied with Taq polymerase, New England BioLabs). Reactions were run in an Eppendorf Thermocycler (Eppendorf ) with an initial incubation at 95°C for 5 min, followed by 35 cycles of 95°C for 30 sec, 60°C for 30 sec, 72°C for 1–3 min, and then a final incubation at 72°C for 5 min. DNA fragments were resolved by gel electrophoresis on 1% (w/v) agarose gel, stained with ethidium bromide (5 μg/ml), and visualized using a GelDoc1000 image analysis station (BioRad). Amplicons were identified using standards and identities confirmed either by digestion with restriction enzymes, as per the manufacturer's instructions (New England BioLabs), or by DNA sequencing.
Primers used in this study to screen for antibiotic resistance genes, class 1 integrons, insertion sequences, or merA genes from mercuric ion resistance transposons have been previously described. 15 The merA amplicons were digested with RsaI to distinguish the mer module type. Primers used to identify the location of integrons in class II transposon backbones were, for the IRi end of the integron, a primer in the 5′-CS of class 1 integrons (RH546, 5′-GCT CGG CCA TTC CGA CGT CTC-3′) and a primer (RH620, 5′-GTT CCG GGT TCT GGT CGA AG-3′) that was designed to detect the tnpR gene from a range of class II transposons including Tn21, Tn1696, and Tn1403. 20 Digestion with BamHI, which removes a fragment of 250 bp of the 5′-CS, was used to confirm the identity of the product. For the IRt end, primers in IS6100 (DB-T1, 5′-TGCCACGCTCAATACCGAC-3′) and merE (RH306, 5′-CGCAGCTTGCCGTGTTG-3′) were used. R1033 containing Tn1696, 10 pACYC184::Tn21 3 and R388::Tn1403 20 were used as standards.
DNA sequencing and sequence analysis
Automated sequencing was performed at Macquarie University on an ABI PRISM 377 DNA Sequencer (AME Bioscience) or a 3130 Exel Genetic Analyzer (Applied Biosystems) using the Big Dye system and sequences were assembled using Sequencher version 4.9 (Gene Codes Corporation). All regions of Tn6026 were sequenced in both directions and Tn6025 in one direction. Differences between Tn6026 or Tn6025 and the published Tn1696 sequence (GenBank accession no. U12338) were confirmed using more than one PCR product as sequencing template. Differences found in comparison to Tn1696 were a single base change in urf2Y in both pSRC26 and pSRC125 and a silent mutation in sul1in pSRC26. Sequenced regions were identified using Basic Local Alignment Search Tool (BLAST) or in pair-wise comparisons using the BLAST paired alignment facility (http://blast.ncbi.nlm.nih.gov). Gene Construction Kit version 2.5 (Textco) was used to create figures to scale.
Nucleotide sequence accession numbers
The nucleotide sequences of Tn6025 and Tn6026, including Tn6029, have been submitted to GenBank under accession nos. GU562437 and GQ150541, respectively.
Results
A collection of multiply antibiotic resistant S. enterica serovar Typhimurium isolates from a variety of sources 7 were screened for the presence of various antibiotic resistance genes as well as for class 1 integrons and merA genes. Among them, two isolates, designated SRC26 and SRC125, recovered from cattle in 1999 and 2000 were resistant to sulfonamides, trimethoprim, tetracycline, kanamycin, and neomycin. SRC26 was also resistant to ampicillin and streptomycin. SRC26 and SRC125 both carried a class 1 integron, with the intI1 gene and the sul1 sulfonamide resistance gene present together with the dfrA5 trimethoprim resistance gene cassette. IS6100, which is characteristic of In4-type class 1 integrons, 14 and an merA gene of the type associated with Tn1696 (and Tn5036) were also detected. This combination of features suggested the presence of a transposon related to Tn1696. Both SRC26 and SRC125 also contained the tetA(B) tetracycline resistance gene, an aphA1 gene conferring kanamycin and neomycin resistance, and the strA and strB genes for streptomycin resistance. SRC26 carries further sul2 and blaTEM genes. These genes account for the resistance phenotypes, except that SRC125 did not exhibit streptomycin resistance.
