
Abstract
Abstract
This study was conducted to identify the β-lactamase content of 30 metallo-β-lactamase-producing Pseudomonas aeruginosa isolated in 2007 from two Tunisian hospitals and to investigate their genetic relatedness. All these isolates produced VIM-2. blaPER-1, blaPSE-1, blaOXA-2, and blaOXA-10 were identified in 17, 5, 21, and 1 isolates, respectively. These enzymes were often associated in the same isolate: 26 isolates had at least two β-lactamases. The predominant serotype was O12. Pulsed-field gel electrphoresis revealed genetic diversity among the metallo-β-lactamase-producing P. aeruginosa isolates. This is the first report on the existence of blaPER-1, blaPSE-1, blaOXA-2, and blaOXA-10 in Tunisia.
Introduction
Five types of acquired metallo-β-lactamase (MBL) have been identified: IMP, VIM, SPM, GIM, and SIM.31,37 They have been detected primarily in P. aeruginosa.31,37 The first member of the VIM family determinants, VIM-1, was identified from a clinical isolate of P. aeruginosa in Verona, Italy, in 1997. 13 Subsequently, blaVIM positive P. aeruginosa strains have been reported from many countries of the world.38,39 However, the most common MBL identified worldwide is now VIM-2, which was identified from a P. aeruginosa strain in France in 1996.27,37,39,41
In contrast to the enterobacterial species in which TEM- and SHV-type enzymes are most frequent, OXA- and PSE-types are the most frequently acquired β-lactamases in P. aeruginosa.14,19 Other types of class A ESBL (PER, VEB, GES, and IBC) have spread in this species in specific geographical locations19,21,40: PER-1 β-lactamase is most widespread in Turkey, VEB-1, in Southeast Asia; and GES-like β-lactamases have been identified in French, Greek, and South African isolates.19,28,29 Very few data are available regarding the prevalence of the different acquired β-lactamases among P. aeruginosa in Tunisia. Only OXA-18, SHV-2a ESBLs, and VIM-2 MBL were described in this specie in three Tunisian hospitals.8,11,17,18
The aim of this study was to investigate the diversity and frequency of β-lactamases in 30 MBL-producing P. aeruginosa isolated in two Tunisian hospitals in 2007 and to ascertain their clonal relationship.
Materials and Methods
Bacterial strains
In 2007, 30 nonrepetitive of carbapenem resistant P. aeruginosa isolates were obtained from clinical specimens of 11 different wards of two hospitals in Sfax-Tunisia, the majority of which were isolated from patients hospitalized in the intensive care unit (15 isolates, 50%). All the strains were identified with the API 20NE system (BioMérieux). The screening EDTA-test gave positive results to all isolates.
P. aeruginosa ATCC 27853 and Escherichia coli ATCC 25922 were used as controls during susceptibility testing.
A multidrug resistant Klebsiella pneumoniae isolate that harbored two conjugative plasmids carrying the blaVIM-4, blaCMY-2, blaCTX-M-15, and blaTEM-1 genes was used as positive control for resistance transfer determination. 12
Serotyping
The clinical isolates of P. aeruginosa were serotyped by slide agglutination with O-specific antisera (Bio-Rad).
Antibiotic susceptibility and screening for ESBLs
Antibiotic susceptibility was determined by the disk diffusion method on Mueller-Hinton (MH) agar plates with β-lactam and non β-lactam antibitic-containing disks (Bio-Rad). Minimal inhibitory concentrations of β-lactams (ticarcilline, ceftazidime, cefepime, aztreonam, and imipenem) were determined by the standard agar dilution method on MH agar. Antimicrobial susceptibility results were interpreted according to the criteria of the French Society for Microbiology. 2 (www.sfm.asso.fr).
Detection of ESBL was carried out by a double-disk synergy method testing ceftazidime, aztreonam, and cefepime at a distance of 15 mm from ticarcillin clavulanic acid disks on MH agar plates.
