
Abstract
A retrospective study was conducted to determine the prevalence of plasmid-mediated quinolone resistance (PMQR) determinants in uropathogenic Escherichia coli isolated from inpatients and outpatients in a teaching hospital of northern Italy. The presence of qnrA, qnrB, qnrS, aac(6′)-Ib-cr, and qepA was evaluated in 76 and 72 nalidixic acid-resistant E. coli, isolated in 2004 and 2006, respectively. Positivity for the aac(6′)-Ib-cr gene was demonstrated in 3 of 76 (3.9%) and 8 of 72 (11%) isolates, respectively; no other PMQR determinant was found. All aac(6′)-Ib-cr-positive strains also showed two point mutations in the gyrA and parC genes. Most aac(6′)-Ib-cr-positive isolates demonstrated the contemporary presence of blaCTX-M-15, blaOXA-1/30, and blaTEM-1 genes and 4/11 harbored a class 1 integron with a dfrA17-aadA5 gene cassette arrangement. Interestingly, all aac(6′)-Ib-cr-positive isolates belonged to B2 phylogenetic group, O25b antigen type, multi locus sequence type 131, and to a cluster of approximately 70% similarity level by pulsed-field gel electrophoresis (PFGE). These findings suggest the circulation of the previously described intercontinentally spreading E. coli O25:H4-ST131 clone in our geographical area since 2004. Hybridization studies of the PFGE profiles showed the aac(6′)-Ib-cr gene to be associated with different molecular weight bands (40–350 kb) and interestingly aac(6′)-Ib-cr chromosomal integration was demonstrated in one strain by I-Ceu I method. This represents the first report to investigate the presence and diffusion of PMQR determinants in northern Italy and to describe aac(6′)-Ib-cr chromosomal integration in E. coli.
Introduction
The plasmid-mediated quinolone resistance (PMQR) qnr determinants were first described in Klebsiella pneumoniae in 1998 25 and further characterized in 2002. 44 These groups of genes (qnrA, qnrB, qnrS, qnrC, and qnrD), sharing 40%–60% sequence homology, confer resistance by binding to DNA-gyrase and topoisomerase IV, thereby protecting them from the lethal gyrase-DNA-quinolone cleavage complex.8,27,38,45–47
Robicsek et al. described, for the first time in 2006, a gene variant of the aminoglycoside acetyltransferase enzyme, called aac(6′)-Ib-cr, responsible for resistance to kanamycin, amikacin, and tobramycin, from Enterobacteriaceae strains isolated in China in 2003.38,39 This gene variant has been described to confer bacterial resistance not only to aminoglycosides but also to some fluoroquinolones by modifying the chemical structure of the unsubstituted N group of the piperazinic ring in C7 position. This aminoglycoside acetyltransferase variant represents today the only known single mechanism of enzymatic drug modification able to confer antimicrobial resistance to two different classes of antibacterial agents.
The AAC(6′)-Ib-cr enzyme, one of the 30 known variants of AAC(6′)-Ib enzyme, is characterized by two point mutations: Trp102Arg and Asp179Tyr.
The acetyltransferase action of this enzyme has an effect against some fluoroquinolones but not others. This enzyme, responsible for approximately three- to fourfold reduction in the susceptibility to norfloxacin and ciprofloxacin, shows an additive effect with other plasmid-encoded resistance mechanisms. 39
The aac(6′)-Ib-cr gene has been described predominantly in Escherichia coli, 30 frequently associated with other antimicrobial resistance genes (such as extended-spectrum beta-lactamases [ESBL] genes).
More recently, a third PMQR determinant, the quinolone efflux pump qepA gene, has been identified.5,32,33,52
In Europe, many authors2,4,7,15,16,18,20,23,35 have reported the dissemination of PMQR determinants in Enterobacteriaceae and non-Enterobacteriaceae strains, although no data for Italy was reported in a recent review. 42
In Italy, only two previous studies have shown the presence of PMQR genes in E. coli; Perilli et al. 34 described the simultaneous presence of aac(6′)-Ib-cr, qnrB1, and blaCTX-M-15 resistance determinants in one E. coli strain isolated from a urine sample of a nonhospitalized patient in Italy. More recently, Cerquetti et al. described the presence of PMQR genes in 64 extraintestinal pathogenic E. coli isolated in Rome in 2006 showing the presence of only the aac(6′)-Ib-cr gene and of aac(6′)-Ib-cr together with qnrB1 in 12 (18.8%) and 2 (3.1%) of isolates, respectively 9 ; moreover, most of these strains were also shown to produce CTX-M-15 and to belong to the recently reported intercontinental spreading multi locus sequence type (MLST) 131 clone of E. coli.11,26
The aim of this study was to conduct a retrospective analysis on a selection of uropathogenic E. coli strains collected during 2004 and 2006 to evaluate the prevalence and diffusion of PMQR genes in an area of northern Italy (Monza).
Nalidixic acid resistance was arbitrarily chosen to screen isolates for the presence of PMQR determinants, this being the least active quinolone.
In particular, this study focused on the detection of qnrA, qnrB, qnrS, aac(6′)-Ib-cr, and/or qepA in nalidixic acid-resistant isolates of E. coli.