Both isolates also contained an IncHI2 plasmid and IncHI2 plasmids conferring the same resistance profiles, as the Salmonella donor strain was transferred by conjugation into the E. coli strain E294 at 37°C and 26°C at frequencies between 10−5 and 10−6 transconjugants per donor. The sequence of the IncHI2 amplicon was identical to that of the prototype IncHI2 plasmid R478 (GenBank accession no. NC_005211). These IncHI2 plasmids, named pSRC26 and pSRC125, respectively, carried all of the resistance genes found in their parent Salmonella strain.
Integron location and structure of the transposons
The association of the integron with a class II mercury (II) resistance transposon and its position was established by PCR using primers designed to detect the tnpR gene from a range of class II transposons including Tn21, Tn1696, and Tn1403. The size of this product varies with the position of the integron (Fig. 2). In SRC26, SRC125, and the IncHI2 plasmids derived from them, the IRi end of the integron was linked to the tnpR gene as in Tn1696; and sequencing confirmed that it was in precisely the same position. For pSRC125, the IS6100 at the right end of the integron was also linked to a mer module, as it is in Tn1696, using primers in IS6100 and merE; but the partial copy of IS6100 found in In4 (Fig. 1A) was missing (Fig. 1C). However, no product was obtained with pSRC26 (see below). Instead, the IS6100 could be linked to the blaTEM gene.
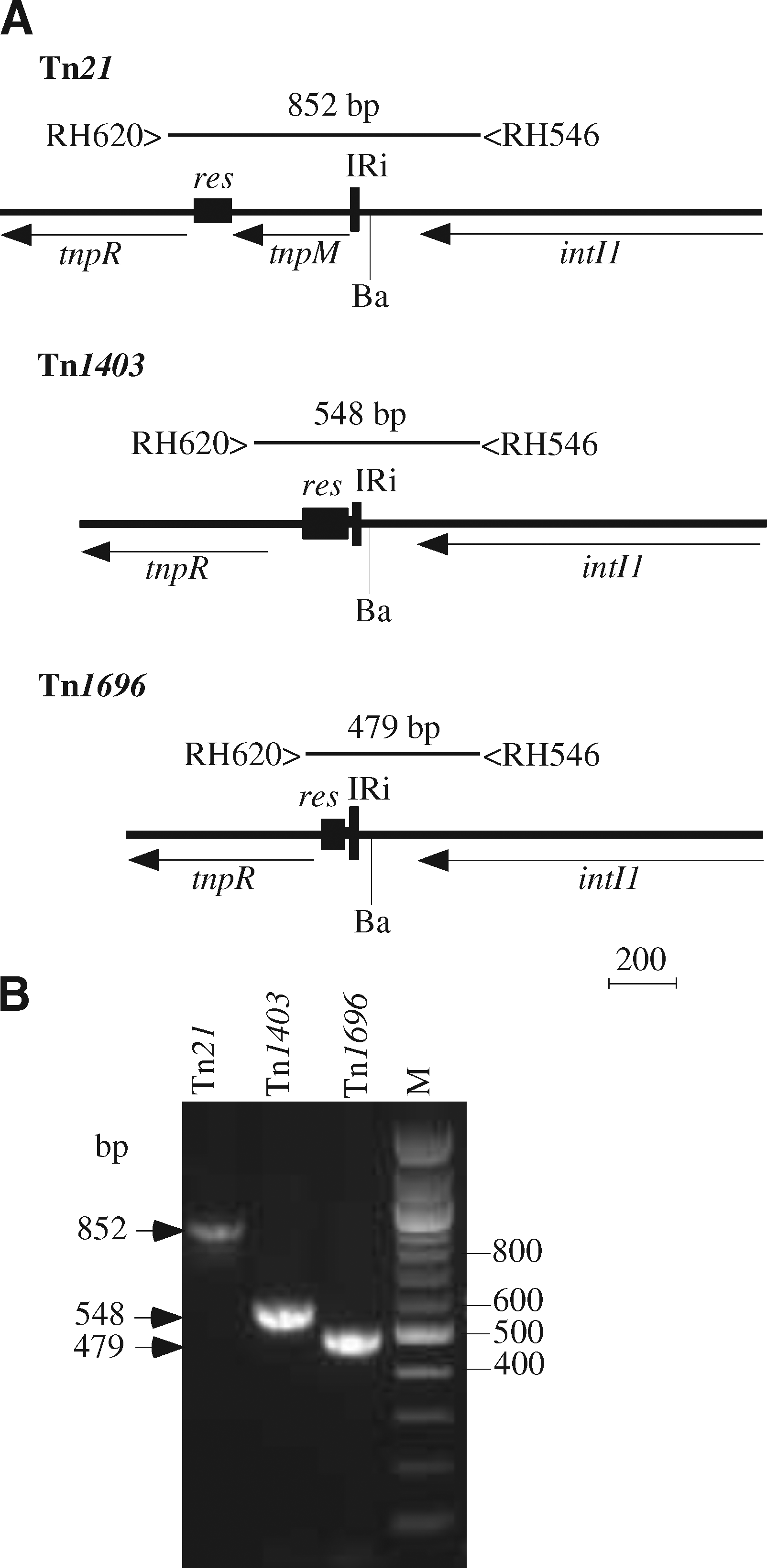
Location of integrons in class II transposon backbones. (
The complete sequence (13,302 bp) of the transposon, designated Tn6025, from pSRC125 was determined using overlapping PCR products as templates, and the backbone mercury resistance transposon is 99.9% identical to that in Tn1696 (GenBank accession no. U12338). The integron, shown in Fig. 1C, carries the strong version of the cassette promoter, Pc, but