
Abstract
Historically regarded as a skin commensal, Staphylococcus epidermidis has been increasingly implicated in invasive foreign body infections such as catheter-related bloodstream infections, indwelling device infections, and prosthetic joint infections. We report a case of an aggressive, difficult-to-eradicate, invasive prosthetic hip infection occurring early after hardware implant and associated with a high-grade bacteremia and assess its salient molecular characteristics. The clinical and molecular characteristics of this isolate mirror the pathogenesis and persistence commonly seen with invasive methicillin-resistant S. aureus and may be attributed to the combination of resistance genes (SCCmec type IV), putative virulence factors (arcA and opp3a), cytolytic peptide production (α-type phenol-soluble modulins), and biofilm adhesion, interaction, and maturation (bhp, aap, and β-type phenol-soluble modulins).
Introduction
Generally viewed as an opportunistic pathogen, S. epidermidis has a biofilm-forming capacity, allowing it to elaborate an ecological niche and colonize biomaterials such as indwelling catheters, prosthetic heart valves, and other implanted devices. Adhesion factors, such as biofilm associated–homologous protein (bhp) and intercellular adhesin locus (ica) composed of the genes icaA, icaD, icaB, and icaC, which code for polysaccharide intercellular adhesin (PIA), play a putative role in the ability of S. epidermidis to persist, leading to difficulty in its eradication. Activation of the ica locus results in PIA synthesis and thick biofilm production. This can be inactivated by insertion sequences, specifically IS256, which reversibly disrupts the ica locus, leading to a phenotypic switch from biofilm-positive to -negative organisms. 22 In ica-negative strains, surface proteins such as the accumulation-associated protein (aap) have been shown to play a primary role in biofilm formation. 4
mecA is harbored on a mobile staphylococcal cassette chromosome (SCC) and encodes for an altered penicillin-binding protein (PBP2′) that imparts resistance to methicillin and other β-lactam antibiotics. 9 Neighboring the SCCmec element on the S. epidermidis chromosome is a large operon, the arginine catabolic mobile element (ACME), a virulence determinant encoding arginine deaminase pathway genes, including arcA and oligopeptide permease (opp3a). 6 The SCCmec-ACME association, previously described in USA300 S. aureus, was also noted in S. epidermidis and postulated to confer a selective survival advantage. 6
Additionally, we investigated the expression of phenol-soluble modulins (PSMs), which have been implicated in enhanced S. aureus and S. epidermidis virulence through leukocyte destruction.3,18 S. epidermidis produces several PSMs that share similarity with those found in S. aureus. 3 The β-type PSMs of S. epidermidis have been implicated in biofilm maturation and the dissemination of biofilm-associated infection, 19 whereas the frequently cytolytic α-type PSMs, such as δ-toxin, are commonly produced at lower levels in S. epidermidis compared with S. aureus. 3
We hypothesize that a combination of molecular determinants imparts a selective advantage, allowing certain strains of S. epidermidis to become important clinical pathogens, as typified by the following case. Using this acute, rapidly progressive early prosthetic hip infection with associated high-grade bacteremia as a representation of an invasive S. epidermidis infection, we sought to determine the combination of molecular characteristics of this isolate and further define associated genetic markers of this early invasive and persistent organism, which behaves like an aggressive Staphylococcus aureus. In addition to molecular typing methods, including SCCmec analysis, pulsed-field gel electrophoresis (PFGE), and multilocus sequence typing (MLST), we assessed the presence of adhesion factors and mobile elements bhp, icaA, IS256, IS257, and ACME as well as PSM production and the biofilm formation capacity of this organism.
Case report
A 60-year-old diabetic woman sustained a femoral neck fracture necessitating a left hip hemiarthroplasty. Two weeks postprocedure, she developed purulent drainage from the surgical site, requiring incision and drainage of the wound and treatment for 4 days with an unknown oral antibiotic, presumed to be a fluoroquinolone. She was subsequently hospitalized with fever, chills, increasing pain, erythema, and extensive drainage at the incision at 4 weeks after initial hemiarthroplasty.