Materials and Methods
Bacterial isolates
Two collections of 235 and 240 consecutive nonduplicate strains of uropathogenic E. coli isolated at San Gerardo Hospital of Monza between November and December 2004 and during the same period in 2006, respectively, were analyzed.
The distribution of strains isolated from urine samples of outpatients and inpatients was 116 (49.4%) versus 119 (50.6%) in 2004 and 155 (64.6) versus 85 (35.4%) in 2006.
Seventy-six and 72 nalidixic acid-resistant E. coli, isolated in 2004 and 2006, respectively, were further analyzed to evaluate the presence of PMQR determinants.
Antibiotic susceptibility determinations
The identification and preliminary susceptibility of bacterial strains to different antimicrobial agents were determined by the hospital Microbiology Laboratory using Vitek 2 Compact (BioMérieux). The in vitro susceptibility of E. coli isolates shown to be nalidixic acid-resistant (Clinical and Laboratory Standards Institute [CLSI] breakpoints: susceptible ≤16 μg/mL; resistant ≥32 μg/mL) was determined to amoxicillin-clavulanate (GlaxoSmithKline SpA), ceftazidime, cefotaxime, aztreonam, norfloxacin, ciprofloxacin, levofloxacin, tobramycin, gentamicin (Sigma-Aldrich), ulifloxacin, the active compound of the lipophilic prodrug prulifloxacin (Angelini ACRAF SpA; Aprilia), and trimethoprim-sulfametoxazole (Roche; SpA) by broth microdilution minimum inhibitory concentration (MIC) determination in accordance with the Clinical and Laboratory Standards Institute (CLSI; formerly National Committee for Clinical Laboratory Standards) guidelines. 13 The breakpoints for resistance were those recommended by the CLSI. 12 For ulifloxacin, the proposed European susceptibility breakpoints were as follows: susceptible ≤1 μg/mL, intermediate 2 μg/mL, resistant ≥4 μg/mL. 17 E. coli American Type Culture Collection (ATCC) 25922 was used as control strain.
Gene amplification and sequencing
DNA extraction
Bacterial DNA was extracted by Genomic DNA kit by Nucleospin (Macherey-Nagel) according to the manufacturer's recommendations.
PMQR genes
Clinical isolates were screened for the plasmid-mediated resistance genes qnrA, qnrB, qnrS, aac(6′)-Ib-cr, and qepA.
The presence of qnr genes (qnrA1, qnrB1, and qnrS1 and their variants) was determined using a modified polymerase chain reaction (PCR)-based technique as described by Cattoir et al., 6 whereas for the determination of the presence of the qepA gene, a PCR assay as described by Liu et al., 21 was performed. The quality control strains used for these determinations were E. coli Lo positive for qnrA1, K. pneumoniae B15 positive for qnrB1, and E. cloacae LEM positive for qnrS1 kindly donated by Prof. P. Nordmann. A qepA-positive control strain was supplied by Prof. P. Courvalin. The most recently described qnrC 47 and qnrD 8 genes were not included in this study, as they had not yet been described when the study was planned.
The presence of aac(6′)-Ib gene was determined by a PCR assay followed by enzymatic digestion using BstF5I and confirmed by sequencing of the amplicon to discriminate the -cr variant, as described by Park et al. 30 The positive control strain used for this determination was an internal clinical isolate proven to be positive for aac(6′)-Ib-cr.
QRDR of gyrA and parC
Mutations in gyrA and parC genes were determined by PCR amplification and subsequent sequencing as described by Weigel et al. 49 The E. coli ATCC 25922 reference strain with wild-type topoisomerase genes served as sequencing control.
Beta-lactamase determination
The aac(6′)-Ib-cr-positive isolates were further investigated for the presence of blaTEM, blaSHV, blaCTX-M, and blaOXA genes by PCR using specific primers as previously reported36,29,40 and confirmed by sequencing. ESBL production was confirmed by a standard double-disk synergy test. 29
Phylogenetic classification
The aac(6′)-Ib-cr-positive isolates were classified according to E. coli phylogenetic groups A, B1, B2, and D by multiplex PCR, as previously described. 10
O25b typing
The O25b antigen type was determined by a PCR-based method as previously described by Clermont et al. 11 for all aac(6′)-Ib-cr-positive isolates. This O antigen type has been associated to the recently described intercontinental emergence of a highly virulent CTX-M-15 ESBL producing and ciprofloxacin-resistant E. coli, characterized by belonging to the B2 phylogenetic subgroup and MLST profile ST131. The control strain used for this determination was E. coli UR40/R: ST131, aac(6′)-Ib-cr and CTX-M-15-positive, kindly donated by Dr. A. Carattoli. 9
Detection of class I integrons
To identify the presence of class 1 integrons in aac(6′)-Ib-cr-positive isolates, a CS-PCR (conserved-sequence PCR) assay of crude bacterial lysate was performed, using primers for the 5′-CS and 3′-CS region, according to the method described by Levesque et al. 19
Sequencing analysis
Each PCR product that appeared as a unique size when observed on an agarose gel was purified (mix of Exonuclease I+ Phosphatase A) and used directly for sequencing. Sequencing was performed by “Consorzio per la Genetica Molecolare Umana” sequencing facility of the University of Milano-Bicocca, Monza, on an Abi Prism 3130 DNA Analyzer (Applied Biosystems) using a Big Dye Terminator ver. 3.1 sequencing kit (Applied Biosystems) and a Sequencing Analysis ver. 5.3 program (Applied Biosystems) for data analysis. DNA sequences were visually inspected and edited with Chromas (Technelysium Pty Ltd) and BioEdit (Ibis Biosciences), ver. 7.0.9.0. ClustalX was used to perform multiple alignment analyses of the sequences. All nucleic acid sequences were compared against cassette sequences of human uropathogenic E. coli isolates deposited in the National Center for Biotechnology Information database by the BLAST program.
