
Abstract
Abstract
The in vitro activity of colistin was evaluated against 215 nonduplicated Pseudomonas aeruginosa isolates, including 53 multidrug-resistant isolates, which were collected between 2006 and 2007 from nine tertiary care hospitals in Korea. Colistin-nonsusceptible P. aeruginosa (CNPA) isolates were genotyped using multilocus sequence typing. Sixteen (7.4%) CNPA isolates (minimum inhibitory concentration [MIC], >2 mg/l) were identified, including three resistant isolates. All but one of the MDR P. aeruginosa isolates was susceptible to colistin. Multilocus sequence typing analysis identified 12 sequence types (STs) among 16 CNPA isolates, indicating that colistin nonsusceptibility might arise independently. However, ST244 and ST292, which may be international clones, were found in multiple CNPA isolates. Our data indicate an increase of P. aeruginosa isolates with reduced colistin susceptibility, suggesting the need for continuous surveillance of P. aeruginosa.
Introduction
In Korea, colistin resistance has been reported for Acinetobacter baumannii and Klebsiella pneumoniae.9,22 Further, it has been demonstrated that colistin resistance in A. baumannii and K. pneumoniae was acquired independently.21,22 Independent and repeated acquisition of colistin resistance may be related to the increasing use and/or inadequate use of colistin. However, colistin resistance in P. aeruginosa isolates from Korea and their clonality have not been investigated. In this article, we report the repeated and independent emergence of colistin-nonsusceptible P. aeruginosa isolates in Korea.
Materials And Methods
P. aeruginosa isolates
A total of 215 P. aeruginosa clinical isolates from patients with bacteremia (101 isolates) or UTIs (114 isolates) were collected between November 2006 and August 2007 from nine Korean tertiary-care hospitals: Samsung Medical Center (SMC, Seoul), Kangbuk Samsung Hospital (KSH, Seoul), Kyungpook National University Hospital (KNUH, Daegu), Daegu Fatima Hospital (DFH, Daegu), Chonnam National University Hospital (CNUH-G, Gwangju), Chungnam National University Hospital (CNUH-D, Daejeon), Chungbuk National University Hospital (CNUH-C, Cheongju), Gyeongsang National University Hospital (GNUH, Jinju), and Jeju National University Hospital (JNUH, Jeju). UTI was diagnosed according to the definition put forth by the Centers for Disease Control and Prevention (CDC). A bacterial isolate with a density ≥105 cfu/ml in urinary culture or ≥108 cfu/ml in suprapubic puncture was identified as a causative pathogen of UTI. Only the first isolate from each patient was included in this study. Species identification of isolates was performed by VITEK-2 system and were confirmed by a PCR method. 2
Antimicrobial susceptibility testing
In vitro antimicrobial susceptibility testing was performed by a broth microdilution method according to the Clinical and Laboratory Standards Institute (CLSI) guidelines. 4 MICs were determined by triplicate experiments for each isolate in cation-adjusted Mueller-Hinton broth and were incubated at 37°C for 20 hours. Fourteen antimicrobial agents were tested: colistin sulfate, polymyxin B, imipenem, meropenem, piperacillin/tazobactam, ampicillin/sulbactam, cefepime, ceftriaxone, ceftazidime, cefoperazone/sulbactam, tetracycline, ciprofloxacin, amikacin, and rifampin. Susceptibility was defined according to CLSI breakpoints. Colistin-nonsusceptible P. aeruginosa (CNPA) isolates were selected on the basis of intermediate susceptibility (MIC, 4 mg/l) or resistance (MIC, ≥8 mg/l) to colistin. 4 Escherichia coli ATCC 25922 and P. aeruginosa ATCC 27853 were employed as reference strains. An MDR isolate was defined as an isolate that demonstrated resistance to three or more different classes of the four following classes: (1) aminoglycoside (amikacin); (2) fluoroquinolones (ciprofloxacin); (3) cefepime, ceftazidime, or piperacillin/tazobactam (considered together as one class); and (4) carbapenems (imipenem or meropenem). Polymyxins were not used in the classification of MDR isolates.
