
Abstract
In this study, the fitness cost of fluoroquinolone resistance was evaluated in vitro, on food matrices, and in vivo, using Campylobacter coli and Campylobacter jejuni in vitro selected mutants. In vitro, the growth rate of the susceptible (wild type) and resistant (mutant) strains did not differ when cultured separately. However, by conducting sequential passages of mixed cultures, the ratio of the resistant mutant to the susceptible strain decreased for C. coli but not for C. jejuni. When the wild type and the mutant were co-inoculated on food matrices, mutants were no longer detectable 3 to 5 days after artificial contamination, but the wild-type strains remained detectable for over 13 days. In mono-inoculated animals, no difference was observed between wild-type and mutant fecal titers. When co-inoculated into chickens, the susceptible strain outcompeted the resistant mutant for C. coli and for C. jejuni. However, for C. coli, if the resistant strain was already present in animals, it could persist at high titers in the digestive tract even in the presence of the wild-type strain. Together, these findings suggest that, depending on strain and study conditions, fluoroquinolone resistance can impose a fitness cost on Campylobacter.
Introduction
The main targets of FQs in bacteria are topoisomerase IV and/or DNA gyrase. 11 A single point mutation in the quinolone resistance-determining region (QRDR) of gyrA can lead to relevant levels of resistance. Although there are many positions and types of mutation in gyrA such as Thr86Ala, Asp90Asn, Asp90His, Pro104Ser, and Ala70Thr, the majority of FQ resistant isolates, whether they are C. jejuni or C. coli, have a mutation in gyrA that results in the Thr86Ile substitution.1,16
Resistance acquired by chromosomal mutation has been shown to impose fitness costs both in vitro21,24,27 and in vivo,7,8,14 even though in some cases, no-cost mutations or enhanced fitness have been observed. 19 Fitness cost is defined as reduced growth and/or persistence of resistant bacteria in the host or environment, or increased clearance from infected host and reduced transmission between hosts compared with susceptible bacteria. It is typically measured via laboratory experiments in culture media or animal models.3,5,20 Reduction in fitness is expressed as reduced growth, virulence, or transmission. 4 Nevertheless, some resistant isolates develop compensatory mutations that reduce fitness cost associated with antimicrobial resistance. These compensatory mutations depend on many factors such as the environment (in vivo or in vitro), the antibiotic agent, or its target in the cell.
This study aimed at determining whether the acquisition of FQ resistance, through the most frequent mutation (Thr86Ile), affects the fitness of C. coli and C. jejuni on three different conditions: in vitro, on food matrices, and in vivo in the chicken digestive tract. Chicken skin fragments were chosen as food matrices, as food of chicken origin is considered as a main source of human contamination. Chicken digestive tract is known to be the reservoir of Campylobacter.
Materials and Methods
Bacterial strains and growth conditions
Two strains C. coli 103 and C. jejuni 126 were used as susceptible strains for selection of ciprofloxacin resistant mutants. Two ciprofloxacin resistant strains C. coli 157 and C. jejuni 109 were used as DNA donors for transformation. All strains had been previously isolated from healthy chicken caeca at slaughterhouse, by culture on Karmali agar plates supplemented with selective agents (Oxoid, Dardilly, France), and identified to the species level by multiplex polymerase chain reaction (PCR). 10 Campylobacter strains were grown routinely on Mueller-Hinton agar (MHA) plates or in Mueller-Hinton broth (MHB) (Biorad, Marnes la Coquette, France) at 42°C, under microaerophilic conditions (O2: 5%; CO2: 10%; N2: 95%). When needed, the culture medium was supplemented with ciprofloxacin (Sigma, St Quentin Fallavier, France) at a final concentration of 4 mg/L.
Construction and characterization of genetically defined mutants
PCR was used to amplify a 306 bp fragment of the QRDR of the gyrA sequence from the resistant strain C. coli 157 and a 254 bp fragment of the QRDR of the gyrA sequence from the resistant strain C. jejuni 109, as described next. DNA was purified using the Mini-elute gel extraction kit (QIAgène, Courtaboeuf, France) following the manufacturer's instructions. The purified PCR products prepared from C. coli 157 and C. jejuni 109 strains were used to transform C. coli 103 and C. jejuni 126 strains, respectively, by electroporation using 1 μg of DNA (PCR products). Transformants were selected on MHA plates supplemented with ciprofloxacin (4 mg/L). Multiple transformants obtained from each transformation were sequenced to verify the presence of the unique C257T mutation in the QRDR of the gyrA sequence and to verify the absence of any mutation in the gyrB sequence. Minimal inhibitory concentration (MIC) determination was used to confirm ciprofloxacin resistance character.
PCR amplification and sequencing of QRDRs of the gyrA and gyrB genes
Primers used to amplify the QRDR of the gyrA and gyrB sequences in C. coli and C. jejuni sequences are listed in Table 1. A reaction mixture containing the primers at 0.5 μM each, deoxynucleoside triphosphates at 200 μM each, 1 × reaction buffer (50 mM KCl, 10 mM Tris-HCl), MgCl2 at 1.5 mM, 1 U of Extra pol II DNA polymerase (Eurobio, les Ulis, France), and 5 μl of genomic DNA template was used for PCR amplification. After an initial denaturation step of 5 min at 94°C, thirty-five amplification cycles were performed. Each one consisted of 30 sec of denaturation at 94°C, 30 sec at a hybridization temperature of 53°C, and 40 sec of extension at 72°C. A final extension step of 10 min was performed at 72°C. DNA was purified, and sequence reaction in a final volume of 20 μl was performed using the same primers listed in Table 1 at 0.16 μM, 4.5 μl of the Big Dye Ready Reaction Mix (Applied Biosystems, Marnes la Coquette, France), and 1 μl of purified PCR products. The sequence reaction conditions began with a denaturation step at 96°C for 1 min followed by 25 cycles at 95°C for 10 sec, at 55°C for 5 sec, and at 60°C for 4 min. After DNA precipitation, the obtained fragments were separated in a 3130 Genetic Analyzer capillary sequencer (Applied Biosystems). Nucleotide sequences were aligned using the site: http://bioinfo.genotoul.fr/multalin/. C. coli strain sequences were aligned using C. coli NCTC 11353. C. jejuni strain sequences were aligned using C. jejuni NCTC 11168.