MLS-typing
Gene amplification and sequencing of seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were performed according to the recommendations of the E. coli MLST Web site (http://mlst.ucc.ie/mlst/dbs/Ecoli). 50 Allele and sequence type (ST) were determined following the Web site instructions.
PFGE analysis
Pulsed-field gel electrophoresis (PFGE) analysis using the standardized E. coli (O157:H7) protocol established by Centers for Disease Control was performed on all the tested strains to evaluate their clonal distribution. Genomic DNA of each nalidixic-resistant E. coli was analyzed by PFGE after digestion with XbaI (Fermentas Inc.; M-Medical). DNA fragments were separated by electrophoresis in 1% agarose in 0.5 × Tris Borate EΔTA buffer with CHEF–DR II apparatus (Bio-Rad) at 14°C, 6 V/cm, and with alternating pulses at a 120° angle in a 5–35 sec pulse time gradient for 20 hr. A phage lambda DNA ladder (50–1,000 kb Pulse marker, Sigma-Aldrich) was used as a size marker. Interpretation of PFGE patterns was based on the criteria of Tenover et al. 43
Southern blot hybridization
For Southern blot analysis, DNA fragments present in electrophoresed PFGE agarose gel were transferred to a Hybond N+ membrane (GE Healthcare) and cross-linked with UV light. Subsequently, the membrane was hybridized with a specific radiolabelled probe using standard procedure. 24 The probe used for the hybridization was the same PCR-generated amplicon used for aac(6′)-Ib-cr detection. The PCR product was purified (MinElute PCR Purification Kit, Qiagen) and radiolabeled by random priming (Rediprime II; GE Healthcare). After hybridization the membrane was exposed to Kodak XAR films to generate radiograms.
Plasmid determination and sizing
The detection and sizing of plasmids present in strains carrying PMQR determinants was performed using incubation with S1 nuclease (Sigma-Aldrich) followed by a PFGE analysis as previously described by Barton et al. 3 S1 nuclease converts supercoiled plasmids into full-length linear molecules, which migrate appropriately according to their molecular weight and can be distinguished from chromosomal genome after ethidium staining. E. coli UR40/R strain, kindly donated by Dr. A. Carattoli and coworkers, 9 was run as control as it was known to posses 3 different molecular weight plasmids (48, 100 and 170 kb).
Chromosomal localization of the aac(6′)-Ib gene by PFGE-I-Ceu I
To confirm the chromosomal location of the aac(6′)-Ib-cr gene, genomic DNA of E. coli was digested with the I-Ceu I endonuclease (New England Biolabs) according to Liu et al. 22 An aac(6′)-Ib-cr-negative strain (E. coli 3236) was used as a control. The digested fragments were separated by PFGE using the CHEF–DR II system (Bio-rad) as previously described. The I-Ceu I restriction fragments were subjected to Southern hybridization with two different PCR-generated probes for aac(6′)-Ib-cr gene and for the chromosomal housekeeping parC gene, 49 the latter used as positive control.
Statistical analysis
Differences between groups were assessed for statistical significance using the Chi-square test or Fisher's exact test when appropriate. All tests were carried out two-sided. A p-value of ≤0.05 was considered statistically significant.
Results
Presence of PMQR determinants
None of the 76 and 72 nalidixic acid-resistant E. coli, isolated in 2004 and 2006, respectively, were found to be positive for the qnr gene variants investigated or for the plasmidic qepA gene.
The positivity for the aac(6′)-Ib-cr variant was confirmed in three and eight nalidixic acid-resistant E. coli isolated in 2004 and 2006, respectively, with an overall prevalence of 7.4%, increasing from 3.9% (3/76) to 11.1% (8/72) in all nalidixic acid-resistant E. coli (p = 0.0883) during the study time.
Presence of QRDR determinants
All strains found to be positive for PMQR determinants, represented in our study by the presence of aac(6′)-Ib-cr gene, were screened for quinolone-resistance determining region (QRDRs). These isolates were shown to have multiple point mutations in the gyrA and parC genes. In particular, all strains showed the contemporary presence of Ser83Ile and Asp87Asn mutations in GyrA enzyme and Ser80Ile and Glu84Val mutations in ParC enzyme (Table 1).