Population analysis profiles
Colistin heteroresistance was also analyzed by population analysis profiles. Population analysis of CNPA isolates was performed as described previously 13 with slight modification. Briefly, 50 μL of bacterial cell suspension (∼109 cfu/ml) and/or its serial dilutions was plated on Muller-Hinton agar plates containing 0, 0.5, 1, 2, 4, 6, 8, and 10 mg/l of colistin sulfate. Colonies were counted after 48 hours of incubation at 37°C. Two colistin-susceptible P. aeruginosa (CSPA) isolates were used as controls. The limit of detection was 20 cfu/ml (i.e., 1.3 log10 cfu/ml), and the lower limit of quantitation (LOQ) was 400 cfu/ml (i.e., 2.6 log10 cfu/ml).
Multilocus sequence typing
Multilocus sequence typing (MLST) was performed as described previously. 5 PCR fragments of seven housekeeping genes, namely, acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE, were obtained from chromosomal DNA and directly sequenced. Allelic profiles and sequence types (STs) were assigned at the Web site (www.pubmlst.org/paeruginosa). New alleles and STs were submitted to the MLST Web site and approved.
Statistical analysis
Pearson's Chi-square test was used to compare categorical variables. All data were analyzed using the SPSS statistical software package (version 11.5; SPSS). Differences were considered statistically significant at a p-value of <0.05 for all tests.
Results
Bacterial isolates and susceptibilities
Of the 215 P. aeruginosa isolates, 3 (1.4%) and 13 isolates (6.0%) were estimated to be resistant (MIC, 8 mg/l) and to have an intermediate susceptibility (MIC, 4 mg/l) to colistin, respectively (Table 1). Thus, a total of 16 isolates (7.4%) were defined as nonsusceptible to colistin. Seven CNPA isolates were from blood, and nine were from patients with UTIs. Thus, 6.9% and 7.9% of isolates from bacteremia and UTIs were nonsusceptible to colistin, respectively. Sixteen of the CNPA isolates were isolated from only three hospitals: SMC (10 isolates), CNUH-G (5 isolates), and JNUH (one isolate). No polymyxin B-nonsusceptible P. aeruginosa isolates were found. However, most of colistin-nonsusceptible isolates displayed rather high MIC values of polymyxin B, 2 mg/l. In addition, the MIC90 value of both colistin and polymyxin B was 2 mg/l. Imipenem and meropenem resistance rates were 23.7% and 20.0%, respectively. The MIC90 of imipenem and meropenem was 16 and 32 mg/l, respectively (Table 1). In this study, most P. aeruginosa isolates were resistant to tetracycline (85.1%) and ceftriaxone (68.8%). About 20% to 35% of isolates were resistant to other antimicrobial agents such as amikacin, ciprofloxacin, piperacillin-tazobactam, cefepime, and ceftazidime.
MIC, minimum inhibitory concentration.
CSPA, colistin-susceptible P. aeruginosa.
CNPA, colistin-nonsusceptible P. aeruginosa.
CNPA, isolates (MIC, > 4 mg/l).
ND, not determined. Breakpoints are not available in CLSI. 4
The antimicrobial resistances of CNPA and CSPA isolates were compared, and the results are shown in Table 1. For all antimicrobial agents evaluated in this study, CSPA isolates showed higher antimicrobial resistance rates than CNPA isolates, and amikacin, ciprofloxacin, and meropenem resistance rates were different significantly between CSPA and CNPA isolates (p < 0.05). While 24.1% and 32.2% of CSPA isolates were resistant to amikacin and ciprofloxacin, respectively, no CNPA isolates were resistant to both (Table 1). For carbapenems, only one CNPA isolate was resistant to imipenem (MIC, 16 mg/l) and no CNPA isolates were resistant to meropenem. Fifty-one P. aeruginosa isolates (23.7%) were revealed to be MDR. Eight and 43 MDR isolates were from bacteremia and UTIs, respectively. That is, the MDR rate among P. aeruginosa isolates from UTIs was significantly higher than that among isolates from bacteremia (37.7% vs. 7.9%, respectively). All MDR isolates were susceptible to colistin.