Antimicrobial susceptibility testing
MICs of ciprofloxacin, enrofloxacin, nalidixic acid, ampicillin, erythromycin, and tetracycline (Sigma) were determined for all strains using the agar dilution method (CLSI, M31-A3). According to the recommendations of EUCAST (www.eucast.org/), strains with MIC(Cip) > 1 mg/L were considered resistant to FQs.
Frequency of spontaneous FQ resistance
C. coli 103 and C. jejuni 126 cultures were grown in antibiotic-free MHB to the logarithmic phase. The cultures were plated in triplicate on antibiotic-free MHA plates and ciprofloxacin-containing plates. The frequency of FQ resistance was calculated as a ratio of the numbers of colony forming units (CFUs) on ciprofloxacin-containing plates to the CFUs on the ciprofloxacin-free plates.
Growth kinetics and in vitro competitive assay
Growth curves of the susceptible strains and ciprofloxacin mutants were obtained separately and by competition of wild type (wt) and mutant of the same species, with three repeats conducted for each experiment. A 24-hr culture of each strain was inoculated in 15 ml of MHB to obtain a final optical density (OD) of 0.015 at 600 nm. For growth competition, equal volumes of parent and mutant cultures were added together in 15 ml of MHB to also obtain a final OD600 of 0.015. All cultures (wt, mutant, and wt + mutant) were incubated at 42°C with shaking (140 rpm) under microaerophilic conditions for 48 hrs. About 0.9-ml samples from each culture were collected at 0, 3, 6, 9, 24, and 48 h. The OD was measured, and the number of viable cells in each sample was estimated by spreading in triplicate a 100 μl aliquot to the appropriate dilutions onto MHA plates and MHA plates supplemented with ciprofloxacin 4 mg/L. After incubation, the number of viable cells in the suspension was estimated in terms of CFUs. The number of viable ciprofloxacin-susceptible cells in growth competition was calculated by subtracting the number of colonies on MHA plates with ciprofloxacin from the number of colonies on MHA plates without ciprofloxacin.
To assess small differences during long-term bacterial growth, serial passages were performed: 100 μl of a culture in log phase was removed at 24 h and added to 15 ml of fresh MHB medium. The cultures were incubated under the same conditions for 24 h and underwent a new transfer of 100 μl of the culture to 15 ml of fresh medium. Eight passages were successively performed. The CFUs of the resistant and susceptible strains were evaluated at each passage as previously described. Results were expressed by the ratio of ciprofloxacin-resistant (FQR) to susceptible (FQS) colonies (ratio CFU/ml [FQR]/CFU/ml [FQS]).
Comparison of survival on food matrices
Skin fragments of specific pathogen free (SPF), Campylobacter spp. free chickens, were used as examples of food matrices. 5 cm × 5 cm fragments from the skin of recently euthanized birds were placed individually in Petri dishes and inoculated with cultures of the susceptible strain, the mutant or both (susceptible and mutant) of each species. For each fragment, 5 ml of an inoculum suspension adjusted to 105 CFU/ml was used. Fragments were incubated at 22°C ± 2°C or 5°C ± 3°C under normal oxygenation conditions. Campylobacter titers in each well plate were analyzed on several days after artificial contamination, by vigorous agitation of the skin fragment for 60 seconds in 45 ml of MHB, by spreading in triplicate a 100 μl aliquot to the appropriate dilutions onto MHA plates and MHA plates supplemented with ciprofloxacin. This experiment was repeated twice.
In vivo experiments
Two independent experiments were conducted with 10-day-old SPF Campylobacter spp. free chickens. The experimental design is detailed in Table 3 and was decided on to enable comparison of the colonization, persistence, and diffusion capacities of the isolates and their mutants, when inoculated separately or in competition. Moreover, when the birds were inoculated with both the susceptible strain and its resistant mutant, either the inoculations were performed on the same day, or the resistant mutant was inoculated four days before or four days after the isogenic wild type isolate, to study competition between susceptible and resistant isolates or to evaluate whether the presence of the resistant isolate in the digestive tract could prevent colonization by the isogenic susceptible one and inversely. In experiment I, chickens were inoculated with C. coli (susceptible and/or resistant strains); whereas in experiment II, chickens were inoculated with C. jejuni (susceptible and/or resistant strains). Twenty birds were housed in each isolator, and five isolators were used in each experiment. To compare the diffusion capacity of the susceptible and the resistant strains, only 10 chickens per isolator were infected as indicated in Table 2, whereas the other ten birds were not inoculated. Thus, in each isolator, 10 birds were inoculated, via oral gavage, with 100 μl of an inoculum suspension adjusted to 0.5 McFarland (∼108 CFU/ml). The bacterial inoculums of the initial suspensions were quantified by plating 100 μl 1:10 serial dilutions on MHA plates. After inoculation, fecal samples were collected from five randomly selected inoculated chickens and five randomly selected contact animals (noninoculated chickens) from each isolator at different time points. Samples were serially diluted to one-tenth in MHB, and then 100 μl were plated in triplicate, onto MHA with Campylobacter selective agents to detect total Campylobacter colonies, and onto MHA with Campylobacter selective agents and ciprofloxacin to detect Campylobacter colonies resistant to ciprofloxacin. MICs and gyrA and gyrB genes sequences of a few clones obtained on the last sampling day were determined as previously described. Assays were performed according to the French animal welfare regulations (Authorization B-22-741-1, granted by the Departmental Veterinary Services Authority).
MICs determined by the agar dilution method.
