
Abstract
Staphylococcus epidermidis have become important causes of nosocomial infections, as its pathogenesis is correlated with the ability to form biofilms on polymeric surfaces. Production of poly-N-acetylglucosamine (PNAG) is crucial for S. epidermidis biofilm formation and is synthesized by the gene products of the icaADBC gene cluster. Production of PNAG/polysaccharide intercellular adhesin and biofilm formation are regulated by the alternative sigma factor, σB, and is influenced by a variety of environmental conditions including disinfectants and other antimicrobial substances. The susceptibility of five S. epidermidis strains to antibiotics alone and in double combination was previously tested. Our results demonstrated that some combinations are active and present a general broad spectrum against S. epidermidis biofilms, namely rifampicin–clindamycin and rifampicin–gentamicin. In the present study, it was investigated whether the combination of rifampicin with clindamycin and gentamicin and these antibiotics alone influence the expression of specific genes (icaA and rsbU) of S. epidermidis within biofilms using real-time polymerase chain reaction. The data showed that in most cases the expression of both genes tested significantly increased after exposure to antimicrobial agents alone and in combination. Besides having a similar antimicrobial effect, rifampicin combined with clindamycin and gentamicin induced a lower expression of biofilm-related genes relatively to rifampicin alone. Associated with the advantage of combinatorial therapy in avoiding the emergence of antibiotic resistance, this study demonstrated that it can also cause a lower genetic expression of icaA and rsbU genes, which are responsible for PNAG/polysaccharide intercellular adhesin production, and consequently reduce biofilm formation recidivism, relatively to rifampicin alone.
Introduction
The tight pathogenic association is essentially linked to the species' ability to form adherent biofilms on artificial surfaces, 30 which is the main virulence mechanism of S. epidermidis. Biofilm-associated bacteria are 100–1,000 times less susceptible to antibiotics than are planktonic cells. 6 A number of factors have been postulated as mechanisms of biofilm-associated antibiotic resistance, including binding of antibiotics to the slime, poor penetration of antibiotic into the biofilm, slow growth rate of organisms in the biofilm, high bacterial density, and change in gene expression in biofilm bacteria.6,33
Antibiotic combinations are being increasingly used as therapeutic option as a result of the increasing appearance of multiresistant microorganisms. 21
Many S. epidermidis strains produce poly-N-acetylglucosamine (PNAG), also named polysaccharide intercellular adhesin (PIA) homopolymer, a crucial factor for biofilm formation, which surrounds and connects S. epidermidis cells in a biofilm. 20 Synthesis of PIA is essential for bacterial cell accumulation because it mediates cell-to-cell adhesion of proliferating cells. 12 In addition to its role as part of the extracellular biofilm matrix, PNAG/PIA has been found to protect the bacterium from important mechanisms of innate host defense, namely neutrophil killing, complement deposition, immunoglobulins, and antimicrobial peptides.14,35 The correlation of the presence of PNAG/PIA with the invasiveness of the bacterium may be due to the roles of this exopolymer in biofilm formation and immune evasion. 26 Two main putative determinants of S. epidermidis invasiveness have been identified: the ica genes, which regulate the production of PNAG/PIA, and the insertion element IS256. The latter is thought to contribute to the genetic adaptation that may have a role during infection. 36 The biosynthesis of PNAG/PIA is accomplished by the gene products of the ica (intercellular adhesion) locus,8,10,20 which comprises four intercellular adhesion genes icaA, icaB, icaC, and icaD. 23 The expression of the ica operon and biofilm formation depends on a variety of environmental conditions. The expression of at least four unlinked genes (icaR, 7 σB, the purR locus, and sarA) is known to control PNAG/PIA synthesis at the level of transcription.3,18,19 Production of PNAG/PIA is subject to a range of regulatory influences, 24 including many global virulence regulators. PNAG/PIA and biofilm formation is regulated by the alternative sigma factor σ B , which is presumed to play a crucial role in the global regulation of gene expression. 11 RsbU is a positive regulator of the activity of σB, the general stress-response factor of Gram-positive microorganisms. 5 σB may act only indirectly via an additional, unknown factor or rsbU may, by itself, be a regulator of icaADBC transcription. 12
Activation of PNAG/PIA expression by different stress stimuli apparently uses different pathways. This implies that a number of environmental conditions and regulatory systems can influence the expression of staphylococcal biofilms, reflecting the magnitude of the complexity associated with biofilm formation. 27
In previous studies, we assessed the effect of various antibiotics alone and in double combination against S. epidermidis biofilms. Rifampicin combined with gentamicin and clindamycin showed to be the most active combinations, promoting reductions of about 3 log colony forming units (CFUs) in biofilm cells in the majority of clinical isolates tested. However, the effect of these treatments in the subsequent response of survival cells was never determined. Accordingly, the main goal of the present work was to assess the expression of some genes of S. epidermidis, namely icaA and rsbU, both responsible by PNAG/PIA production after antibiotic treatment.
