
Abstract
Abstract
The objective of this study was to examine the distribution of multidrug resistance in Escherichia coli in relation to plasmid replicon types, animal sources, and genotypes. E. coli isolates (n = 35) from seven different animal sources were selected and tested for susceptibility to 15 antimicrobials; pulsed-field gel electrophoresis was used to determine genetic relationships among the E. coli isolates. Plasmid types based on their incompatibility (Inc) replicon types were determined, and linkage disequilibrium analysis was performed for antimicrobial resistance profiles, replicon types, and animal source. A high degree of genotypic diversity was observed: 34 different pulsed-field gel electrophoresis types among the 35 isolates examined. Twelve different plasmid Inc types were detected, and all isolates carried at least one replicon type. IncF (n = 25; 71.4%) and IncFIB (n = 19; 54.3%) were the most common replicon types identified. Chloramphenicol resistance was significantly linked with four Inc types (A/C, FIIA, F, and Y), and amoxicillin/clavulanic acid was linked with three Inc types (B/O, P and Y). Resistance to any other antimicrobial was linked to two or fewer replicon types. The isolate source was linked with resistance to seven antimicrobials and IncI1. We conclude that commensal E. coli from animal sources are highly variable genotypically and are reservoirs of a diverse array of plasmids carrying antimicrobial resistance.
Introduction
It appears that plasmids encoding antimicrobial resistance genes are the main factors affecting MDR in E. coli.2,4 These mobile genetic elements are readily acquired by E. coli and can thereby confer resistance to one or more antimicrobials. Resistance plasmids can be transferred between strains of the same species or different genera (such as Salmonella or Klebsiella), and such exchanges may serve to establish reservoirs of resistance genes in animals and the environment. 26
Plasmids encode sequences called replicons that are required for self-replication in a host cell, including the origin of replication, control of initiation, and replication functions.3,7 Plasmid incompatibility (Inc) types have been defined based on the inability of different plasmids with incompatible replicons to coexist in the same cell.7,20 In the Enterobacteriaceae, 27 Inc or replicon types have been identified and a PCR-based replicon typing assay has been developed that distinguishes 18 of the 27 most common Inc types. 5
Certain replicon types are associated with MDR and with bacteria implicated in disease outbreaks or found in food-producing animals.12,14 For example, MDR Salmonella have been responsible for human outbreaks and may be acquired through food animals that harbor zoonotic pathogens.22,28,31 However, much less is known about the range and types of resistance plasmids in E. coli that confer MDR phenotypes. Instead, most reports on E. coli have focused on the relationships of specific plasmid-borne resistance genes, particularly those identified with extended-spectrum β-lactamases (e.g., Marcade et al. 17 and Mshana et al. 18 ) and the replicon types of these vectors. This study was designed to characterize a small set of E. coli isolates representative of seven common animal sources for their antimicrobial resistance phenotypes, pulsed-field gel electrophoresis (PFGE) genotypes, and plasmid replicon types to learn more about the distribution of MDR in relation to plasmids based upon replicon types.
Materials and Methods
Bacterial strains and plasmids
Escherichia coli isolates were obtained from National Antimicrobial Resistance Monitoring System (NARMS) (www.ars.usda.gov/Main/docs.htm?docid=6750) and the Bacterial Epidemiology and Antimicrobial Resistance Research Unit (BEAR-RU) culture collection.
This collection is comprised of isolates from studies conducted by the USDA or collaborators from the National Animal Health Monitoring System (NAHMS), the Regional Dairy Quality Management Alliance (RDQMA), the USDA Food Safety Inspection Service (USDA FSIS), and other studies. A total of 35 isolates collected during 2005–2008 from seven different sources, including beef cattle, chicken, dairy cattle, dog, domestic cat, horse, and swine, were selected for this study (Table 1). In addition to host source, isolates were chosen based on resistance to β-lactam antimicrobials (amoxicillin-clavulanic acid, ampicillin, cefoxitin, and ceftiofur) and to tetracycline. Three pan-susceptible isolates were also included. Positive controls used in the replicon typing procedure were originally developed in the laboratory of Werner K. Maas5,7 and generously provided by Alessandra Carattoli (Istituto Superiore di Sanita, Rome, Italy). All bacterial strains were stored at −80°C in LB lennox (Hardy Diagnostics, Santa Maria, CA) with 15% glycerol or stored at room temperature on tryptic soy agar (Hardy Diagnostics) slants.