On admission, she was febrile with a temperature of 38.2°C and appeared acutely ill. Purulent material was draining from the surgical wound, and erythema, warmth, and tenderness were present over the lateral hip. Laboratory tests revealed a leukocyte count of 14.6 × 109 cells/L, 83% neutrophils, and a serum blood glucose of 368 mg/dl, consistent with infection. Hip radiograph showed a well-fixed cemented hemiarthroplasty.
Extensive surgical debridement and modular component exchange were done. Copious purulent periprosthetic material noted intraoperatively was sent for microbiological culture. Four intraoperative tissue cultures and two sets of blood cultures (four of four bottles) grew methicillin-resistant S. epidermidis (MRSE). Intravenous vancomycin and oral rifampin were initiated after cultures were taken. After 8 days, therapy was switched to intravenous linezolid because of worsening renal insufficiency. A transthoracic echocardiogram showed no valvular abnormalities or vegetations, and the patient completed 6 weeks of oral linezolid as an outpatient. Five months after the debridement, she continued to have pain and underwent resection arthroplasty with placement of a temporary high-dose antibiotic–loaded cement spacer. Screening preoperative nasal and axillary cultures grew out methicillin-susceptible S. epidermidis and all four intraoperative tissue cultures grew out methicillin-resistant S. epidermidis, with the same antibiogram as the original isolates. The patient was subsequently treated with vancomycin, awaiting joint re-implantation.
Materials and Methods
Two sets of blood cultures from admission and four tissue cultures were collected intraoperatively at the time of initial incision and drainage, and four repeat intraoperative cultures were obtained during the revision surgery. As per routine, all microbiology cultures were processed according to Clinical and Laboratory Standards Institute (CLSI) procedures and MICs were determined by Vitek2 automated system (bioMérieux, Durham, NC).
Pulsed-field gel electrophoresis following an established protocol was performed on the four initial incision and drainage cultures, two blood cultures, four revision arthroplasty cultures, and the nasal and axillary swab cultures using SmaI. 13 These strains and the reference strains American Type Culture Collection (ATCC) 35984, that is, RP62A, and ATCC 12228 were analyzed by the MLST protocol as previously published. 16
Biofilm assay was performed according to previously published techniques in a 96-well microtiter plate with the following changes to the protocol. 10 Two blood culture isolates, eight intraoperative cultures, four from the first incision and drainage and four from the revision arthroplasty, and positive (RP62A) and negative controls (tripticase soy broth [TSB]) were grown overnight in a 37°C incubator on 5% Trypticase® Soy Agar with 5% sheep blood (BD, Sparks, MD). One loopful (107–10) was added to 100 μl of 900 ml TSB with 6 g yeast extract and 8 g glucose per liter and vortexed gently to suspend. Bacterial suspension was diluted 1:1000, and four duplicate 200 μl samples were added to a 96-well tissue culture–treated polystyrene plates (Corning Costar, Corning, NY) and incubated at 37°C overnight in air. To stain for biofilm production, nonadherent cells were washed in cold water by immersing the plate in a basin and tapping off loose cells. Two hundred microliters of crystal violet was added to each well for 60 seconds and washed by immersing in cold water 10 times. The crystal violet was solubilized with the addition of 100 μl ethanol for 10 minutes. After allowing to air dry for 30 minutes, the optical density was measured at OD595 nm (VersaMax; Molecular Devices, Sunnyvale, CA).
SCCmec analysis was performed using our recently developed multiplex molecular beacon assay that distinguishes SCCmec types I–IV.