Materials and Methods
Bacterial strains and antimicrobial agents
In this study, five good-biofilm–producing and ica-positive S. epidermidis strains were used: 117977, 132034, 150271, 1457, and 9142. The first three strains were kindly provided by Dr. Howard Ceri, Department of Biological Sciences, University of Calgary, Calgary, and the last two by Dr. G. B. Pier, Channing Laboratory, Department of Medicine, Brigham and Women's Hospital, Harvard Medical School, Boston. These strains are clinical isolates (isolated from infected catheters) and were stored at −80°C. Antibiotics tested were rifampicin, gentamicin, and clindamycin (Sigma) and were used at break-point concentration. Interpretation criteria for susceptibility testing were based on National Committee for Clinical Laboratory Standards, 22 now Clinical Laboratory Standards Institute (CLSI) guidelines (Table 1).
Concentration used in bactericidal antibiotic test.
NCCLS, National Committee for Clinical Laboratory Standards.
Biofilm formation
Several colonies of the isolates grown on tryptic soy agar plates were suspended in saline (0.9% NaCl) to a density of 1.0 on the McFarland scale, as indicated by the manufacturer. Then the bacterial suspension was resuspended in tryptic soy broth to obtain a cellular concentration of circa 1 × 107 CFU/ml. This solution was used as inoculum for the MBEC™ device (MBEC Biofilm Technologies Ltd.). The biofilms were grown for 48 hours at 37°C and 150 rpm on a rocking platform where the shear force was created against the pegs forming 96 equivalent biofilms. At the end of the incubation period, a biofilm of approximately 6 logs per peg was formed.
Biofilm challenge and recovery
The challenge plates were prepared using the antibiotics at break-point concentration (Table 1) alone and in combination (rifampicin + clindamycin and rifampicin + gentamicin). The biofilms formed on the lid of the MBEC were rinsed twice with 0.9% saline and placed into the challenge plate overnight at 37°C, 150 rpm on a rocking platform, and 95% relative humidity. Then, the challenged biofilms were rinsed twice in saline and transferred to a recovery plate that consisted of tryptic soy broth medium plus 1% Tween. Biofilms were removed from all pegs at once by sonication for 8 minutes on high with an Aquasonic sonicator (model 250HT; VWR Scientific). 2 The vibration disrupted biofilms from the surface of the 96 pegs into the recovery plate. Then, CFUs were determined as follows: the recovery medium (containing the sonicated biofilms) was serially diluted. The biofilm cultures (10-fold diluted) were spotted on tryptic soy agar plates. Then the plates were incubated for 48 hours at 37°C to ensure maximum recovery of the surviving microorganisms and CFUs were counted. The same protocol was repeated and all samples were collected, preserved in RNAlater® solution (Ambion-Applied Biosystems), and kept at −80°C for further study of gene expression.
Genetic expression analysis
Primer design
Primers used for S. epidermidis icaA and rsbU genes analysis by real-time polymerase chain reaction (PCR) were designed using Primer3 web-based software (http://fokker.wi.mit.edu\cgi-bin\primer3\primer3_www.cgi) 31 and are listed in Table 2. To verify the specificity of each primer pair for its corresponding target gene, PCR products were first amplified from 1457 strain genomic DNA (data not shown).