Indicates the molecular weight of plasmid that hybridizates in the Southern blot assay.
NOR, norfloxacin; CIP, ciprofloxacin; LEV, levofloxacin; ULI, ulifloxacin; AMC, amoxicillin-clavulanate; CAZ, ceftazidime; FUR, cefuroxime; ATM, aztreonam; TOB, tobramycin; GEN, gentamicin; SXT, trimethoprim-sulfamethoxazole; MIC, minimum inhibitory concentration; QRDR, quinolone-resistance determining region.
Antimicrobial susceptibility
MIC distributions of norfloxacin, ciprofloxacin, levofloxacin, and prulifloxacin for nalidixic acid-resistant E. coli strains are shown in Table 2.
Susceptibility testing of PMQR-positive E. coli strains revealed MIC values ranging from 8 μg/mL to >128 μg/mL for the four studied fluoroquinolones; norfloxacin and ciprofloxacin shared higher MIC values as compared with that of levofloxacin and prulifloxacin.
All aac(6′)-Ib-cr-positive strains were resistant to amoxicillin/clavulanate, cefuroxime, and tobramycin; 8/11 (72.7%) were resistant to ceftazidime and aztreonam; 4/11 (36.4%) to trimethoprim-sulfamethoxazole, and 5/11 (45.5%) to gentamicin (Table 1).
Presence of beta-lactamase genes
In Enterobacteriaceae isolates, aac(6′)-Ib-cr has been linked to ESBL genes. 18 In this study, a total of 10 of 11 (90.9%) aac(6′)-Ib-cr-positive E. coli confirmed an ESBL phenotype by double-disk synergy test. BlaOXA-1/30 was found among all aac(6′)-Ib-cr-positive strains, whereas blaCTX-M-15 was shown in 10 of 11 isolates (90.9%). BlaTEM-1 (8/11; 72.7%) was always detected in copresence with blaCTX-M-15 and blaOXA-1/30. None of the strains were seen to possess blaSHV genes. These results are shown in Table 1.
Presence and analysis of class 1 integrons
Class 1 integrons were detected in only 4 (36.4%) of 11 aac(6′)-Ib-cr-positive uropathogenic E. coli isolates. The arrangement of the detected gene cassette was characterized by the presence of a 1.8 kb two-gene dfrA17-aadA5 element (Table 1).