Antimicrobials: AUG, amoxicillin/clavulanic acid; AMP, ampicillin; FOX, cefoxitin; TIO, ceftiofur; AXO, ceftriaxone; CHL, chloramphenicol; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; STR, streptomycin; SMX, sulfamethoxazole; TET, tetracycline; COT, trimethoprim/sulfamethoxazole.
The different plasmid replicon types detected in each isolate.
Antimicrobial susceptibility testing
The E. coli isolates were tested for susceptibility to a panel of 15 antimicrobials using the Sensititer™ system (Trek Diagnostic Systems Inc., Westlake, OH) as described by NARMS. Antimicrobials included amikacin (AMI), amoxicillin/clavulanic acid (AUG), ampicillin (AMP), cefoxitin (FOX), ceftiofur (TIO), ceftriaxone (AXO), chloramphenicol (CHL), ciprofloxacin (CIP), gentamicin (GEN), kanamycin (KAN), nalidixic acid (NAL), streptomycin (STR), sulfamethoxazole (SMX), tetracycline (TET), and trimethoprim/sulfamethoxazole (COT). Each isolate was classified as susceptible, intermediate, or resistant using Clinical and Laboratory Standards Institute (CLSI) breakpoints when available. 19 Otherwise, breakpoint interpretations from the NARMS program were used. For the linkage analyses described below, antimicrobials identified as intermediate were considered susceptible.
Plasmid replicon typing multiplex PCR
E. coli isolates were examined for the presence of 18 plasmid replicon (Inc) types by polymerase chain reaction (PCR) using three multiplex primer panels as described by Johnson et al. 14 This procedure is a modified version of the eight PCR multiplex and simplex reactions described by Carattoli et al. 5 PCR reactions were performed as previously described by Lindsey et al. 16 and consistent with the AmpliTaq DNA polymerase manufacturer's instructions (Applied Biosystems, Foster City, CA).
Pulsed-field gel electrophoresis
All isolates were analyzed at the USDA VetNet Laboratory (Athens, GA) using the standardized 24–28 hours E. coli PFGE protocol developed at the Centers for Disease Control and Prevention by the PulseNet program. 23 Briefly, genomic DNA was digested with 10 U of XbaI (Roche Molecular Biochemicals, Indianapolis, IN). The resulting DNA fragments were analyzed with the CHEF-Mapper XA PFGE system (Bio-Rad, Hercules, CA) in 0.5 × Tris-borate-EDTA buffer at 14°C at 6 V for 18 hours with a ramped pulse time of 2.16–63.8. The BioNumerics software program (Applied Maths, Austin, TX) was used for cluster analysis using the Dice coefficient, 1.5% band tolerance, 1.5% optimization, and the unweighted pair group method with arithmetic mean.
Statistical analysis
Linkage disequilibrium (LD) was calculated as an extension of Fisher's exact probability test on contingency tables 25 using the program Arlequin. 8 Standard settings were used: 10,000 steps in the Markov chain and 1,000 dememorization steps; calculations of D, D′, and r 2 coefficients were made using a significance level of 0.05.