Population analysis profiles
Population analysis was performed for four CNPA (two colistin-resistant and two colistin-intermediate isolates) and two CSPA isolates. As shown in Fig. 1, the four CNPA isolates showed growth over LOQ even at 10 mg/l of colistin. However, growth of the two CSPA isolates could not be detected at 2 or 6 mg/l of colistin. Thus, population analyses confirmed the colistin nonsusceptibility of some of the CNPA isolates. In addition, it suggested that further step mutants still more resistant to colistin could be potentially selected under treatment from CNPA isolates.
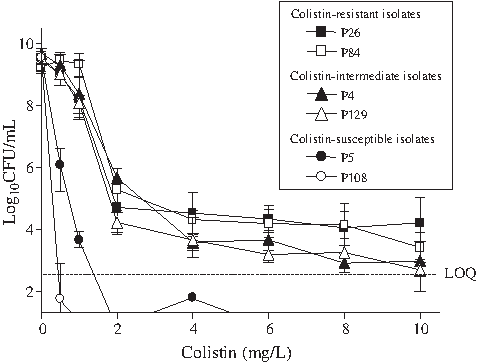
Population analysis profiles of colistin-nonsusceptible Pseudomonas aeruginosa isolates (P26 and P84, MIC = 8 mg/l; P4 and P129, MIC = 4 mg/l) and CSPA isolates (P5, MIC = 1 mg/l; P108, MIC = 0.5 mg/l). The Y-axis starts from the limit of detection, and the limit of quantitation (LOQ) is indicated in the figure.
Genetic backgrounds of CNPA isolates
MLST analysis revealed 12 STs among the 16 CNPA isolates, that is, ST207, ST244, ST292, ST319, ST389, ST488, ST527, ST641, ST968, ST969, ST970, and ST971 (Table 2). Of these STs, four STs (ST968 to ST971) were newly identified in this study. Four CNPA isolates (25.0%) were ST244, the most frequent genotype. All four ST244 CNPA isolates were from patients with bacteremia in the same hospital (SMC). In addition, three of these isolates showed the same antimicrobial resistance profile. Two ST292 CNPA isolates were found, and these isolates were from two different hospitals. The other 10 STs were represented by one isolate.
R, resistant; I, intermediate.
SMC, Samsung Medical Center; CNUH-G, Chonnam National University Hospital; JNUH, Jeju National University Hospital.
PB, Polymyxin B; IMI, Imipenem; MRP, Meropenem; AMK, Amikacin; CPM, Cefepime; CRO, Ceftriaxone; CAZ, Ceftazidime; CIP, Ciprofloxacin; TET, Tetracycline; P/T, Piperacillin-tazobactam; S, susceptible; I, intermediately resistant; R, resistant.
Discussion
The rapid increase in MDR has resulted in increased use of colistin and polymyxin B to treat serious P. aeruginosa infections. Although colistin and/or polymyxin B resistance has been reported in P. aeruginosa from several countries,11,23 the resistance rates for these two compounds are still relatively low on a worldwide scale. 11 In addition, colistin resistance in P. aeruginosa has rarely been reported in Asian countries, including Korea. However, emergence of colistin resistance has been documented in other Gram-negative pathogens in Korea, such as A. baumannii and K. pneumoniae. In this study, we isolated colistin-resistant (1.4%) and colistin-nonsusceptible (7.4%) P. aeruginosa isolates from patients in Korean hospitals with bacteremia or UTIs. Although the frequency of colistin resistance in P. aeruginosa isolates was lower than that in Acinetobacter spp. and K. pneumoniae isolates, the emergence of CNPA isolates in several hospitals is of concern. It is likely that the recent increase in use of colistin and polymyxin B in a clinical setting is responsible for the increasing rates of colistin resistance in Gram-negative bacteria. Consistent with this hypothesis, recent reports documented that colistin resistance arose during treatment of patients with colistin.8,12 However, we have no information whether the patients carrying CNPA isolates specifically received prior polymyxin therapy or whether the use of polymyxins was more intensive in SMC with higher rate of CNPA (10 out of 16 isolates) than in other hospitals.