Phylogenetic classification, O25b typing, and MLST profile
All studied aac(6′)-Ib-cr-positive uropathogenic E. coli were shown to belong to the B2 phylogenetic group, to possess the O25b antigen type, and to belong to the MLST profile ST131.
PFGE profiles
The PFGE analysis of aac(6′)-Ib-cr-positive E. coli strains demonstrated the presence of several clonal types with most of the isolates showing only approximately 70% relatedness (Fig. 1 and Fig. 2).
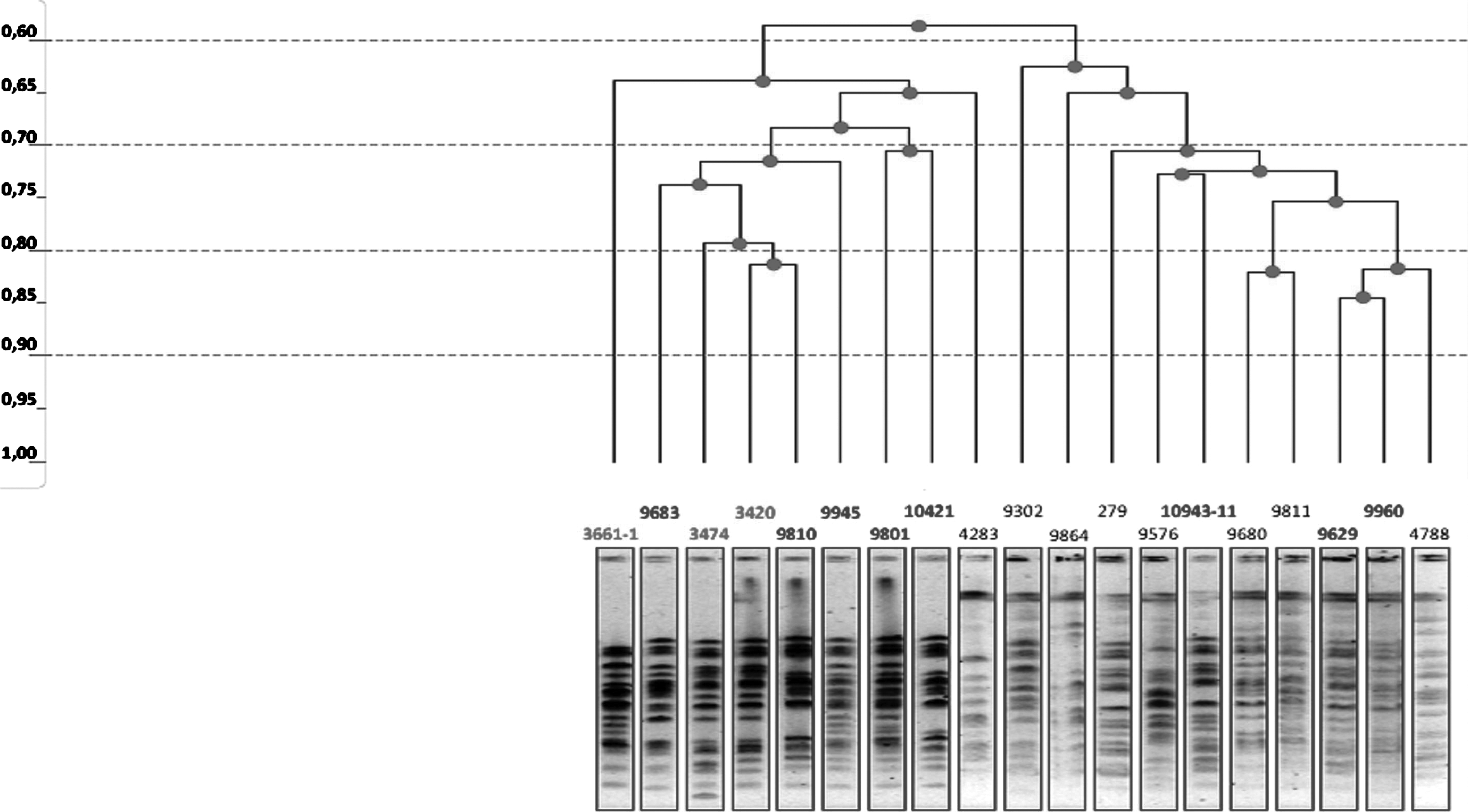
Dendrogram showing the similarity among aac(6′)-Ib-cr-positive Escherichia coli strains isolated in 2004 (3420, 3474, and 3661-1) and in 2006 (9629, 9683, 9801, 9810, 9945, 9960, 10421, and 10943-11) compared with aac(6′)-Ib-negative isolates (279, 4283, 4788, 9302, 9576, 9680, 9811, and 9864). PFGE profiles were generated by XbaI digestion of genomic DNA. The dendrogram was constructed utilizing the Bio-rad Quantity One software and unweighted pair group method with arithmetic mean algorithm. PFGE, pulsed-field gel electrophoresis.
Plasmid determination and sizing
The aac(6′)-Ib-cr-positive strains were all found to possess plasmids that ranged in number, according to their molecular weights, from one to two varying in size from 48 to 250 kb (Table 1 and Fig. 3).
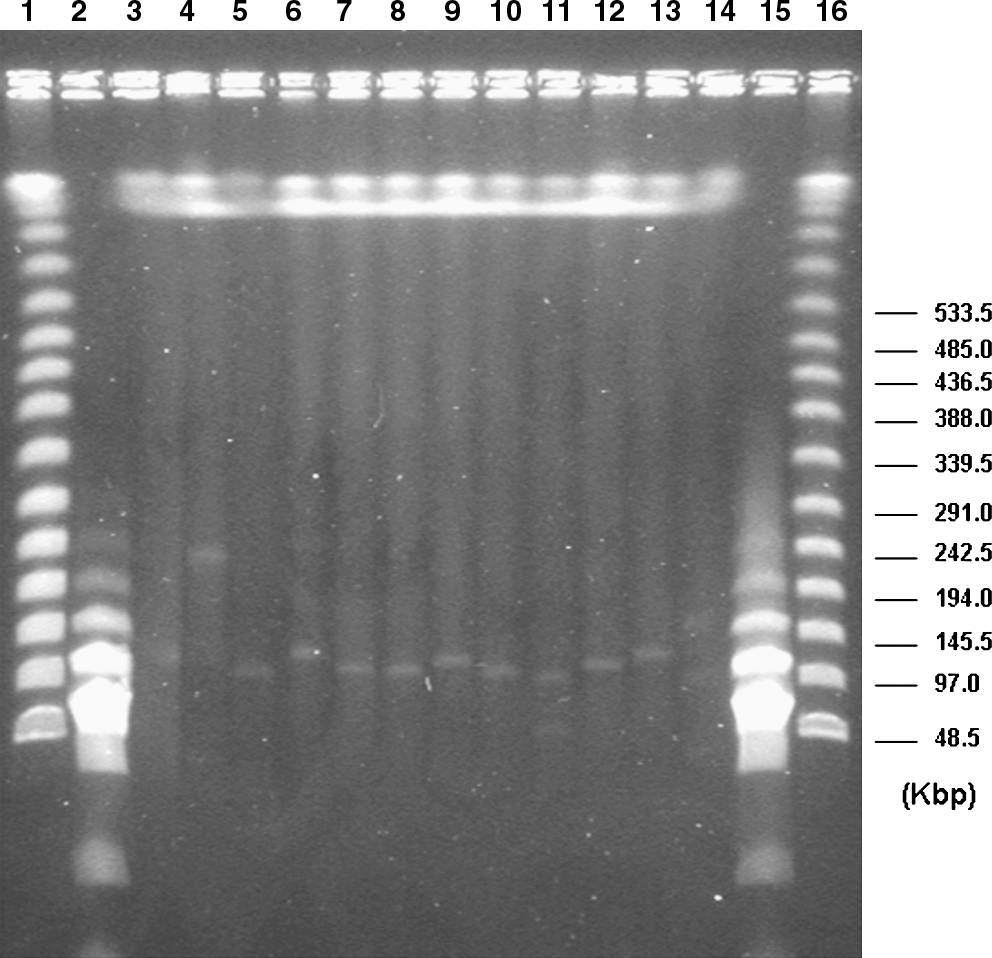
Plasmid profile obtained after S1 nuclease restriction and PFGE electrophoresis. Lanes 1 and 16: Marker 48.5–1,000 kb (Sigma-Aldrich). Lanes 2 and 15: Marker 0.1–100 kb (Sigma-Aldrich). Lane 3 (3420), 4 (3474), 5 (3661-1), 6 (9629), 7 (9683), 8 (9801), 9 (9810), 10 (9945), 11 (9960), 12 (10421), and 13 (10943-11): aac(6′)-Ib-cr-positive E. coli strains. Lane 14 (UR40/R): E. coli strain kindly donated by Dr. A. Carattoli and coworkers 9 and used as control.