MIC, minimal inhibitory concentration; CIP, ciprofloxacin; NAL, nalidixic acid; ENR, enrofloxacin; ERY, erythromycin; AMP, ampicillin; TET, tetracycline.
CFU, colony forming units.
Statistical analysis
In the animal experiments, the detection limit of the plating method was considered to be 100 CFU/g of feces. Comparison of the geometric mean values between the titers of the wild type and mutant strains was performed using Student's t-test or the Mann and Whitney test depending on the normality of distributions and homogeneity of variances. Differences between the mean values were considered significant at p < 0.05.
Results
Characteristics of FQR Campylobacter wild-type strains and mutants
For wild-type strains C. coli 103 and C. jejuni 126, the MICs of ciprofloxacin were, respectively, 0.25 mg/L and 0.06 mg/L. MICs of other antibiotics are represented in Table 2. After sequencing of the QRDRs of the gyrA and gyrB genes, no mutations either in gyrA or in gyrB were detected in either wild-type strains. For resistant strains C. coli 157 and C. jejuni 109, the MICs of ciprofloxacin were 8 mg/L. MICs of other antibiotics are represented in Table 2. After sequencing of the QRDRs of the gyrA and gyrB genes, only the C257T mutation in the gyrA gene corresponding to the Thr86Ile substitution in the GyrA subunit was detected. No mutation was detected in the QRDR of the gyrB gene.
The specific C257T mutations in gyrA genes were amplified from C. coli 157 and C. jejuni 109 FQR strains and introduced by transformation into the wild-type strains C. coli 103 and C. jejuni 126, respectively. Characterization of C. coli 103T86I and C. jejuni 126T86I transformants was performed by measuring the MIC of ciprofloxacin, enrofloxacin, and nalidixic acid and by sequencing the QRDR of their gyrA and gyrB genes. The MICs of ciprofloxacin were 4 mg/L for both isogenic mutants C. coli 103T86I and C. jejuni 126T86I. The MICs for the other antibiotic families of mutants were unchanged (Table 2). Both transformants had the C257T mutation in the gyrA gene corresponding to Thr86Ile substitution in the GyrA subunit; no other mutation was detected in their gyrA and gyrB QRDR.
Effect of FQ resistance on the in vitro fitness of C. coli and C. jejuni
The in vitro study was performed using mono-inoculated cultures or competition tests. Thus, to determine whether the number of colonies counted on ciprofloxacin-supplemented MHA plates during the competitive assay corresponded to the FQR population obtained from the inoculated mutant only, or was a mixture of inoculated mutants and spontaneous resistant mutants arising from the inoculated wild-type strains, the frequency of spontaneous FQ resistance acquisition was calculated for strains C. coli 103 and C. jejuni 126. Spontaneous FQ-resistant mutants were obtained with a mean frequency of 6 × 10−8 for C. coli 103 and 2.5 × 10−7 for C. jejuni 126. Thus, spontaneous resistance mutation frequency was considered negligible compared with the number of inoculated resistant mutants.
Inversely, to determine whether the number of FQS colonies, calculated by subtracting the number of colonies on MHA plates with ciprofloxacin from the number of colonies on MHA plates without ciprofloxacin, corresponded to the inoculated wild-type strains or to a mixture of inoculated wild-type cells and revertants from inoculated 103T86I and 126T86I mutants, spreading of cultures was always performed in parallel on MHA plates and MHA plates supplemented with ciprofloxacin. The number of colonies on the two types of plates was not significantly different for the two mutants (data not shown). Thus, reversion of resistant strains to susceptibility during the competitive assay was considered negligible, and the number of colonies calculated by subtraction did actually correspond to the FQS inoculated population.
Kinetics curves of susceptible and isogenic mutants separately or in competition for 48 hrs did not show any significant difference of the titers at 3, 6, 9, 24, and 48 hrs (data not shown). To assess slight differences in bacterial growth during the long term, eight sequential passages were conducted. When grown separately, again no differences were observed between the daily titers of FQS and FQR strains either in C. coli or in C. jejuni (data not shown). Results for growth competition passages are represented in Fig. 1A as a ratio of CFU/ml (FQR)/CFU/ml (FQS) according to 8 passages. For C. coli, this ratio decreased with sequential passages, thus indicating that the FQS strain 103 became dominant in the medium with regard to the FQR strain; after 5 passages, the FQR C. coli strain 103T86I could no longer be detected. For C. jejuni, when strains were in competition (Fig. 1A) no difference in the CFU/ml (FQR)/CFU/ml (FQS) ratio was detected during the 8 passages; the value of this ratio remained approximately constant during the experiment.