2
Multiplex PCR for ACME (arcA and opp3) and Panton-Valentine leukocidin (PVL) was performed in 30 μl reaction mixtures with Denville Taq® reaction mix. DNA amplification was performed on a Biogene automated thermocycler using the following conditions: 94°C for 2 minutes, followed by 35 cycles of 94°C for 30 seconds, 55°C for 60 seconds, and 72°C for 90 seconds, and a final elongation step at 72°C for 5 minutes. Primer sequences used are as follows:
Conventional PCR for bhp, IS256, IS257, and 16s RNA for internal positive controls was performed using previously published primer sequences. 8 Conventional PCR and southern blotting for ica were completed according to previously published methods. 11 All clinical isolates were compared with biofilm-producing reference strain RP62A (ATCC 35984) and non–biofilm-producing strain ATCC 12228 (Manassas, VA). The PCR for aap was performed according to previously published literature. 12
Cultures in which PSM production has to be determined were inoculated from precultures grown overnight at 1:100 and grown to stationary growth phase (8 hours). One hundred microliters of culture filtrates were used to measure PSMs by reversed-phase high-pressure chromatography/electrospray ion mass spectrometry (HPLC/MS) on an Agilent 1100 system coupled to a Trap SL mass spectrometer using a water/acetonitrile gradient, as previously described. 20 PSM production was quantified by calculating extracted ion chromatograms for the major m/z peak of each PSM, using Agilent Quant Analysis software. PSM production in strain ATCC12228 was set to 100%. 3 Measurements were performed using three independent cultures for each strain.
Results
The PFGE pattern demonstrated that the S. epidermidis from intraoperative hip cultures, both from the initial surgery and the revision arthroplasty, were identical to the two blood isolates using SmaI restriction enzymes, but different from the axillary and nasal isolates as well as the reference strain (Fig. 1). Clinical isolates from the hip and bloodstream were susceptible to rifampin (MIC <1), tetracycline (MIC <4), trimethoprim/sulfamethoxazole (MIC <0.5/9.5), linezolid (MIC = 2), daptomycin (MIC <0.5), and vancomycin (MIC <2 mg/L), resistant to nafcillin (MIC >2), amoxicillin/clavulanate (MIC >4/2), cefazolin (MIC >8), and macrolides (MIC >4), and intermediate to levofloxacin (MIC = 4).

PFGE of Staphylococcus Epidermidis. Lane designation: L: ladder; 1: RP62A; 2: blood isolate; 3: blood isolate; 4–7: tissue isolate from first surgery; 8–11: tissue isolate from second surgery; 12: nares; 13: axilla.
MLST sequencing identified clinical isolates as sequence type (ST) 5, a double locus variant of the ST10 biofilm-producing RP62A, whereas reference strain ATCC 12228 belonged to ST8 (Fig. 2). SCCmec typing showed that the clinical isolates contained SCCmec IV, compared with RP62A, which contains SCCmec II. The PCR results demonstrated that the invasive clinical isolates carried ACME (arcA and opp3), but not PVL. Conventional PCR for virulence and persistence factors demonstrated that the clinical isolates and RP62A control possess biofilm-associated homologous protein (bhp) and IS257. IS256 and ica were present in the biofilm producer RP62A. Interestingly, our clinical strains, although possessing the bhp locus, were not biofilm positive in our assay (data not shown). Aap was present in RP62A, ATCC 12228, all blood and introperative cultures, and the axilla screen, but was negative in the nasal screen S. epidermidis isolate (PCR not shown). The clinical isolates showed an overall stronger production of PSMs when compared with the biofilm-producing strain RP62A and nonbiofilm producer ATCC 12228, as shown for the most strongly produced δ-toxin and β-type PSMs (Fig. 3). Compared with the nonbiofilm producer ATCC 12228, the biofilm producer RP62A demonstrated a reduced delta toxin (PSMγ) production (41.9%) as well as a reduction in PSM-β1 (41.0%) and PSM-β2 (22.0%) toxins. The clinical isolates (two blood cultures and primary incision and drainage cultures) demonstrated an increased production of δ-toxin, PSM-β1, and PSM-β2 production when compared with RP62A, (Table 1) whereas the production compared with ATCC 12228 was significant only for some isolates and when values for all isolates were averaged (Fig. 3).