Southern blot
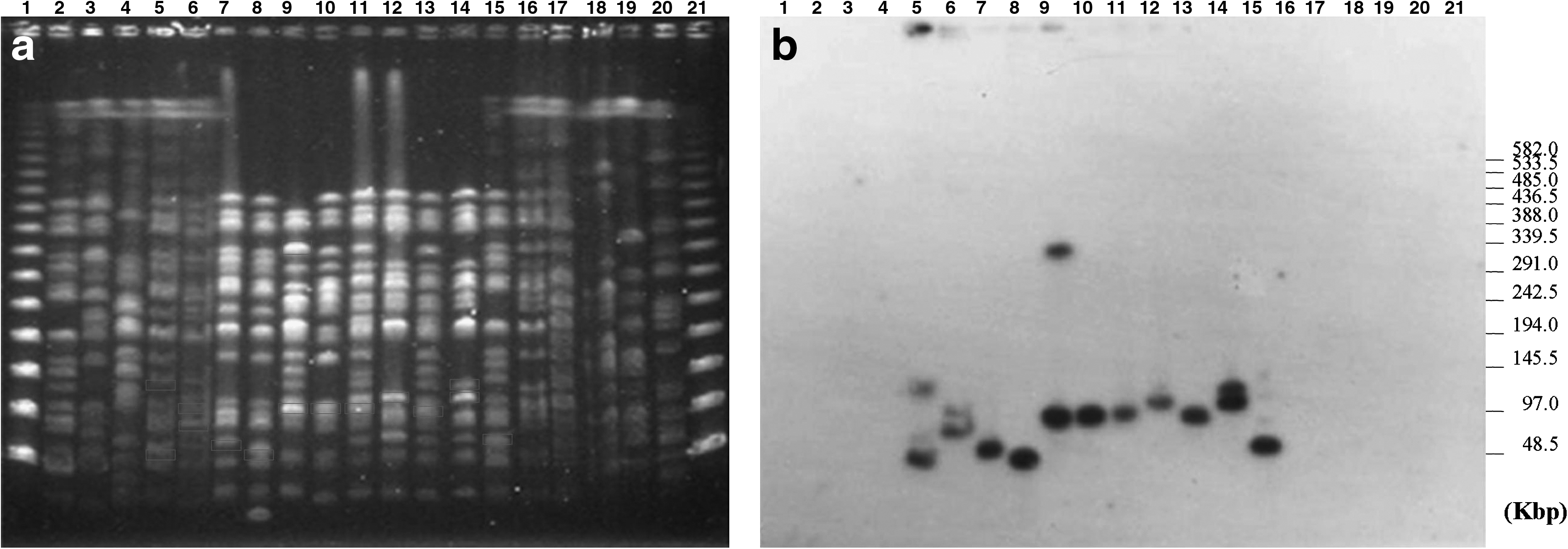
Southern blot hybridization analysis of XbaI PFGE DNA profiles of aac(6′)-Ib-cr-positive E. coli showed that the aac(6′)-Ib-cr gene was located on bands of different molecular weights corresponding to approximately 48–120 kb (Table 1; Fig. 2a and b).
However, E. coli strain 3661–1, isolated in 2004, showed an additional hybridization band of 350 kb in its PFGE profile. This higher molecular weight band was present in the XbaI PFGE profile of this strain but not in its S1 nuclease plasmid profile (Fig. 3). The integration of aac(6′)-Ib-cr within this bacterial chromosome was confirmed by PFGE I-Ceu I hybridation. Both aac(6′)-Ib-cr and parC probes were found to hybridize with the same endonuclease I-Ceu I 820 kb fragment (Fig. 4).