Consistent with previous studies of A. baumannii and K. pneumoniae,17,22 we found that CNPA had a lower antimicrobial resistance rate than CSPA, especially against meropenem, amikacin, and ciprofloxacin (p < 0.05). Notably, coresistance to colistin and carbapenems (imipenem and meropenem) was rare in this study, although about 20% of P. aeruginosa isolates were resistant to carbapenems. It has been speculated that changes in the outer membrane associated with colistin resistance may affect susceptibility to other antimicrobial agents. 14 In addition, it was reported that the complete loss of LPS production may mediate the colistin resistance in A. baumannii. 19 However, no experimental data on the changes of outer membrane and LPS production in colistin-resistant P. aeruginosa isolates are currently available. Identification of the exact mechanism underlying colistin resistance and determination of the relationship between colistin resistance and resistance to other antimicrobial agents would inform the selection of therapeutic regimens and facilitate the development of new antimicrobial agents.
MLST analysis showed that most of the CNPA isolates occurred independently. Independent occurrence of colistin resistance has also been demonstrated in A. baumannii and K. pneumoniae.21,22 Independent occurrence of colistin resistance implies that colistin resistance may be acquired repeatedly, which may be related to the use of colistin. P. aeruginosa can exhibit adaptive resistance linked to LPS modification under certain environmental conditions such as subinhibitory concentrations of antimicrobial agents. 7 Thus, appropriate doses and intervals of colistin treatment of P. aeruginosa infections are critical to prevent the emergence and spread of colistin resistance.
Clonal dissemination of CNPA isolates was also evident in this study. That is, four ST244 CNPA isolates from blood were found in a single hospital (SMC) and two ST292 CNPA isolates from urine were identified in two different hospitals (SMC and CNUH-G). So far (as of July 19, 2010), seven ST244 P. aeruginosa isolates have been deposited in the MLST database (www.pubmlst.org/paeruginosa): six isolates from Europe and one isolate from China. Further, three ST244 isolates were from UTIs. That is, ST244 may be disseminating through Europe and Asia. Although there is no report of this clone acquiring colistin resistance in other studies, the acquisition of colistin resistance due to dissemination of this clone would be of great concern. Other STs such as ST319, ST292, ST641, and ST488 have been reported from countries in Europe such as Austria, the Czech Republic, and the Netherlands. In addition, ST207 and ST389 were reported from Canada and Australia, respectively. That is, CNPA isolates from Korea may belong to clones disseminating worldwide. Although no data are available concerning what clones have colistin resistance in other countries, our data suggest that colistin resistance may not be restricted to a few clones. In addition, acquisition of colistin resistance may be a frequent event related to inappropriate dosages and intervals of colistin treatment. Thus, the relationships between colistin resistance and the appropriate dosage and interval of colistin treatment should be investigated to prevent the spread of colistin resistance.
Footnotes
Acknowledgments
The authors thank members of the Korean Network for Studies of Infectious Diseases (KONSID) for their collaboration. P. aeruginosa isolates used in this study were obtained from the Asian Bacterial Bank (ABB) of the Asia Pacific Foundation for Infectious Diseases (APFID, Seoul, Korea).
Disclosure Statement
No competing financial interests exist.