Polymerase chain reaction amplification of β-lactamase genes and sequence analysis
Total DNA of P. aeruginosa isolates was extracted by boiling. We investigated the presence of β-lactamases by polymerase chain reaction (PCR) using the previously published specific primers for blaTEM, blaSHV, blaPSE, blaCARB-4, blaCTX-M-1, blaCTX-M-2, blaCTX-M-9, blaPER, blaGES, blaVEB, blaVIM, blaIMP, blaOXA-1, blaOXA-2, blaOXA-10, blaOXA-18, blaOXA-5, blaOXA-20, and blaLCR-1.1,3 Cycling parameters for all tested genes were one cycle of 95°C for 5 min followed by 35 cycles of 95°C for 30 sec, 55°C or 60°C for 40 sec, and 72°C for 1 min. The PCR products were sequenced on ABI Prism 3100 automated sequencer (Applied Biosystems) and were analyzed using NCBI BLAST program. (www.ncbi.nlm.nih.gov/).
Plasmid content analysis and mating-out assays
Plasmid DNA extraction was carried out for the clinical isolates of P. aeruginosa and the positive control strain using two different extraction methods.10,33
Conjugation experiments were performed with E. coli J53-2 rifR as a recipient strain. Transconjugants selection was performed on MH agar plates containing aztreonam (4 μg/ml), or cefotaxime (4 μg/ml), or ceftazidime (10 μg/ml), or imipenem (2 μg/ml) and rifampin (250 μg/ml).
For transformation experiments by electroporation, E. coli DH10B and E. coli Top10 cells were used according to the manufacturer's instructions (Bio-Rad). Transformants were selected on MH agar plates containing cefotaxime or ceftazidime or imipenem.
Pulsed-field gel electrphoresis typing
Pulsed-field gel electrphoresis (PFGE) was carried out with a GenePath system (Bio-Rad). Genomic DNA was digested with SpeI, and the fragments were separated in agarose gels by electrophoresis according to the manufacturer's recommendations. Image normalization and construction of similarity matrices were carried out using the Fingerprinting II software (Bio-Rad).
Results
Bacterial strains, antimicrobial susceptibility, transfer of resistance, and molecular characterization of β-lactamase genes
By disk diffusion method, all isolates of P. aeruginosa were resistant to ticarcilline, ticarcillin-clavulanic acid, ceftazidime, cefotaxime, cefepime, and imipenem. Twenty-one isolates were susceptible to aztreonam. The minimal inhibitory concentration determinations showed high levels of resistance to all β-lactams tested (Table 1). The percentages of resistance to gentamicin, amikacin, ciprofloxacine, and fosfomycine were 97.5%, 40%, 57.5%, and 22.5%, respectively.
ICU, Intensive care unit; ER, Emergency room; NS, Neurosurgery; M, Internal Medicine; BU, Burn unit; O, Orthopedics; S, Surgery; H, Hematology; N, Neurology; MF, Maxillofacial; PFGE, pulsed-field gel electrphoresis; IMP, imipenem; CAZ, ceftazidime; ATM, aztreonam; FEP, cefepime; TIC, ticarcillin.
PCR amplification and sequencing of the amplified fragments confirmed the presence of blaVIM-2 gene in all isolates (Table 1). The blaPER-1 was identified in 17 isolates; only five of them exhibited an ESBL phenotype. OXA-2 and PSE-1 were detected in 21 and 5 of the isolates, respectively. The blaOXA-10 was recognized in one isolate. TEM, SHV, CTX-M, CMY, IMP, VEB, CAR-4, GES, BES, LCR, IMP, and OXA-1 enzymes were not detected.
The serotypes of all isolates are listed in Table 1. The majority of P. aeruginosa MBL-producing isolates (16 isolates, 53%) belonged to serotype O12. The serotypes O4, O5, O7, O11, and O16 were observed for thirteen other strains. Only one isolate was polyagglutinable (Table 1).
Repetitive attempts to transfer β-lactam resistance to E. coli recipient by conjugation and by electroporation assays remained unsuccessful.