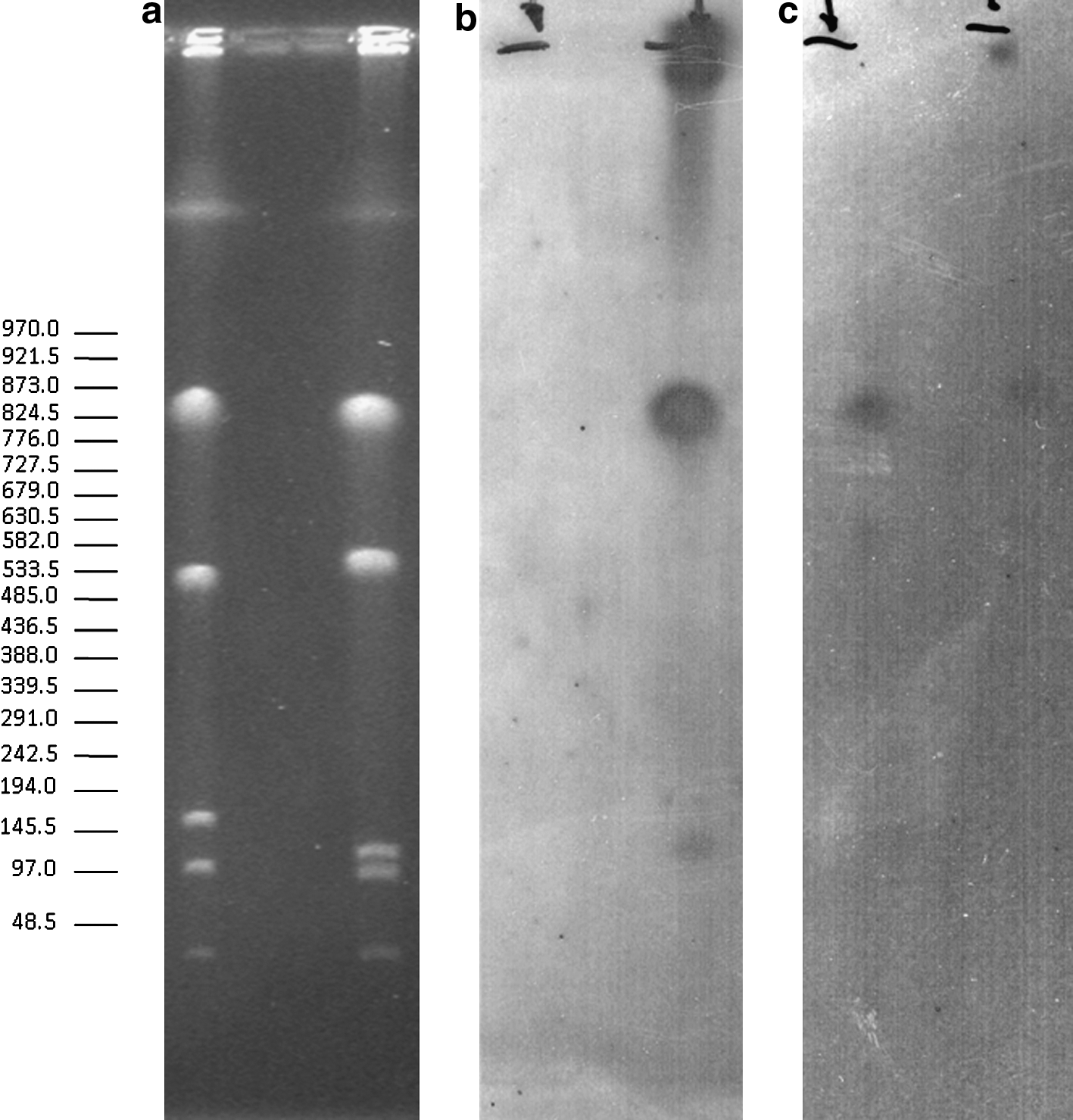
Chromosomal location of the aac(6′)-Ib-cr gene.
Discussion
In the last decade, several PMQR mechanisms have been discovered and described worldwide, 40 showing a high prevalence in both clinically relevant and commensal strains of enterobacteria. Little data are, however, available on the epidemiology of PMQRs in clinical strains of Enterobacteriaceae isolated in Italy.
This retrospective study demonstrated that aac(6′)-Ib-cr is the only PMQR determinant present in nalidixic acid-resistant uropathogenic E. coli strains isolated in inpatients and outpatients from a large teaching hospital of northern Italy during 2004 and 2006. Further, aac(6′)-Ib-cr-positive E. coli isolates, selected for nalidixic acid resistance, increased from 3.9% in 2004 to 11.1% in 2006. Although statistical significance was not reached (p = 0.088), these results may indicate a raise in the prevalence of this gene in isolates from northern Italy.