does not contain the 19-bp duplication found in the 5′-CS of In4, 10 and this difference may reflect an exchange of cassettes by homologous recombination. The sequence of Tn6026 (24,253 bp) from pSRC26 was essentially identical to Tn6025, except that the partial copy of IS1600 was present and a 10,622-bp insertion was located within the complete IS6100 copy (Fig. 1D), which accounted for the failure to link merE to IS6100.
Structure and origin of the insertion in Tn6026
The insertion in Tn6026 (Fig. 1D) was located at one end of the complete copy of IS6100 and consisted of a set of four antibiotic resistance genes and four copies of IS26. It is bounded by directly-oriented copies of IS26 and flanked by a direct duplication of 8 bp, as expected for an IS26 insertion. The insertion includes the aphA1-containing structure known as Tn4352. 25 It also includes a second potential transposon, here designated Tn6029, that consists of three IS26 and the sul2, strAB, and blaTEM-1 genes. However, Tn4352 and Tn6029 overlap, sharing one copy of IS26. The presence of only one IS26 between these two entities indicates that their movement is not via a standard transposition mechanism as used by the class I and class II transposons studied to date. This would bring in both copies of the IS26 bounding the incoming entity (Tn4352 or Tn6029) and would result in the presence of an additional IS26. It would also create duplication flanking that entity.
The segments between the IS26 in Tn6029 are derived from two sources, Tn1, Tn2, or Tn3 13 and plasmid pSRC15 26 (GenBank accession no. GQ379901), which is almost identical to RSF1010. 18 The route by which Tn6029 appears to have been generated is illustrated in Fig. 3. It involves insertion of Tn1, Tn2, or Tn3 into RSF1010 and insertion of three copies of IS26 into the resulting structure followed by the inversion of one of the RSF1010-derived segments.