RNA extraction
Total RNA of each sample was extracted using the PureLink™ RNA Mini Kit (Invitrogen) according to the manufacturer's recommended protocol. Potential DNA contamination was removed during RNA purification procedure by On-column PureLink DNase treatment (Invitrogen). RNA concentration (ng/μl) and purity (OD260nm/OD280nm) were assessed by spectrophotometric measurements using a NanoDrop device (NanoDrop 1000 Spectrophotometer, V3.6.0; Thermo Fisher Scientific, Inc.).
cDNA synthesis
To ensure equivalent starting amounts of RNA from control and respective treated samples to be converted into cDNA, appropriate dilutions in RNase-free water were performed to obtain a final concentration of 2 μg of RNA. cDNA of each sample was synthesized using the iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories, Inc.). Each reaction contained 2.5 μl of iScript Reaction Mix +iScript Reverse Transcriptase and 7.5 μl of RNA template, in accordance with the proportions recommended by the kit manufacturer, making a final reaction volume of 10 μl. Complete reaction mix was incubated in a thermocycler (MyCycler; BioRad) with the following reaction protocol: 5 minutes at 25°C, 30 minutes at 42°C, and 5 minutes at 85°C.
Quantitative real-time PCR
Real-time PCRs were performed on a CFX96TM real-time PCR Detection System (Bio-Rad Laboratories, Inc.). Each 20 μl of reaction mixture contained 2 μl of cDNA, 1 μl of each primer (forward and reverse primers), 10 μl of 1 × SsoFast™ EvaGreen® Supermix (Bio-Rad Laboratories, Inc.), and 6 μl of nuclease-free water. Thermal cycling conditions were as follows: 3 minutes initial denaturation at 95°C, followed by 40 cycles of 5 seconds denaturation at 95°C, 10 seconds annealing/extension at 60°C (this step was performed at 60°C, concerning primers efficiency previously determined [data not shown]). A melt curve was inserted at the end of each run, with readings from 65°C to 95°C, every 1°C for 5 seconds, to confirm that only the desired product was amplified.
Gene analysis and expression
Samples for real-time PCRs were run in triplicate. Data were analyzed using the CFX Manager™ version 1.6 (Bio-Rad Laboratories, Inc.) and the relative quantification method (2−ΔΔCT; Livak and Schmittgen), 17 which describes the change in expression of the target genes relative to the 16S rRNA reference genes from untreated control samples,15,34 and the cycle threshold values (CT; cycle at which each sample amplification curve crosses a specific threshold) were averaged for triplicate samples. The ΔCT values of the target genes were determined by normalizing to the endogenous control genes 16S rRNA. These samples were subsequently subtracted from the 16S rRNA genes of the untreated control samples. The ΔΔCT was used to calculate relative expression using the following formula: 2−ΔΔCT.9,16,17
The cDNA was validated by comparing with an RNA sample not subjected to reverse transcriptase reaction, to determine possible DNA contamination. All these no-reverse transcriptase controls showed a ΔΔCT above 10 cycles, indicating that the cDNA present resulted from the conversion of the RNA.
Statistical analysis
The data were analyzed by applying two-tailed independent samples t tests. SPSS software was used to perform the analysis. All tests were performed with a confidence level of 95%.
Results
We have previously demonstrated that some combinations of antibiotics are significantly effective against S. epidermidis biofilm cells. Rifampicin alone was generally as or more effective than when used in combination with other antibiotics (Fig. 1). However, this kind of combinatorial therapy aims to prevent the emergence of resistance usually associated with monotherapy. Two of the most efficient combinations that induced a greater reduction of bacterial biofilm population and demonstrated a broader spectrum range, being active against all strains tested, were rifampicin plus clindamycin or gentamicin. Both these combinations were able to reduce bacterial cell number by approximately 3 log (Fig. 1) for all strains tested.

Effect of rifampicin (Rif), clindamycin (Clind), and gentamicin (Gent) alone and in combination (Rif + Clind; Rif + Gent) on 48-hour-old biofilm cells of five Staphylococcus epidermidis clinical isolates, after overnight exposure. Δ Log reduction = differences between positive control (untreated control: 6 log10 colony forming units/peg) and treated samples in log10 colony forming units/peg. Error bars represent standard deviation. *Combinations statistically different from rifampicin alone (p < 0.05).
To determine the effect of rifampicin, clindamycin and gentamicin, and rifampicin combined with clindamycin or gentamicin on gene expression of S. epidermidis icaA and rsbU genes, real-time PCR analysis was performed. The results obtained for the different strains tested are represented in Fig. 2.