Epidemiological analysis by PFGE
PFGE showed that P. aeruginosa isolates producing MBL were genetically unrelated in our hospital (Fig. 1). Twenty-three genotypically different strains were identified (Table 1), suggesting that P. aeruginosa isolates have originated from different ancestors. However, six serotype O12 isolates were clonally related, as they revealed a similar pulsotype (A) (shared ≥90% genetic similarity by the Dice coefficient). These six isolates produced VIM-2, OXA-2, and PER1 β-lactamases. Two strains isolated in the same ward and producing VIM-2 associated with OXA-2 or PSE-1 belonged to genotype B. One genotype (clone C) included two isolates with two different subclones C1 and C2. The remaining 20 isolates were each distributed in a single pulsotype (Fig. 1).
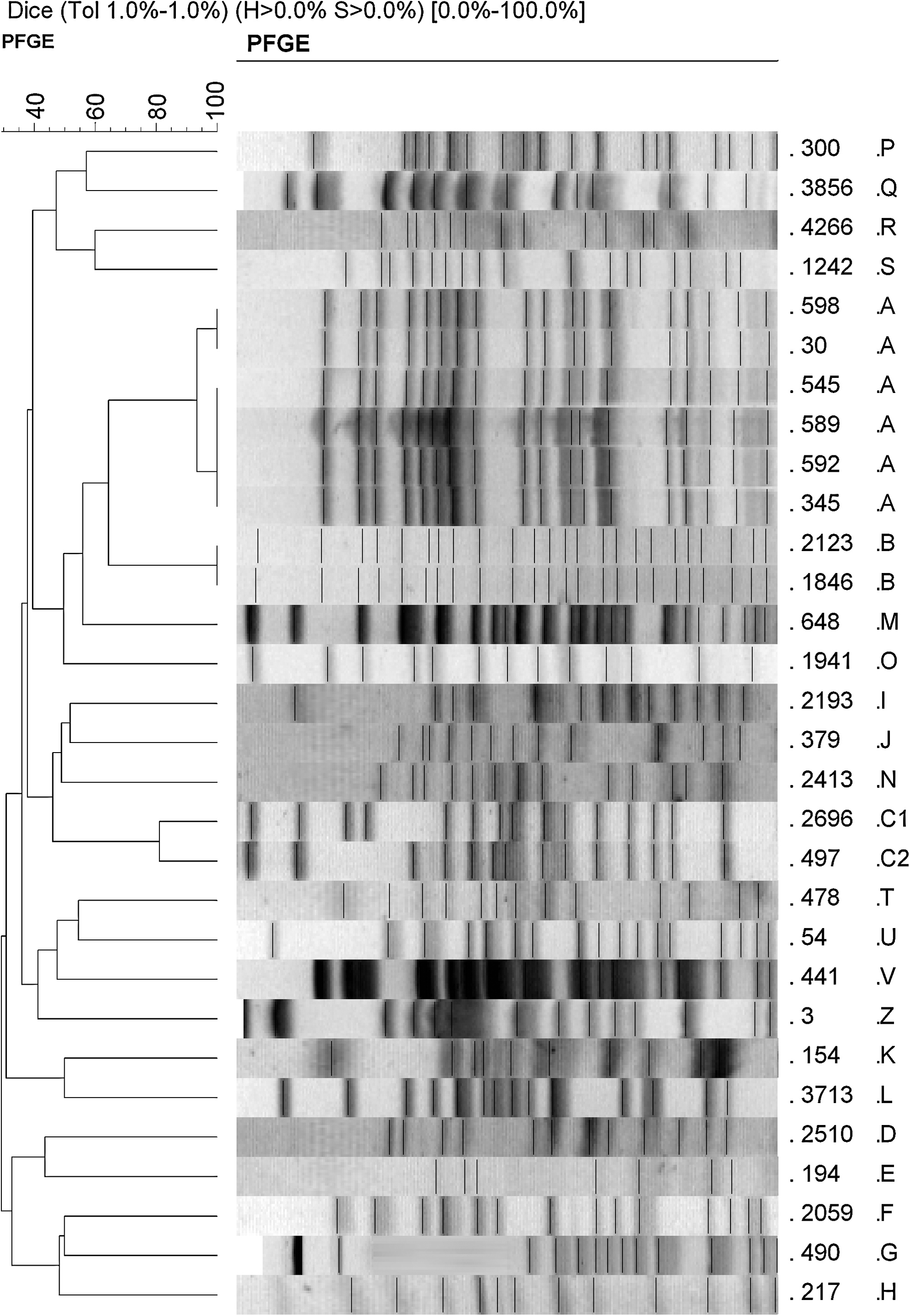
Dendrogram generated by PFGE (PFGE analysis of genomic DNA fragments digested with SpeI). PFGE, pulsed-field gel electrphoresis.