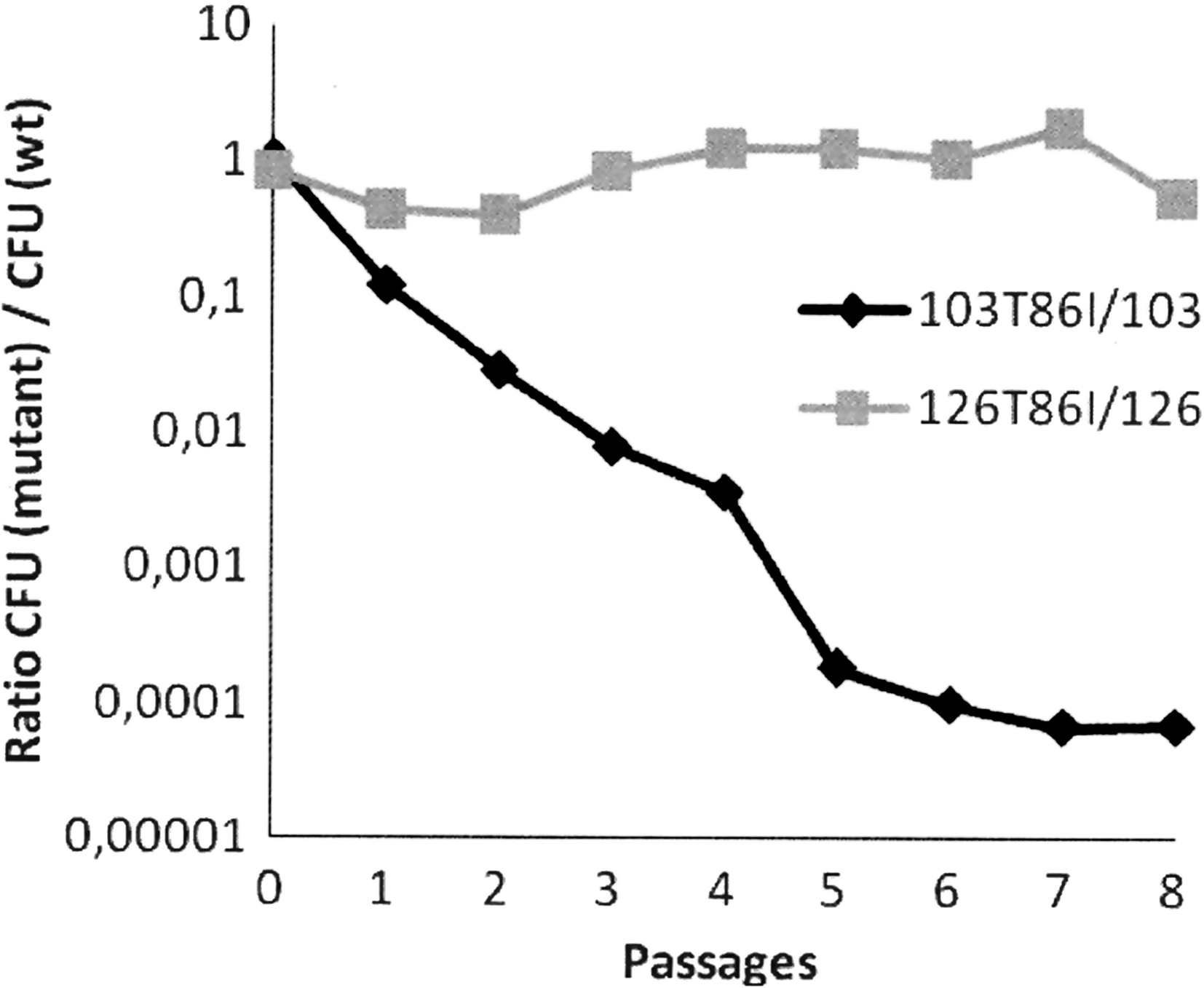
In vitro population dynamics of the susceptible strain (wt) and its resistant mutant in mixed cultures. Results were expressed as a ratio of CFU (mutant)/CFU (wt) at the respective passage points. Light grey line: CFU (126T86I)/CFU (126) and dark grey line: CFU (103T86I)/CFU (103).
Effect of FQ resistance on the survival of C. coli and C. jejuni on food matrices
To assess the fitness impact of FQ resistance on food matrices, skin fragments were inoculated with FQS and/or FQR strains separately or in competition, either for C. coli or for C. jejuni. At 5°C ± 3°C, no significant difference between the titers of FQS strains and their respective FQR isogenic mutants was detected (data not shown). At 22°C ± 2°C, no significant difference was noted between the duration of detection of the strains and their respective mutants on the skin fragments when they were mono-inoculated (12 days for C. coli 103 and 103T86I; 12 days for C. jejuni 126 and 126T86I), but a difference was observed when C. coli or C. jejuni FQS strains and their respective FQR mutants were co-inoculated. As shown in Fig. 2A, C. coli 103 was still detected after 13 days on food matrices, whereas the corresponding mutant 103T86I was no longer detected after only 7 days. Similar results were observed for C. jejuni 126, which remained detectable for more than 13 days on chicken skin whereas the mutant C. jejuni 126T86I was no longer detected after only 5 days (Fig. 2B).
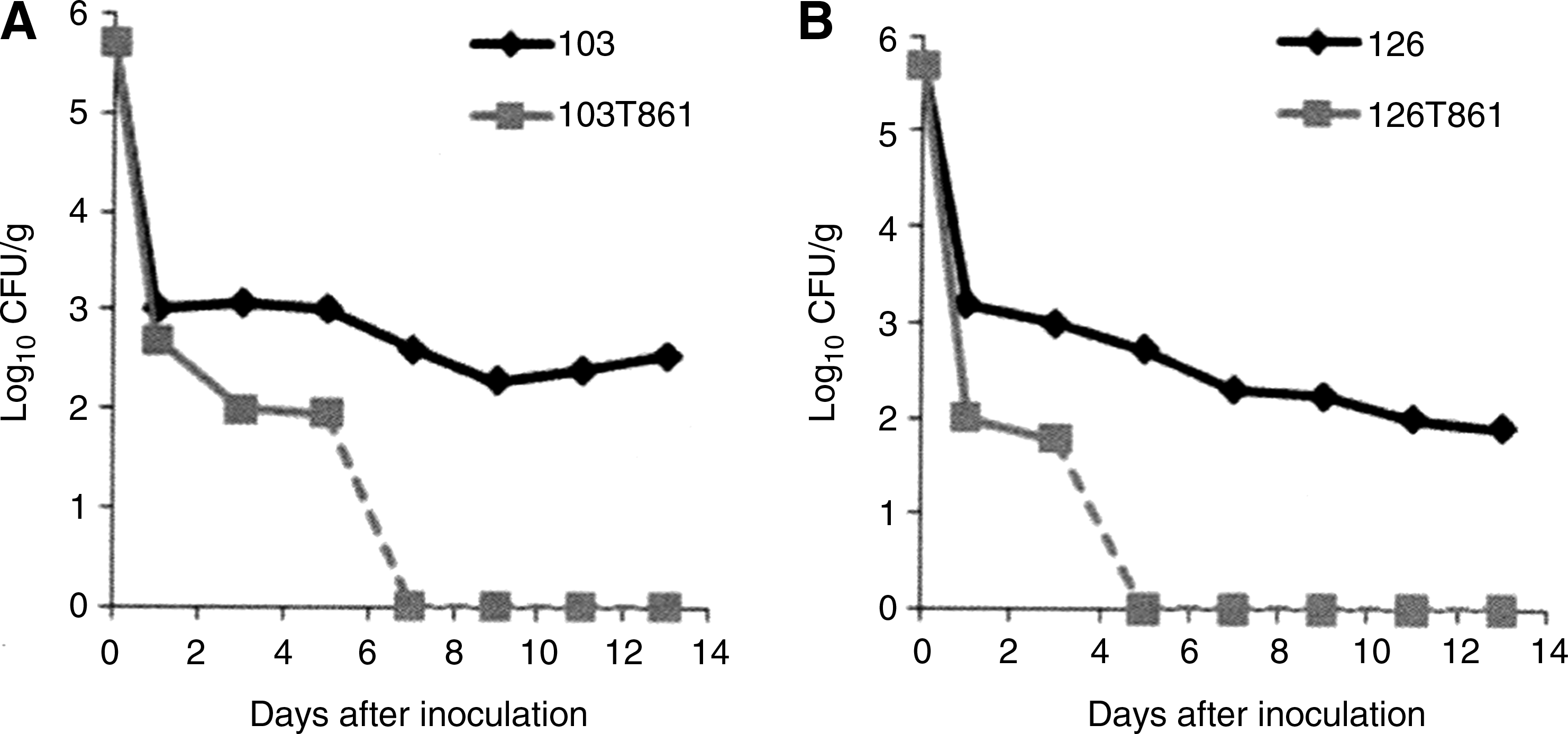
Effect of FQ resistance on the in vivo fitness of C. coli
To determine whether the acquisition of FQ resistance influences the in vivo fitness of C. coli, SPF chickens were infected with either 103 or its mutant 103T86I, or both strains 103 and 103T86I in competition. The FQS or FQR C. coli mono-inoculated chickens were colonized within 4 days (first sampling day). The individual titers varied greatly among chickens, between 105 and 108 CFU/g of feces. Thus, as shown in Fig. 3A no significant difference was observed in the mean level of FQS and FQR strains when mono-inoculated in animals. However, as shown in Fig. 3B, a significant difference in the mean level of colonization between 103 and 103T86I (p < 0.001) was observed when the strains were simultaneously co-inoculated into chickens. Mean titers of 103 were always higher than those of 103T86I: thus, as early as day 7 postinoculation, for C. coli 103 the mean titer was 2.35 ± 1.08 × 105 CFU/g of feces whereas the mean titer for C. coli 103T86I was 5.4 ± 1.5 × 103 CFU/g of feces (p < 0.001). At day 11, 17, 