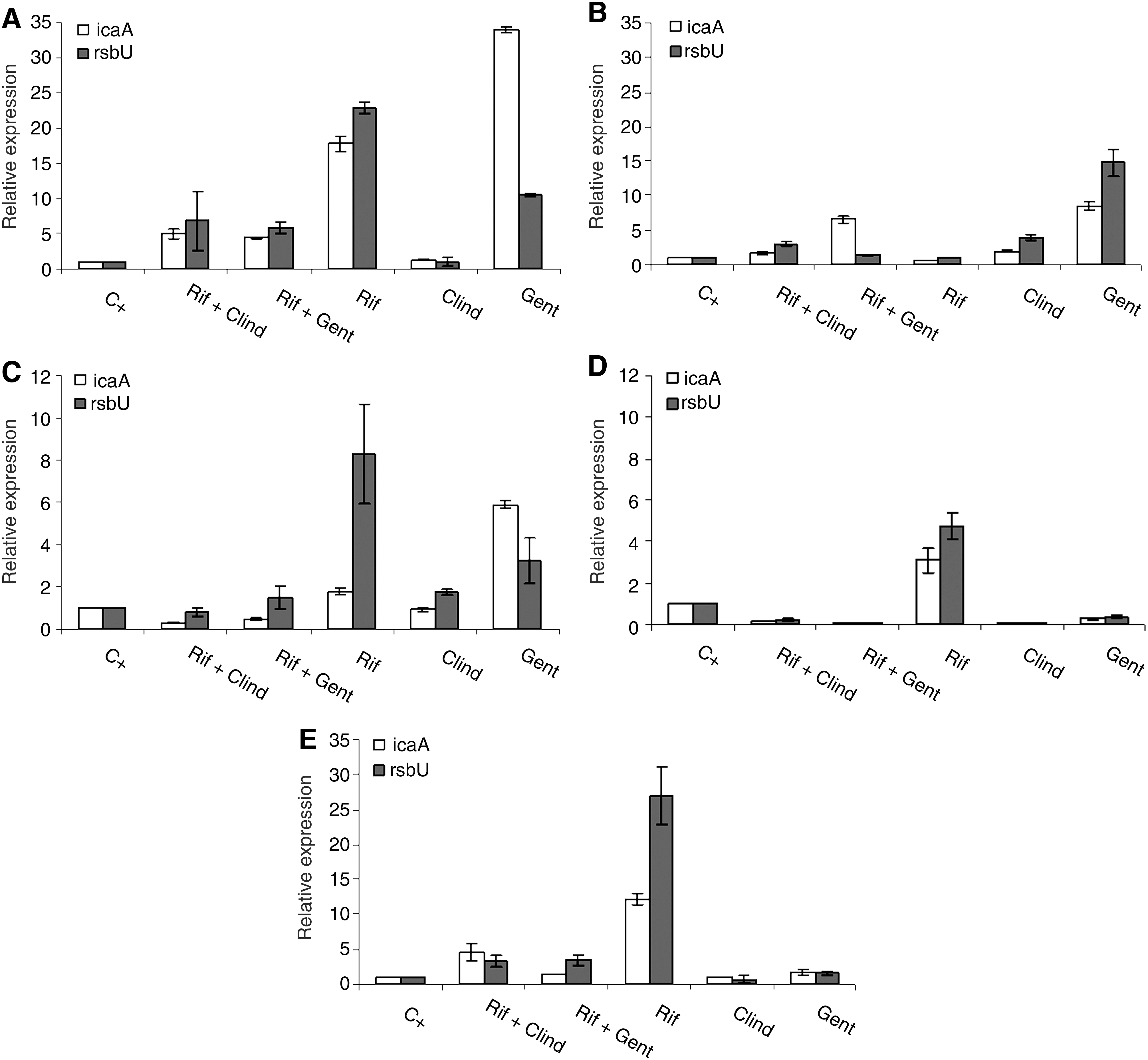
Expression of icaA and rsbU in S. epidermidis strains
Relative to the unexposed control, icaA and rsbU were overexpressed in all strains when exposed to rifampicin at break-point concentration (p < 0.05), except for strain 132034 (Fig. 2B). The same was observed for gentamicin, which caused the overexpression of both genes in four strains (p < 0.05), with the exception of 1457 S. epidermidis. Generally, no overexpression of icaA and rsbU genes was observed in biofilm cells exposed to clindamycin. Relatively to combinations of antibiotics, the response of S. epidermidis was strain dependent. Three strains demonstrated overexpression of both genes after exposure to rifampicin + clindamycin and rifampicin + gentamicin when compared with untreated biofilms (p < 0.05). On the contrary, these two combinations induced subexpression of icaA and rsbU on 1457 S. epidermidis. On the other hand, in strain 150271, both combinations provoked subexpression of icaA and the expression of rsbU was unaffected comparatively to the positive control (p < 0.05).