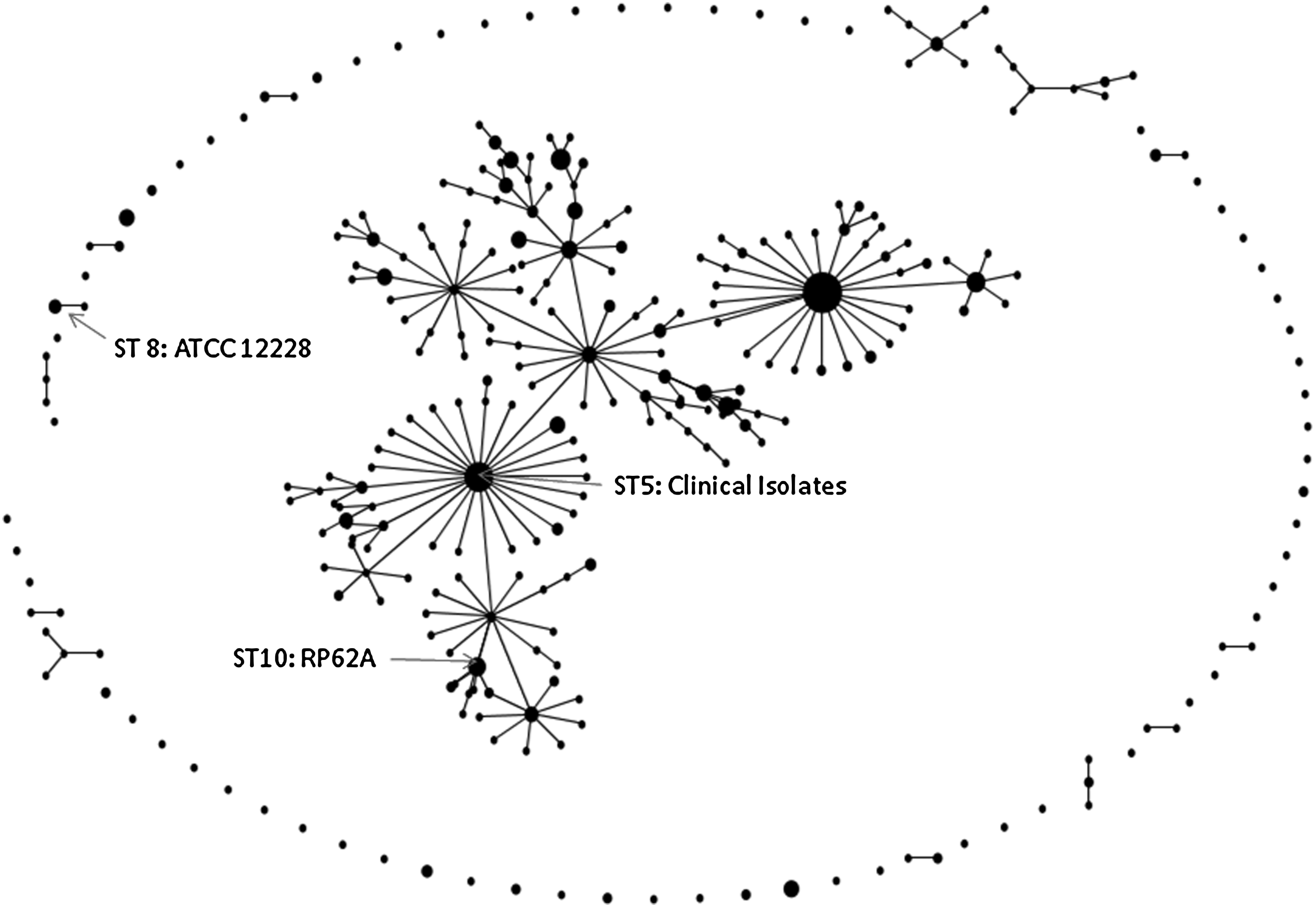
S. epidermidis eBURST (based on repeated sequence types) demonstrating relatedness between ATCC 12228, RP62A, and clinical isolates (accessed February 9, 2011; http://sepidermidis.mlst.net/eburst/database.asp).