Results
The 35 isolates for use in this study (Table 1) were selected based primarily on antimicrobial resistance and also on animal host source. The isolates originated from seven different animal sources: chicken (n = 10), swine (n = 9), dog (n = 6), beef cattle (n = 4), dairy cattle (n = 4), domestic cat (n = 1), and horse (n = 1). This represented the majority of E. coli host sources in the NARMS and BEAR-RU collection, and was among those recently added to the collection (2005–2008). Since resistance to tetracycline was used to select isolates likely to be carrying plasmids, all but the three pan-susceptible isolates were included in this group (n = 32). β-lactam resistance was also a main factor in choosing the study isolates. Accordingly, 32 isolates were ampicillin resistant, 31 isolates were resistant to amoxicillin/clavulanic acid, 30 isolates were cefoxitin resistant, and 28 isolates were resistant to ceftiofur; only three isolates were resistant to another cephem, ceftriaxone. Aminoglycoside resistance was significant among the isolates with 17 resistant to streptomycin, 10 resistant to gentamicin, and 7 resistant to kanamycin. Resistance to sulfa drugs was also identified: 18 isolates were resistant to sulfamethoxazole, whereas only four were resistant to trimethoprim/sulfamethoxazole. Of particular note, a significant portion of the isolates were resistant to quinolone antimicrobials with 12 resistant to nalidixic acid and 9 resistant to ciprofloxacin. Also, although chloramphenicol was not among the antimicrobials considered when selecting the isolates for the study, nearly one-third (n = 10) were chloramphenicol resistant. All of the isolates in this study, except the three pan-susceptible isolates, were resistant to three or more antimicrobials (Table 1).
Between one and six different plasmid Inc types were found in each isolate (Table 1). Overall, 12 of the 18 different plasmid replicon types tested were identified among the 35 isolates in the study. None of the isolates carried IncFIC, IncHI2, IncL/M, IncN, IncT, or IncX. The three most prevalent replicon types were IncF (n = 25), IncFIB (n = 19), and IncA/C (n = 9). Other replicon types often detected included IncFIA (n = 8), IncB/O (n = 7), IncI1 (n = 7), IncW (n = 7), IncP (n = 6), and IncY (n = 5). In contrast, only two isolates had an IncK plasmid, and IncFIIA and IncHI1 were each found in just one isolate. However, all four of these isolates were resistant to the antimicrobials used to select the isolates: amoxicillin/clavulanic acid, ampicillin, cefoxitin, ceftiofur, and tetracycline (Table 1). Of the 25 IncF positives, 21 isolates were resistant to these five antimicrobials, and 16 of the 19 IncFIB-positive isolates were resistant to all five of the drugs. The six isolates carrying the IncP type had the lowest percentage (n = 4) positive for the five selecting antimicrobials among the 12 plasmid replicon types identified. Also, the number of different Inc types identified in a particular isolate was not associated with the degree of MDR. For instance, an isolate (ID #487) identified with only IncA/C was resistant to 12 antimicrobials, whereas the isolate with the most Inc types (ID# 393) (n = 6) was resistant to 9 antimicrobials. Similarly, an isolate resistant to just three drugs (ID# 5) carried four different plasmid replicon types.
To obtain a more complete picture of the significance of the relationships between antimicrobial resistance and plasmid Inc type, linkage disequilibrium (LD) analysis was performed. Using this method, the significance of the associations between the replicon types identified and resistance to the selecting antimicrobials revealed significant linkage (p < 0.05) between amoxicillin/clavulanic acid resistance and IncB/O, IncP and IncY (Fig. 1). Resistance to the other four selecting drugs was only significantly linked with either IncB/O (ampicillin and tetracycline) or IncP (cefoxitin and ceftiofur). IncF was linked with chloramphenicol, kanamycin resistance, and trimethoprim/sulfamethoxazole, whereas IncFIB was not significantly linked with resistance to any of the antimicrobials tested. IncFIA and IncW were also not linked to any resistance, and IncI1 was linked only with resistance to streptomycin. In contrast, IncA/C was significantly linked with resistance to six antimicrobials: chloramphenicol, ciprofloxacin, kanamycin, naladixic acid, streptomycin, and sulfamethoxazole. Although the LD procedure cannot distinguish specific linkages among the seven isolate sources in this study, a correlation between IncI1 and isolate animal source was identified and isolate source was also linked with resistance to seven antimicrobials (Fig. 1).