23, and 30 postinoculation, for C. coli 103 the mean titers were, respectively, 7.56 ± 7.4 × 106, 3.29 ± 1.58 × 105, 1.02 ± 0.7 × 107, and 1.13 ± 1.08 × 107 CFU/g of feces whereas the mean titers for the resistant C. coli 103T86I were, respectively, 3.7 ± 1.7 × 103, 1.48 ± 1.4 × 103, 102, and 2.8 ± 1.85 × 102 CFU/g of feces. To further examine the fitness of FQR C. coli, chickens in two different isolators were inoculated with 103T86I 4 days after inoculation with 103 or inversely. Figure 3C shows the mean level of CFU/g of feces of FQS and FQR C. coli strains when the susceptible strain was introduced before the resistant one. A significant difference (p < 0.001) was observed between titers of 103 and 103T86I in the feces collected 7 to 30 days postinoculation with higher titers for the susceptible strain 103 (approximately 2 logs difference after 30 days of inoculation). Inversely, when the resistant strain 103T86I was introduced into chickens before the susceptible 103, no significant difference was observed between titers of the two strains in the feces (Fig. 3D), indicating that when the FQR strain was already present in animals, it could persist at high titers in the digestive tract even in the presence of the wild-type strain.
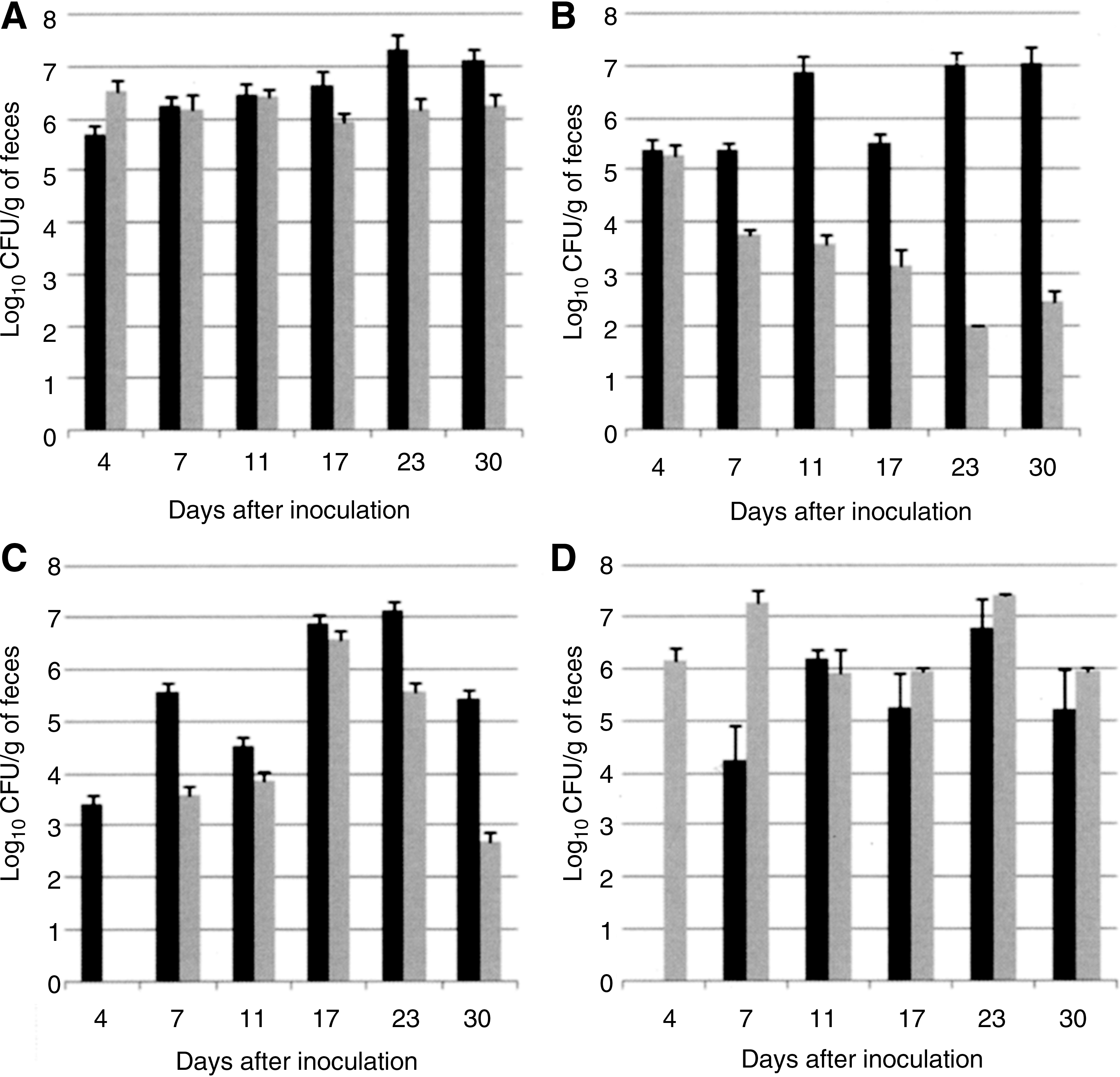
Experiment I: Colonization of FQS and FQR C. coli in the chicken host.
To compare the spread of FQS and FQR strains, titers of 103 and 103T86I were also analyzed in contact in the four previously described conditions. Four days after animal inoculation, the mean titer in the feces of animals in contact with chickens mono-inoculated with 103 was 1.48 ± 1.6 × 106 CFU/g of feces, whereas the mean titer for 103T86I contact chickens was 4.43 ± 0.85 × 103 CFU/g of feces (p = 0.001). However, when animals were placed in contact with co-inoculated birds, samples collected 4 days after inoculation had approximately equal titers of susceptible and resistant strains. The mean titer for C. coli 103 was 1.53 ± 1.8 × 105 CFU/g of feces, and the mean titer for C. coli 103T86I was 1.18 ± 1.5 × 105 CFU/g of feces (p > 0.05). Similarly to mono-inoculated animals, birds placed in contact with chickens inoculated with 103T86I 4 days after inoculation with the susceptible strain 103 had on their first sampling day a mean titer of 6.65 ± 18.3 × 105 CFU/g of feces for 103 and a mean titer of 8.6 ± 11.5 × 102 CFU/g of feces for 103T86I (p = 0.01). The mean titer in the feces of animals in contact with chickens inoculated with 103 4 days before inoculation with 103T86I was 3.25 ± 5.06 × 104 CFU/g of feces for 103, whereas the mean titer for 103T86I in contact chickens was 2.26 ± 3.02 × 103 CFU/g of feces (p > 0.05).