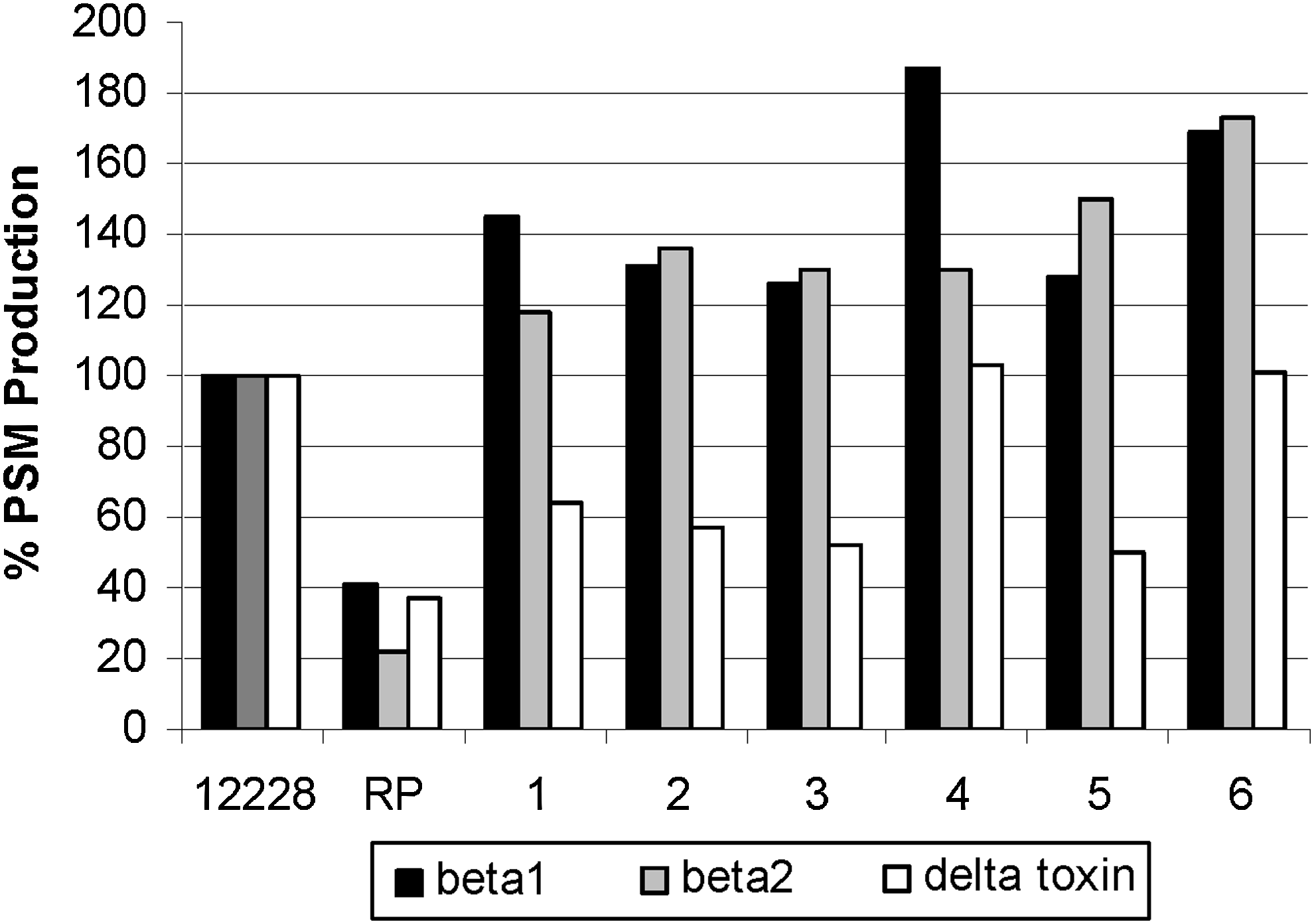
Phenol-soluble modulin (PSM) production of RP62A and clinical isolates (1–2: blood isolates; 3–6: tissue isolates) versus ATCC 12228, standardized to 100%; all were statistically significant (p < 0.05).