Comparing the effect of antibiotics alone and in combination, it was observed that the combinations always induced expression of icaA and rsbU genes in a lesser extent than antibiotics individually. As clindamycin and gentamicin are not active antimicrobial agents against S. epidermidis biofilms (Fig. 1), and rifampicin was the only antibiotic presenting similar effect to the most active combinations tested (ca. 3 log reduction), it seems important to compare the genetic expression of both genes after biofilm exposure to these antimicrobial treatments. Therefore, although both combinations of antibiotics induce an increase of icaA and rsbU expression in response to an environmental stress, this increase is not as marked as in cells exposed to rifampicin alone. In fact, comparing the icaA and rsbU gene expression after treatment with rifampicin and rifampicin combined with clindamycin or gentamicin, we can conclude that, with the exception of 132034, there is always a general decrease of the expression of these two genes after biofilm exposure to both combinations (p < 0.05) (Fig. 2).
Discussion
Our results showed that normally the expression of icaA is accompanied by the expression of rsbU. Pintens et al. demonstrated that the increase in icaA expression clearly preceded the increase in rsbU expression in vitro as well as in vivo. 27 This is consistent with factors other than sigB inducing icaADBC operon expression.3,18,19
RsbU is a positive regulator of the alternative sigma factor σB, which is known to control PNAG/PIA synthesis at the transcription level. Moreover, the global stress response regulator σB and rsbU control biofilm development in S. epidermidis. 12 The results clearly indicated that the expression of icaA and rsbU is generally inferior in the presence of break-point concentrations of rifampicin combined with clindamycin or gentamicin in comparison to rifampicin alone. Although all treatments studied induced the expression of the two genes assayed, after overnight contact, rifampicin is a stronger inducer of icaA and rsbU genes expression, possibly inducing in S. epidermidis persister cells a high ability for biofilm formation. In fact, biofilm formation has been reported to be induced by conditions that are potentially toxic for bacterial cells, such as high levels of osmolarity, detergents, urea, ethanol, oxidative stress, and the presence of sub-MICs of some antibiotics such as tetracycline and quinupristin–dalfopristin.4,13,28 In the present study, it was verified that a toxic condition, caused by rifampicin exposure, promoted an increased expression of icaA gene by surviving cells. In S. epidermidis, the induction of PNAG/PIA production and biofilm formation can improve the ability of biofilm immune evasion. PNAG/PIA is crucial for connection of cells in a biofilm, avoiding the detachment of biofilm cells, mechanism that can make the cells more susceptible to antimicrobial agents. Further, this exopolymer has also a protective function. Consequently, the induction of PNAG/PIA formation can be a possible defense and resistance mechanism of cells.
Therefore, in addition to the already known advantages of antibiotic combinatorial therapy, for example, as a strategy to reduce resistance development, our results show that the use of rifampicin combined with gentamicin and clindamycin may be a better therapy strategy and more effective than rifampicin alone (the most efficient antibiotic against S. epidermidis used in clinical practice). The combination therapy, in addition to allowing for an almost equally effective killing of bacterial cells, results in lower expression of icaA and rbsU, and consequently less PIA production, than if rifampicin was used alone, thereby ensuring a more efficient control of S. epidermidis biofilm-associated infections.
Footnotes
Acknowledgments
F. Gomes and P. Teixeira fully acknowledge the financial support of Fundação para a Ciência e Tecnologia through the grants SFRH/BD/32126/2006 and SFRH/BPD/26803/2006, respectively.
Disclosure Statement
No competing financial interests exist.