Effect of FQ resistance on the in vivo fitness of C. jejuni
To determine whether FQ resistance influences the in vivo fitness of C. jejuni, SPF chickens were infected with either FQS C. jejuni 126 or FQR C. jejuni 126T86I, or with a mixture of both strains at the same titers. Chickens were colonized by both strains on day 11 postinoculation and shed the organism at levels between 102 and 105 CFU/g of feces.
Titers were compared for the samples collected during all sampling days and date by date. When mono-inoculated, no significant difference was observed between mean titers of 126 and 126T86I regardless of the sampling day. As shown in Fig. 4A, both strains were shed by the hosts at approximately equal CFU levels (between 104 and 107 CFU/g of feces 17 days postinoculation).
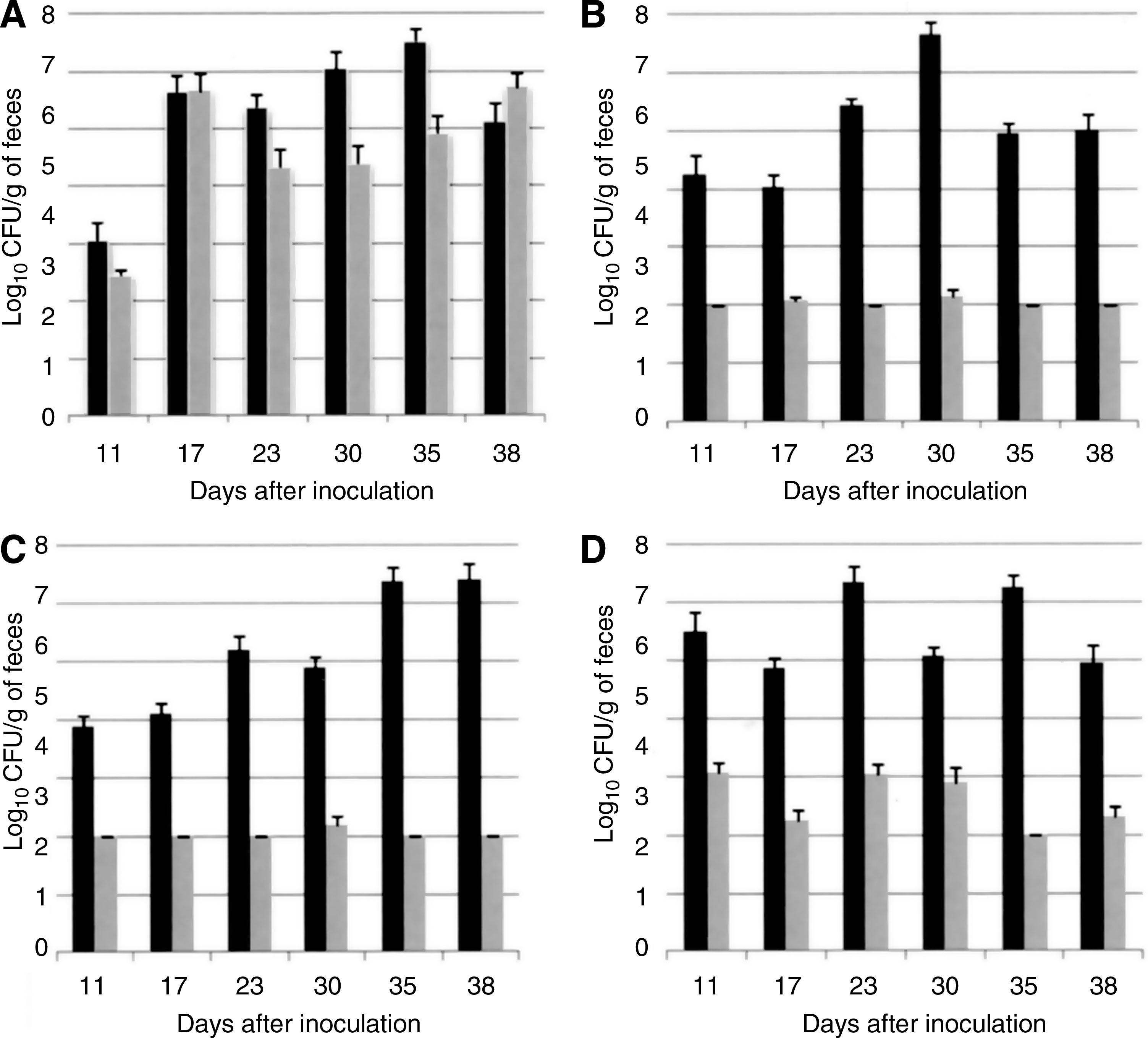
Experiment II: Colonization of FQS and FQR C. jejuni in the chicken host.
When co-inoculated into the host, the susceptible strain 126 colonized animals more efficiently than 126T86I. Significant differences in the level of colonization between 126 and 126T86I (p < 0.001) were observed on all sampling days when strains were inoculated in competition in chickens. Resistant Campylobacter could not be detected by direct plating of fecal samples collected 11, 23, 35, and 38 days after inoculation, and, according to the detection limit, was considered to be less than or equal to 102 CFU/g of feces for 126T86I whereas those of 126 varied between 104 and 107 CFU/g of feces (Fig. 4B).
To further examine the fitness of FQR C. jejuni, chickens in two different isolators were inoculated with 126T86I four days after inoculation with 126 or inversely. The mean levels of CFU/g of feces for FQS and FQR C. jejuni are shown in Fig. 4C for 126T86I introduced in chickens 4 days after 126, and in Fig. 4D for 126 introduced in chickens 4 days after 126T86I. Regardless of the order of inoculation, the two groups of animals yielded the same results, that is, the dominance of 126 over 126T86I (p < 0.001). Mean titers of 126T86I remained between 102 and 103 CFU/g of feces from the beginning until the end of the experiment, whereas mean titers of 126 were greater than 104 CFU/g of feces after 17 days and reached 106 CFU/g of feces 35 days after inoculation.