Discussion
In our study, we have investigated 30 multidrug resistant isolates of P. aeruginosa producing MBL isolated in 2007. The majority of these strains belonged to serotype O12. In fact, it has been repeatedly demonstrated over the past 20 years that serotype O12 dominates among multiresistant P. aeruginosa isolates, especially MBL producers. 16 Moreover, it was suggested that serotype O12 strains are particularly adept at acquiring or hosting the integrons that encode the MBL enzymes and are responsible for the dissemination of blaVIM genes. 6 The 30 investigated strains were producing a VIM-2 MBL. Among these isolates, 17, 21, and 5 harbored PER-1, OXA-2, and PSE-1, respectively. These enzymes were often associated in the same strain: 26 isolates (90%) had at least two β-lactamases, among which 4 isolates harbored VIM-2 and OXA-2, 3 harbored VIM-2 and PSE-1, 1 harbored VIM-2 and PER1, and 1 harbored VIM-2 and OXA-10. VIM-2 was associated with PER-1 and OXA-2 in 15 isolates. Two strains harbored four β-lactamases: VIM-2, PER-1, OXA-2, and PSE-1.
Five important transferable MBL families, VIM-, IMP-, GIM-, SPM-, and SIM, were identified in P. aeruginosa. The most common and widespread acquired MBL is now VIM-2, which exhibits a worldwide distribution.37–39,41 The VIM-2 enzyme has been described in several countries, namely, France, Italy, Greece, Turkey, Japan, South Korea, Portugal, Spain, Poland, Croatia, Chile, Venezuela, Argentina, Belgium, and the United States. 39
In Tunisia, acquired MBLs had initially emerged among an epidemic K. pneumoniae clone in our hospital (Sfax-Tunisia) in 2005. 12 This clone produced VIM-4 metallo-enzyme associated with CTX-M-15 ESBL and CMY-4 AmpC enzyme. The VIM-2 enzyme had been previously found in P. aeruginosa in the University Hospital Sahloul, Sousse, and in Charles Nicolle Hospital, Tunis, Tunisia.8,18
Unlike MBL, which had spread among P. aeruginosa, ESBLs remain rare in this species. The ESBLs reported for P. aeruginosa are SHV, TEM, PER, VEB, BEL, GES, and, more recently, CTX-M types.17,19,25,34 There are few reports describing the isolation of TEM- or SHV-type ESBLs in P. aeruginosa. SHV-2a was reported in France and Tunisia, SHV-5 in Greece, and SHV-12 in Thailand and Japan.17,34 The OXA and PER types remain the most common ESBLs found in this species. P. aeruginosa carrying these ESBLs had been shown to have a specific geographical distrubition, for example, PER-1 and OXA-10 are widespread in Turkey and VEB-1 is widespread in Kuwait.19,28 A few ESBL-producing P. aeruginosa isolates were documented in Africa, particularly in North Africa. In fact, the present study is the first report of the blaPER-1 in P. aeruginosa isolates in Tunisia. In 1991, PER-1 first identified from P. aeruginosa were recovered in France from a Turkish patient.21,22 Since 1995, PER-1-producing organisms have been disseminating in Turkey, Italy, and, more recently, in other countries. 5 The prevalence of PER-1 gene reached 34.61% in Italy, 11% in Turkey, and 49.25% in Tehran, Iran.19,36 In Tunisia only, the ESBL similar to OXA-18 and SHV-2a have been detected in P. aeruginosa.8,11,17 Two outbreaks of P. aeruginosa strains carrying SHV-2a have been reported in two university hospitals in Sousse and Tunis-Tunisia.8,17 Another nosocomial outbreak of OXA-18-producing P. aeruginosa was reported in the national Bone Marrow Transplant Centre of Tunisia. In the latter study, TEM-1 was identified in 2 isolates and SHV-1 in 21 isolates; but PSE, CARB type, and PER were not detected. 11
The present study is the first report of co-expressing VIM-2 and PER-1 in the same isolates of P. aeruginosa in Tunisia. The coproduction of VIM-2 and PER-1 in P. aeruginosa was already detected in Turkey, Italy, and, recently, in France.4,9,41