These findings are in keeping with previous reports which indicated that aac(6′)-Ib-cr may be more prevalent than other plasmid-mediated determinants, such as Qnr proteins, particularly among clinical strains of E. coli.39,42,48 In particular, the recent study by Warburg et al., investigating the presence of qnr and aac(6′)-Ib-cr genes in a collection of nosocomial blood stream isolates of E. coli collected from 1991 through 2005 at a tertiary-care center in Jerusalem, demonstrated that aac(6′)-Ib-cr was the predominant PMQR gene in E. coli, showing an increase in prevalence from 1998 to 2005, which coincided with a rise in fluoroquinolone resistance from 11% in 1997 to 37% in 2005. 48 Further, this study demonstrated qnrA to be present in only a single E. coli strain, out of the 718 strains analyzed, in spite of the concomitant circulation of qnr genes in K. pneumoniae and Enterobacter spp. populations described in the same hospital. 41 In Slovenia, the aac(6′)-Ib-cr gene was also reported in Klebsiella spp. isolates; its prevalence increased from 5.8% in 2000–2002 to 42% in 2003–2005. 2
Even though the copresence of aac(6′)-Ib-cr and qnr genes in E. coli strains has been widely reported in literature, some studies have also reported the non contemporary presence of aac(6′)-Ib-cr and of qnr genes in E. coli. 28 Moreover, Cerquetti et al. 9 reported the aac(6′)-Ib-cr gene to be present predominantly alone in ciprofloxacin-resistant E. coli strains isolated in Italy.
Multiple point mutations in gyrA and parC genes were also demonstrated in all aac(6′)-Ib-cr-positive E. coli strains, in keeping with the high level resistance to ciprofloxacin and other fluoroquinolone observed in these isolates. PMQR genes are in fact known to determine only a moderate decrease in susceptibility to fluoroquinolones, but to predispose to high level resistance by greatly increasing mutant selection. 39 It would be interesting in future studies to determine whether PMQR genes are present in apparently susceptible bacterial populations to confirm that the acquisition of plasmid-mediated determinants is a primary event in the development of fluoroquinolone resistance. Moreover, the strain selection criteria used in this study to determine the presence of PMQR determinants may have underestimated the presence of aac(6′)-Ib-cr-positive isolates, as nalidixic acid is not a substrate for AAC(6′)-Ib-cr enzyme. In future studies tobramycin resistance may represent a more useful strategy for selecting possible aac(6′)-Ib-cr-positive strains, as it is the most sensitive among aminoglycosides to this variant of the acetyltrasferase enzyme.
The presence of this PMQR determinant in the strains analyzed was shown to be frequently associated to other plasmid-encoded resistance genes such as blaOXA1/30, blaCTX-M-15, and blaTEM-1. This is in accordance with results of previous studies that have described the frequent association of PMQR with multiple antimicrobial resistance genes, particularly aac(6′)-Ib-cr and blaCTX-M, generally housed within mobile genetic elements such as class I integrons.14,23,29 Class 1 integrons were, however, detected in only 36.4% of studied strains; their particular gene cassette arrangement was characterized by a 1.8 kb fragment containing only dfrA17-aadA5 genes but not aac(6′)-Ib-cr. Similar findings have been described in uropathogenetic E. coli.1,31,51
PFGE analysis showed the presence of several clonal types among the aac(6′)-Ib-cr-positive E. coli strains studied; this is again in keeping with previous reports.42,48 Almost all the aac(6′)-Ib-cr-positive E. coli strains were also shown to be CTX-M-15 ESBL producers and to belong to the B2 phylogenetic subgroup and O25b antigen type. Moreover, MLST profile determination confirmed all aac(6′)-Ib-cr-positive strains to belong to E. coli ST131, suggesting the circulation of the previously described intercontinentally spreading clone O25:H4-ST1319,26,51 in our geographical area since 2004.
Hybridization of PFGE profiles using aac(6′)-Ib-cr probe showed the presence of this gene within a number of heterogeneous molecular weight bands in studied strains. Further, chromosomal integration of the aac(6′)-Ib-cr gene was confirmed in one E. coli strain, isolated in 2004, by performing I-Ceu I method. Interestingly, Cerquetti et al. also recently described the chromosomal integration of another plasmid-encoded resistance gene, blaCTX-M-15, in extra-intestinal strains of E. coli isolated in Rome in 2006 and belonging to ST131 clone. 9 The possibility that some of these mobile genetic elements may have at some point integrated in the bacterial chromosome, as described for other bacterial species, 42 raises particular concern regarding the possible clonal diffusion of genetically more stable antibiotic-resistant strains.
In conclusion, this study demonstrates the diffusion in our geographical area of uropathogenic E. coli characterized by the presence of the aac(6′)-Ib-cr gene, as the only PMQR determinant, in association with ESBL genes such as blaCTX-M-15. Moreover, all aac(6′)-Ib-cr positive strains were shown to belong to B2 phylogenetic group, O25b antigenic type, and MLST type 131. Furthermore, this is, to our knowledge, the first report demonstrating the chromosomal integration of aac(6′)-Ib-cr determinant in E. coli.
Footnotes
Acknowledgments
The authors thank Prof. P. Nordmann and Dr. L. Poirel for generously donating their isolates E. coli Lo positive for qnrA1, K. pneumoniae B15 positive for qnrB1, and E. cloacae LEM positive for qnrS1; Prof. P. Courvalin, who kindly provided QepA-positive strain; and Dr. A. Carattoli, who generously provided O25b, ST131 E. coli–positive strain, used as control. The authors thank Dr. Donato Migliarino for his contribution to experimental results. This work was partially supported by a grant to C.C. from MURST ex 60% (Ministero dell'Istruzione, dell'Università e della Ricerca, Italy).
Disclosure Statement
No competing financial interests exist.