To compare the spread of 126 and 126T86I strains to noninoculated animals, SPF chickens were placed in contact with the inoculated chickens in the four conditions previously described. After 11 days of contact between inoculated and noninoculated animals, titers of 126 and 126T86I were analyzed in fecal samples collected from noninoculated animals. When placed in contact with mono-inoculated chickens, animals had titers of 1.04 ± 1.8 × 104 CFU/g of feces for 126 and titers of 1.2 ± 0.4 × 102 CFU/g of feces for 126T86I (p = 0.02). When placed in contact with co-inoculated chickens, animals had titers of 1.04 ± 2.1 × 104 CFU/g of feces for 126 and titers of 102 CFU/g of feces for 126T86I (p > 0.05). When placed in contact with chickens inoculated with 126T86I, 4 days after inoculation with 126, animals had titers of 5.9 ± 8.8 × 103 CFU/g of feces for 126 and titers of 102 CFU/g of feces for 126T86I (p = 0.01), whereas when 126 was inoculated four days after inoculation with 126T86I, the mean titers of contact birds were 6.6 ± 1.8 × 103 CFU/g of feces for 126 and 2.2 ± 2.46 × 102 CFU/g of feces for 126T86I (p > 0.05). These significant differences between the levels of titers of FQS and FQR C. jejuni indicate that the spread of the C. jejuni susceptible strain took place more rapidly than that of the C. jejuni resistant strain regardless of the conditions of colonization in animals.
Stability of FQ resistant Campylobacter during in vivo experiments
When mono-inoculated into the host, the susceptible and the resistant C. coli and C. jejuni displayed similar levels of colonization. To determine whether the absence of fitness cost in this case was due to compensatory mutations, sequencing of QRDR gyrA and gyrB was performed for 10 clones of C. coli and C. jejuni collected from different mono-inoculated animals at the end of the experiments. Amino acid sequences for the QRDR of GyrA and GyrB between inoculated strains and isolates obtained at the end of the experiment were identical as well as the MICs of ciprofloxacin, enrofloxacin, and nalidixic acid.
These findings suggested that FQ resistant Campylobacter colonized chickens for a prolonged period without losing gyrA mutation, modification of FQs MICs, or acquisition of compensatory mutations in QRDR of gyrA or gyrB genes.
Discussion
To better understand the impact of FQ resistance on C. coli and C. jejuni fitness, isogenic mutants were constructed and studied, in vitro, on food matrices, and in vivo. Mutants were prepared in vitro by transformation of susceptible strains. Thus, for strains of both species, the acquired mutation was the Thr86Ile mutation, which is more commonly observed in field or clinical strains.1,16 The mutation introduced in wild-type strains resulted in increased FQ MICs, but susceptibility to other compounds was maintained.