Further, in our study, we reported the association of PER-1 with OXA-2. The blaOXA-2 gene has been often identified in P. aeruginosa isolates producing the ESBL PER-1. 5 Recently, blaOXA-2 has been identified in P. aeruginosa isolates from Serbia and Hungary that also produced the ESBL PER-1. 15
In addition in our study, VIM-2 was coexpressed with PSE-1. PSE and CARB-type enzymes are the most common β-lactamases in P. aeruginosa. 11 In Bulgaria, PSE-1 β-lactamases were detected in 22.5% of the investigated strains recovered during the period 2001–2006 in five university hospitals. 32 In Korea, of the 252 isolates collected between April and June 2002, 16 (6.3%) harbored PSE-type enzyme and 53 (21.0%) isolates harbored OXA-type enzyme. 14 In the comparative distribution of β-lactamases during two study periods 1984–1989 (128 isolates) and 1994–1999 (131 isolates) at Beaujon Hospital, PSE-1 was predominant (162 isolates, 62.5%) over both periods, followed by enzymes of OXA groupI. 1 In the present study, OXA-10 associated with VIM-2 was described in only one isolate. The prevalence of OXA-10 varies from 13% in Korea, 17.33% in Turkey, to 74.62% in Iran.14,19,35
Repetitive attempts to transfer blaVIM-2 and genes of ESBLs by electroporation and conjugation remained unsuccessful. These results suggest that these genes might be chromosomally located in P. aeruginosa isolates responsible for a polyclonal outbreak. A recent study conducted in the Sahloul Hospital, Sousse, Tunisia, showed that the blaVIM-2 gene was very likely chromosomally located. 18 In Poland, all MBL genes (blaVIM-2 and blaVIM-4) identified were chromosomally encoded. 24 In Turkey, the two β-lactamase genes blaVIM-2 and blaPER-1 were chromosomally located. 36 In addition to the MBL genes, the genes of extended β-lactamase in clinical isolates of P. aeruginosa were found to be chromosomally located, namely, blaSHV-2a, blaCTX-M-2, and blaPER-1.5,17,25 However, other studies showed that blaVIM-2 gene could be transferred horizontally by plasmids.7,20,23,27
As previously reported, in this study PFGE showed that the majority of the investigated P. aeruginosa isolates producing VIM-2 were genetically unrelated.20,26,30 In contrast, VIM-2-producing P. aeruginosa have been shown to be involved in two hospital outbreaks in Tunisia.8,18 So, it is noteworthy that the original fear that a clone of VIM-2-producing P. aeruginosa would become established in our hospital has not been realized. However, we have found that six O12 strains carrying PER-1, OXA-2, and VIM-2 were clonally related. These strains could spread later in our hospital. In fact, the clonal dissemination of P. aeruginosa strains with PER-1 in hospitals in Korea, Turkey, Italy, and Belgium has been previously reported. The concurrent presence of two or more unrelated clones of P. aeruginosa with the PER-1 enzyme in some hospitals has also been previously described. 5
In conclusion, the present study is the first to describe the association of blaPER-1, blaPSE-1, blaOXA-2, and blaOXA-10 among genetically unrelated P. aeruginosa producing VIM-2 in a Tunisian hospital. The emergence of the simultaneous production of four β-lactamases by P. aeruginosa clinical isolates, identified for the first time in Tunisia, is alarming and it poses challenges for the treatment of hospital infections caused due to this species. A continued surveillance of this production of β-lactamases should be adopted in all isolates of P. aeruginosa to limit their further dissemination.
Footnotes
Acknowledgments
This study was financially supported by the Ministry of Scientific Research Technology and Competence Development of Tunisia.
Disclosure Statement
No competing financial interests exist.