
Abstract
A total of 59 and 74 nonduplicate plasmid-mediated quinolone resistance (PMQR) genes-carrying Klebsiella pneumoniae and Escherichia coli isolates were collected. All strains were assayed for fluoroquinolone susceptibility and the prevalence of quinolone resistance-determining regions (QRDRs) mutation. The association between PMQR determinants and common β-lactamase genes was also analyzed. Genetic relatedness of the isolates was analyzed by pulsed-field gel electrophoresis (PFGE). The PMQR genes-carrying K. pneumoniae and E. coli isolates exhibited high fluoroquinolone resistance rates, indicating that PMQR determinants play an essential role in the development of fluoroquinolone resistance. Remarkably, most qnr-carrying strains had only a single or no QRDR mutation in GyrA or ParC, and most exhibited decreased ciprofloxacin (CIP) susceptibility or low-level CIP resistance. However, 71.4% and 98.4% of qnr-negative K. pneumoniae and E. coli contained double QRDR mutations, and most presented high-level CIP resistance. Additionally, K. pneumoniae presented a lower CIP resistance rate than E. coli (59.3% vs. 91.9%) and low carriage of double QRDR mutations (38.9% vs. 89.9%). Also, most (88.1%) K. pneumoniae examined in this study carried qnr and only 14.9% of E. coli were qnr positive. Thus, the high fluoroquinolone susceptibility of K. pneumoniae isolates is primarily due to fewer QRDR substitutions as a result of high prevalence of qnr alleles in the host. Our findings support the hypothesis that chromosomal resistance mutations could be affected by the presence of Qnr, in other words, Qnr may protect the QRDR domains in the gyrase and topoisomerase IV from mutations under the inhibition of fluoroquinolones. Another remarkable finding was that the PMQR genes-carrying K. pneumoniae exhibited much higher proportions of extended-spectrum β-lactamases (ESBLs)-positive phenotype than E. coli (73.5% vs. 59.5%). This is due to not only the high prevalence of SHV-type ESBL-conferring enzymes in K. pneumoniae but also the interference of DHA-producing K. pneumoniae as a result of the strong association between qnrB and blaDHA.
Introduction
To date, three PMQR mechanisms have been described: Qnr, AAC(6′)-Ib-cr, and QepA, which mediate quinolone resistance-determining regions (QRDRs) protection, drug modification, and drug efflux, respectively. The prevalence of PMQR genes has been increasingly reported among common Enterobacteriaceae and strong association of PMQR genes with other resistance determinants has also been documented.2,22 By increasing the minimum inhibitory concentration (MIC), PMQR mechanisms may become complementary or enhanced elements for fluoroquinolone resistance caused by drug target alterations within QRDRs. By increasing the mutant prevention concentration, PMQR determinants may also lead to target substitutions within QRDRs and appearance of high-level fluoroquinolone resistance phenotype as a result of widening the mutant selection window.21,22 However, PMQR determinants confer only low-level resistance or decreased fluoroquinolones susceptibility and do not affect fluoroquinolones bactericidal activity against PMQR gene-carrying isolates. 13 In addition, by lowering gyrase and topoisomerase IV binding to DNA, Qnr may reduce the amount of enzyme–DNA complex, which serves as the target of fluoroquinolones23,24 and protect QRDRs domain from mutation under the inhibition of fluoroquinolones. 4 These conflicting results limit the understanding about the biological functions of PMQR determinants, particularly Qnr.
In this study, the prevalence of common β-lactamases genes and QRDRs alterations among clinical PMQR determinant-carrying K. pneumoniae and E. coli isolates were investigated. In addition to the prevalence of diverse resistance determinants, antimicrobial resistance phenotypes present significant differences between the two species. The relationship between resistance genotype and phenotypic characterizations was also discussed.
Materials and Methods
Bacterial strains
In total, 112 nonduplicated K. pneumoniae strains and 579 E. coli clinical isolates with a ciprofloxacin (CIP) MIC ≥0.25 μg/mL were collected from nine tertiary hospitals in seven cities across China in 2008. All isolates were screened for PMQR determinants qnrA, qnrB, qnrS, aac(6′)-Ib-cr, and qepA. Fifty-nine K. pneumoniae and 74 E. coli isolates were positive for at least one PMQR gene and were analyzed in this study. E. coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as control strains for antimicrobial susceptibility testing and ESBL screening, respectively.
Antimicrobial susceptibility and ESBL phenotype detection
MICs to CIP, levofloxacin, amikacin, cefotaxime, ceftazidime, cefepime, imipenem, and meropenem (National Institutes for Food and Drug Control, Beijing, China) were measured using the agar dilution method. Susceptibility results were interpreted according to the Clinical Laboratory Standards Institute (CLSI) interpretive standards. ESBL phenotype was detected by the disk diffusion method of screening and confirmatory tests for ESBL. 5
Resistance genes investigation
Template DNA was extracted using the boiling method. qnrA, qnrB, qnrS, qepA, blaTEM, blaSHV, blaCTX-M, blaPER, blaVEB, blaGES, and blaOXA and plasmid-mediated blaAmpC were detected by polymerase chain reaction amplification (ABI Veriti Thermal Cycler; Applied Biosystems Asia Pte. Ltd., Singapore). aac(6′)-Ib-cr was identified using a previously described method. 16 Specific primer sets have been previously described.3,6
QRDRs alteration detection
QRDRs of gyrA and parC were amplified by polymerase chain reaction as previously described. 7 The amplicons were sequenced by BGI Life Tech Co., Ltd., Beijing, China. Predicted amino acid sequences of GyrA and ParC were analyzed for amino acid alterations by comparison with wild-type GyrA (GenBank accession number: NP_416734) and ParC (GenBank accession number: NP_417491) of E. coli K-12 (GenBank accession number: NC_000913).