Production is shown relative to the strain ATCC 12228. Statistical evaluation was performed on raw data (clinical samples 1–2: blood isolates; 3–6: tissue isolates) as follows: Single clinical isolates were compared with RP62A or ATCC 12228, respectively, by one-way ANOVA and Dunnett's posttest. Averaged data for clinical isolates were compared with RP62A or ATCC 12228 by one-way ANOVA with Bonferroni posttest.
p < 0.05; **p < 0.01; ***p < 0.001.
ns, non-significant.
Discussion
Adhesion, controlled by polysaccharide intercellular adhesin (PIA), is encoded by the icaADBC locus and associated with insertion sequence (IS) 256 through phase variation of biofilm formation, which alters the expression of ica. 11 This insertion sequence was reported to be a more sensitive molecular marker of invasiveness than ica; however, conflicting evidence has been presented as to the role of ica in S. epidermidis.8,11 In strains lacking the icaADBC locus, biofilm formation is mediated by the biofilm-associated protein or biofilm homologous protein (bap, bhp) or the accumulation-associated protein (aap) 15 Although our clinical infecting strain contained bhp and aap, it was not associated with a biofilm-positive phenotype by standard crystal violet assay, raising the speculation that the expression of the biofilm homologous protein is downregulated, favoring detachment and dissemination, consistent with the clinical presentation of our device infection associated with a high-grade bacteremia. The ACME gene cluster, generally associated with MRSA USA300, was identified on the chromosome of S. epidermidis ATCC 12228, but not in biofilm-producing RP62A (ATCC 35984).6,7 Our non–biofilm-producing clinical isolates, although evolutionarily more closely related to RP62A, contain both arcA and opp3A genes, suggesting that ACME may act as a key virulence factor in non–biofilm-producing S. epidermidis as well.
PSMs are a group of pro-inflammatory and cytolytic peptides with an additional function in biofilm development.19,21 Absence of PSMs is believed to result in more pronounced biofilm formation, likely owing to abolished biofilm detachment, which in S. epidermidis is facilitated by β-type PSMs. In contrast particularly to the biofilm former RP62A, the clinical strains showed pronounced production of both cytolytic α-type and biofilm-active β-type PSMs, in keeping with our in vitro and clinical observations of low biofilm-forming capacity and strong bacteremia caused by those strains.
The clinical presentation of an early PJI described herein, caused by a phenotypically non–biofilm-producing methicillin-resistant ST5 S. epidermidis, is evolutionarily related to the classic biofilm producer RP62A. Our ACME arcA- and opp3A-containing clinical strain caused a clinical syndrome comparable to infections commonly encountered with invasive USA300 methicillin-resistant S. aureus. We hypothesize that this combination of virulence determinants, resistance factors, and the appropriate clinical setting allowed for attachment, invasion, and proliferation of an otherwise commensal organism. Although S. epidermidis is frequently discounted as an invasive pathogen, our representative case report and molecular characterization demonstrate that with acquisition of mobile genetic elements and under appropriate conditions, S. epidermidis can become aggressive early in a prosthetic joint infection, even in the absence of biofilm formation. This enhanced bacterial virulence and persistence superimposed on a background of antimicrobial resistance and medication intolerance not only limited anti-infective treatment options, but also initiated an infection difficult to eradicate with invasive intervention. As seen with this clinical example, a heightened clinical suspicion and aggressive treatment course may be warranted when encountering an otherwise “low-virulence” organism.
Footnotes
Acknowledgment
M.E.R. was supported by Ruth Kirschstein T-32 training grant (T32-AI-7329-16).
Disclosure Statement
No competing financial interests exist.