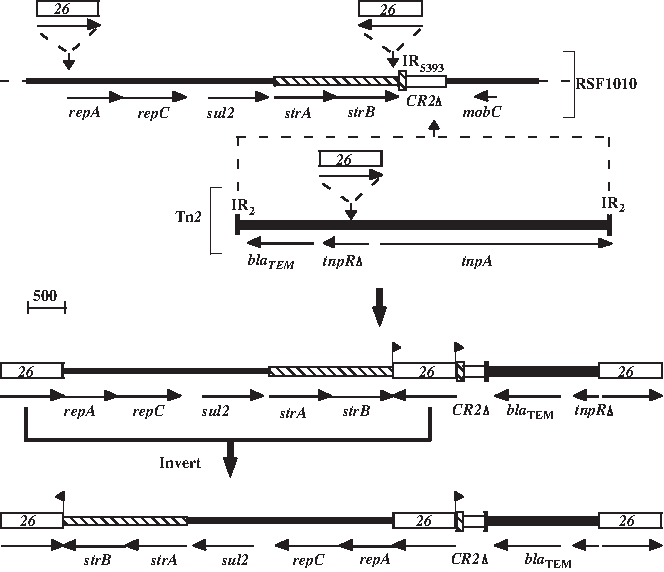
Derivation of the mosaic transposon Tn6029. Horizontal arrows represent the extent and direction of the genes. Lines of different styles represent gene segments with different origins: thin lines, RSF1010; striped open lines, Tn5393; thick lines, Tn2. IS26 are shown as open boxes containing 26. IR represent inverted repeats with the number of the transposon shown as a subscript. The vertical arrows indicate positions of insertion events. The order of these events is unknown. Small flags indicate position and direction of duplications formed from insertion of the central IS26.
A close relative of Tn6029, Tn6029B, is found alone in a Tn21-derived transposon in the IncHI1 plasmid pHCM1 (GenBank accession no. AL513383), 23 and a structure that is the same as the combined Tn6029-Tn4352 entity in Tn6026 is found in precisely the same position in a closely related Tn21-derived transposon in the IncHI1 plasmid, pO111_1, from an enterohaemorragic E. coli strain (GenBank accession no. AP010961). These two structures have potentially been generated in situ by gain or loss of the Tn4352 portion. However, in pHCM1, Tn6029B has suffered an IS26-mediated deletion removing 85 bp of the Tn1-derived portion. Two further Tn6029-Tn4352 copies that share the same location on one side are found in an IncF plasmid, pRSB107, isolated from sewage (GenBank accession no. AJ851089) 21 and an E. coli virulence plasmid, pO26-CRL (GenBank accession no. GQ259888). 22
Discussion
The transposons described here could be directly derived from Tn1696, as they differ from it mainly in features within In4. The cassette array has been exchanged, and the directly orientated partial copy of IS6100 found adjacent to the complete IS6100 in In4 has been lost from Tn6025. Both of these changes can be easily effected by homologous recombination, and the absence of the 19-bp duplication in attI1 that is seen in In4 10 is consistent with this conclusion. Transposons can continue to evolve by the acquisition of further resistance gene carrying transposons, and here a complex structure that carries four additional resistance genes has been added to Tn6026. Recently, another integron-containing relative of Tn1696 has been identified. 9 However, it is likely that Tn6005 is not part of the same lineage as Tn1696, Tn6025, and Tn6026, because it has independently acquired the integron, which is located 6 bp away from In4 in Tn1696 and the integrons in the transposons described here.
We conclude that members of the Tn1696 family are globally disseminated and have invaded various plasmid types. Tn1696 and its relatives should be able to move by forming cointegrates, except if, as in pHCM1, the presence of a copy of IS4321 in one of the IRs of the transposon 11 has immobilized it. However, resolution, the second step of the transposition process, is blocked, because the integron disrupts the res site and resolution presumably occurs via the less efficient homologous recombination. This may account for the multiple cases in GenBank where either the tnp or the mer module of Tn1696 is found alone or associated with another tnp or mer module. Examples are found in pRMH760 12 and the genomic resistance island of Acinetobacter baumannii. 15
Footnotes
Acknowledgments
A.K.C. was supported by a University of Sydney Postgraduate Award, X.L. was supported by an ARC Linkage grant LP0348851 and R.M.H. is supported by NHMRC grant 358713. This work was partly supported by NHMRC project grant 402584 and ARC Linkage grant LP0348851. The S.P.D. laboratory also was partly supported by grants from NSW Department of Primary Industries and the McGarvie Smith Institute. We thank Diane Lightfoot and other staff of the MDU, University of Melbourne, for characterizing and supplying the strains. MDU is supported by The Victorian Department of Human Services.
Disclosure Statement
No competing financial interests exist