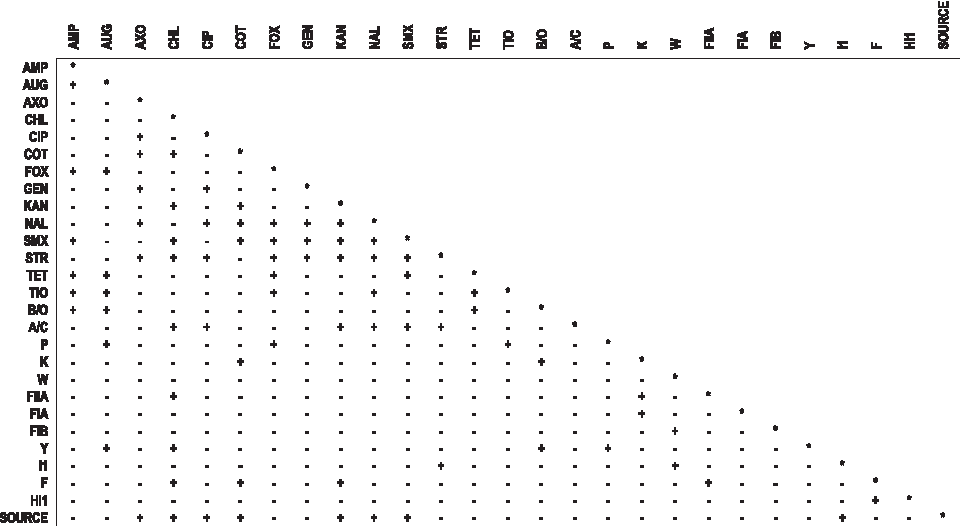
Pair-wise linkage disequilibrium among antimicrobial resistance profiles, 12 replicon types, and source animal of Escherichia coli isolates. (+) denotes a p-value of 0.05 or less and indicates significant linkage; (−) denotes a p-value of greater than 0.05 and no significant linkage.
PFGE analysis was conducted on all 35 E. coli isolates. Cluster analysis based on PFGE of the 35 isolates showed 34 unique types, some of which were less than 70% similar, suggesting a genotypically variable population (Fig. 2). The source animal of the isolate did not appear to have a correlation to cluster analysis.

Pulsed-field gel electrophoresis-based dendrogram, resistance profiles, replicon typing, and isolate source. Dendrogram was constructed based on pulsed-field gel electrophoresis analyses using Bionumerics. For antimicrobial resistance profiles (columns 1–14), black blocks indicate resistant and white indicates intermediate or susceptible. For plasmid replicon types (columns 15–26), black blocks indicate the trait is present and white indicate the trait is absent. Antimicrobials: amoxicillin/clavulanic acid (AUG), ampicillin (AMP), cefoxitin (FOX), ceftiofur (TIO), ceftriaxone (AXO), chloramphenicol (CHL), gentamicin (GEN), kanamycin (KAN), nalidixic acid (NAL), streptomycin (STR), sulfamethoxazole (SMX), tetracycline (TET), and trimethoprim/sulfamethoxazole (COT).
Discussion
This study was designed to characterize a small set of E. coli isolates representative of the most varied animal sources available to us for their antimicrobial resistance phenotype, plasmid replicon type, and PFGE genotypes to learn more about the distribution of MDR in relation to plasmids based upon replicon types. Cluster analysis of the PFGE patterns determined that the isolates selected were genotypically quite diverse. Linkage analysis was performed for antimicrobial resistance profiles, plasmid replicon types, and isolate source animal, revealing some significant correlations between MDR, plasmids, and animal source.
The detailed nature of the phenotypic and genotypic analysis allowed the investigation of only 35 isolates; however, they were selected to represent common animal sources and yielded informative data from a genotypically diverse group of MDR E. coli. Care must be taken not to over interpret or extrapolate the data; nevertheless, some interesting correlations were observed. Several of the plasmid Inc replicon types were found to have significant linkage disequilibrium with each other and with antimicrobial resistance phenotypes. For example, IncF was significantly linked with IncFIA and IncH1, and IncFIB was linked with IncW, indicating that these linked plasmid types are coresident in the same isolates or may be present on multireplicon plasmids. Similarly, IncB/O was linked with IncK and IncY, suggesting that these replicon types may also be found in the same E. coli isolate.