In vitro analysis was first performed with separate cultures in optimal conditions for Campylobacter, that is, rich medium, optimal temperature, and atmosphere conditions. In these conditions, the separated cultures of wild-type strains and mutants did not exhibit significant differences. Other examples of no fitness cost due to antibiotic resistance have already been described.9,13,23 However, during competition tests, a distinct competitive growth advantage of the FQS strain over its resistant mutant was observed for C. coli 103. This was not observed for C. jejuni 126, as the mutant had a titer similar to the wild-type strain even after eight daily passages in competition cultures. It remains to be determined whether the C257T mutation conferring FQ resistance is truly cost-free in vitro for C. jejuni or whether the cost it confers is so low that it cannot be measured with current laboratory methods. 4
Differences between wild-type strains and resistant mutants were sought on food matrices. Chicken skin was chosen as the model, as the presence of skin in chicken products is a determining factor in the overall prevalence of Campylobacter. 25 In our conditions, susceptible and resistant strains exhibited similar levels of survival when inoculated separately, whereas the acquisition of FQ resistance in C. coli and in C. jejuni resulted in a rapid disappearance of FQR strains during competition tests. GyrA is a key enzyme for DNA supercoiling. Since GyrA structure is affected in the mutant, accumulation of positive supercoils in front of the transcription complex could take place. Prakash et al. showed that a large number of stress inducible genes depend on the extent of the genomic DNA supercoiling in Cyanobacterium synechosystis. 22 It is possible that the transcription and the expression of multiple genes affect the viability of the resistant strain on food matrices. Thus, in the case of C. jejuni, FQ resistance could result in a survival disadvantage, which would be beneficial to the consumer. The mechanisms of reduced survival of the resistant strain in the presence of the susceptible one could be due to interactions between strains, leading to decreased viability or an increased sensitivity to competing agents or storage conditions of the mutant, or competition for nutrients. El-Shibiny et al. 12 studied the viability of different Campylobacter strains applied as axenic or mixed inoculums on chicken skin during storage at 4°C. They noticed that only one of the three antibiotic resistant strains tested had enhanced survival capacities compared with antibiotic sensitive strains and reported that the presence of another strain could result in a significant reduction in the rate of viability loss. Their experiment showed that the FQ resistant C. coli strain had an increased survival rate when it was paired with another C. coli strain. However, since the resistant and susceptible strains were not isogenic, it is not possible to attribute the observed differences to the resistance characteristics.
The chicken colonization, persistence, and diffusion capacities of Campylobacter are essential characteristics for this zoonotic bacteria. Mono-inoculations in SPF chickens did not reveal any differences, and mutants appeared to colonize the chicken digestive tract as efficiently as the wild-type strains. However, the spread of the FQS C. coli or C. jejuni to contact birds was apparently more rapid than the spread of the FQR mutant. Moreover, for C. jejuni and C. coli pairs, when co-inoculated in chickens, the susceptible strains outcompeted the resistant ones. Thus, for both species, FQ resistance induced a fitness cost, which was not observed in mono-inoculated birds but was revealed during competitive colonization. The more efficient colonization by the susceptible strains may be due to various mechanisms such as reduced susceptibility to aggressive conditions in the stomach during inoculation, a more rapid multiplication in the digestive tract, reduced susceptibility to the competing flora, or to bacteriocins in the digestive tract. The more rapid spread of the wild-type strains from mono-inoculated birds to contact birds could be explained by a better survival of the wild-type strain in the environment. Thus, FQS strains were better colonizers and spread more rapidly to contact chickens than their respective mutants. However, previous results from our group, using another C. jejuni strain and its mutant bearing the Thr86Ile mutation in gyrA inoculated separately or together in SPF chickens, showed no biological cost for the FQ resistance. 17 In vivo fitness of FQ-resistant C. jejuni was also previously studied by Luo et al., 19 who interestingly showed that a single mutation in the gyrA gene (C257T) enhanced the fitness of the resistant strain in a chicken-infection model, but their work demonstrated that when transferred into a different genetic variant of C. jejuni, the same mutation imposed a fitness cost on two mutants prepared from the same parent strain. Our results confirm the latter observation and again suggest that depending on the genetic background of the recipient strain, the C257T mutation can either enhance or reduce fitness of Campylobacter in the chicken host.
Finally, our results clearly indicate that the acquisition of FQ resistance, through C257T mutation in the gyrA gene, imposed a fitness cost in C. coli and C. jejuni. Two common points for C. coli and for C. jejuni can be deduced from this study: first, when cultured separately, or mono-inoculated into animals, FQS and FQR strains displayed the same level of colonization and survival on food matrices. Second, when cultured in competition, or co-inoculated into animals, FQS and FQR strains exhibited different levels of colonization and survival on food matrices. Thus, an interesting finding of this study was the diverse effect of C257T mutation in gyrA on Campylobacter fitness. FQ resistance may or may not impose a fitness cost depending on the conditions of the study, the environment, and the bacterial species. How the gyrA mutation affects Campylobacter fitness is still unknown. Since DNA gyrase is important for DNA replication, the C257T mutation in the gyrA gene may affect the supercoiling activity of the mutant enzyme and the gene expression in Campylobacter.18,28 Understanding precisely how resistance-conferring mutations affect the adaptation of Campylobacter will facilitate the development of means to predict and prevent transmission of this antibiotic-resistant foodborne pathogen.
Footnotes
Acknowledgments
We are grateful to Odile Balan and all the members of the SEEPA service for their help with the animal experiments and to Corinne Marois for her helpful reading of the manuscript. Funding: Salman Zeitouni was financed by the French Ministry of Research. This work was supported by the French agency for food, environmental and occupational health safety.
Disclosure Statement
No competing financial interests exist.