Although the replicon types IncF and IncFIB were identified in most of the E. coli isolates in our study, they were only linked to a few antimicrobials. This is not surprising because the linkage disequilibrium method cannot detect significant associations if a character occurs frequently in the isolate test population. For instance, Bean et al. 1 and Wu et al. 30 both reported that in E. coli IncB/O is associated with sulfonamide resistance as well as ampicillin and streptomycin resistance, but our results differed from these findings. However, IncB/O plasmids are associated with MDR and also implicated as carriers of virulence factors in E. coli, 13 adding to the potential importance of this replicon type.
The plasmid types IncFIA and IncI1 were also frequently identified in our study. Although IncI1 was only linked with chloramphenicol resistance and IncFIA was not significantly linked with resistance to any of the antimicrobials tested, previous studies have identified these replicon types as major factors in conferring β-lactam resistance in E. coli.6,9,12,17,18 IncFIA is also considered to be important carrier of virulence factors in E. coli. 13
IncP, IncW, and IncY were identified in 14%–20% of the isolates in our study, and, as previously reported for E. coli, 13 all three replicon types were associated with MDR in our isolate group. Of particular note, IncP had significant linkage with resistance to the β-lactamase antimicrobials amoxicillin/clavulanic acid, cefoxitin, and ceftiofur; IncY was also linked to amoxicillin/clavulanic acid resistance.
All of the E. coli isolates in this study that carried the IncA/C plasmids were MDR, which is consistent with the findings of Welch et al. 29 and Lindsey et al. 16 in their studies on Salmonella enterica animal isolates. We found that the IncA/C-positive E. coli isolates were resistant to seven or more antimicrobials and all were resistant to the same six antimicrobials (ampicillin, ceftiofur, ciprofloxacin, sulfamethoxazole, streptomycin, and tetracycline). The IncA/C type was linked to six antimicrobials (chloramphenicol, ciprofloxacin, kanamycin, nalidixic acid, streptomycin, and sulfmethoxazole), twice the number of antimicrobials compared to the other 11 Inc types we identified. This suggests a prominent role for IncA/C plasmids as carriers of antimicrobial resistance and virulence factors in E. coli, an implication supported by recent reports.13,17
The report by Lindsey et al. 16 identified IncA/C and IncI1 as the predominant replicon types in the diverse population of Salmonella animal isolates they studied, and we also found similar percentages of E. coli isolates with these Inc types in our study. They did not identify any isolates carrying IncFIC, IncK, IncT, or IncW. We did not detect IncFIC or IncT in our varied panel of E. coli isolates, but we did identify IncK and IncW. Moreover, while IncFIB and IncF were the main replicon types in our E. coli study, they represented only a small percentage of the Inc types found in Salmonella. 16 Although differences exist, important similarities are found among E. coli and Salmonella plasmid replicon types, and these Inc types may have a prominent role in conferring antimicrobial resistance or other traits in E. coli. 27
In conclusion, the present study showed a high prevalence of multiple plasmid replicon types present in a limited number of genetically diverse E. coli isolated from a variety of animal species. As in Salmonella, IncA/C was found in MDR isolates, which indicates an important role for this replicon type in antimicrobial resistance in E. coli. The PFGE genotypes of the study population were very diverse and did not segregate according to plasmid replicon types or animal sources of the E. coli isolates. Additional research examining a larger E. coli isolate population for further linkages with antimicrobial resistance, plasmid Inc types, and virulence factors associated with Shiga toxin-producing E. coli is warranted.
Footnotes
Acknowledgments
The authors thank Alessandra Carattoli for replicon typing control strains, and Takiyah Ball, Beth McGlinchey, Carolina Hall, Jovita Haro, Georgina Hildago, Sandra House, and Alice Wilcher for technical assistance.
Disclaimer
The mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.