Pulsed-field gel electrophoresis
DNA fingerprinting profiles were analyzed by PFGE resulting from restriction enzyme digestion with XbaI (New England Biolabs, Beijing, China) according to procedures developed by the United States Center for Disease Control and Prevention PulseNet program. 17 Interpretation of PFGE patterns was performed using BioNumerics software (Applied Maths, St-Martens-Latern, Belgium) using the dice similarity coefficient. Clusters were defined as DNA patterns sharing ≥70% similarity. Similarity differing by <5% was considered as representative of subtypes within the main group.
Statistical analysis
Univariate comparisons were conducted using the χ2 test and a p-value of ≤0.05 was considered statistically significant.
Results
Antimicrobial susceptibility
The resistance rate to commonly used antibiotics is listed in Table 1. Approximately 63.6% of qnr– and aac(6′)-Ib-cr–carrying K. pneumoniae isolates were resistant to amikacin, and 59.3% and 91.9% of PMQR-positive K. pneumoniae and E. coli exhibited CIP resistance, respectively (Table 1). Approximately 28.8% of K. pneumoniae and 28.9% of E. coli isolates were multidrug-resistant strains that simultaneously exhibited resistance to cefotaxime, CIP, and amikacin.
ESBL, extended-spectrum β-lactamase; QRDR, quinolone resistance-determining region.
Distribution of PMQR determinants
In this study, a significant difference in PMQR determinants prevalence was identified between clinical K. pneumoniae and E. coli isolates. qnr was the predominant PMQR gene in K. pneumoniae and was rarely detected in E. coli. Among 59 PMQR gene-carrying K. pneumoniae, qnr was detected in 52 isolates (88.1%). qnrB was the most common qnr allele (32/52, 61.5%), followed by qnrS (18/52, 34.6%). qepA was rarely observed in K. pneumoniae. Among 74 E. coli isolates, qnr, aac(6′)-Ib-cr, and qepA were detected in 11 (14.7%), 41 (55.4%), and 28 (37.8%) isolates, respectively (Fig. 1).
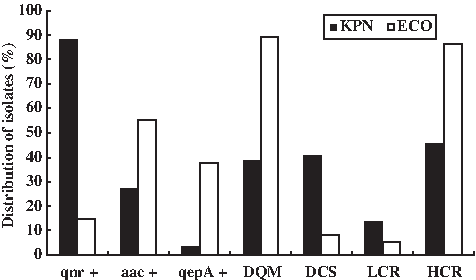
Distribution of clinical plasmid-mediated quinolone resistance-positive KPN and ECO. aac, aac(6′)-Ib-cr; DQM, double quinolone resistance-determining region mutations; DCS, decreased ciprofloxacin; LCR, low-level ciprofloxacin resistance; HCR, high-level ciprofloxacin resistance; KPN, Klebsiella pneumoniae; ECO, Escherichia coli.
QRDRs substitution and CIP susceptibility
E. coli had more double QRDR mutations and exhibited higher fluoroquinolone resistance than K. pneumoniae (Table 1 and Fig. 1). Remarkably, approximately two-third of qnr-carrying isolates have no QRDR substitutions or single mutation in GyrA or ParC, and most strains exhibited decreased CIP susceptibility (MIC = 0.5–2 μg/ml) or low-level CIP resistance (MIC = 4–8 μg/ml) (Table 2). However, most qnr-negative isolates had double QRDR substitutions and exhibited high-level CIP resistance (MIC ≥16 μg/ml) (Table 2).
CIP, ciprofloxacin.
Prevalence of common β-lactamase genes
A significant difference in blaSHV and blaDHA prevalence was identified between PMQR genes-carrying K. pneumoniae and E. coli (Table 1). Twenty-one of 59 (35.6%) K. pneumoniae carried blaDHA but no blaCTX-M, whereas 6 of 17 (35.3%) MDR K. pneumoniae produced SHV-type ESBLs without CTX-M. All of 32 qnrB-positive K. pneumoniae strains carried blaDHA.
Genetic relatedness of PMQR genes-carrying isolates
Forty-eight PFGE subtypes and 9 clusters were identified in 59 PMQR genes-positive K. pneumoniae isolates. Five PFGE subtypes were found in more than one isolate, and all of these strains were isolated from the same city. Among 74 E. coli isolates, 68 PFGE subtypes and 19 clusters were identified, and 5 PFGE subtypes were found in more than 1 isolate.
Discussion
PMQR determinants always coexist with a variety of resistance genes on the same plasmid, such as blaCTX-M, plasmid-mediated blaAmpC, and 16S rRNA methylases genes, and transfer simultaneously cross-species/genera.2,22 The isolates carrying these resistance determinants could be selected out under selective pressure caused by widespread use of fluoroquinolones in human and veterinary medicine. Although low-level fluoroquinolone resistance or decreased fluoroquinolone susceptibility has been identified in PMQR genes-carrying isolates, the impact of these genotypes was similar to that of QRDRs mutations. 1 Therefore, the study on prevalence of PMQR determinants and coexistence with other resistance genes will contribute to understand the characterization of these resistance determinants. In this study, high prevalence of CIP resistance suggests that PMQR determinants play an essential role in the promotion of fluoroquinolones resistance. Although strains with no QRDR substitutions generally exhibited decreased fluoroquinolone susceptibility or low-level fluoroquinolone resistance, these isolates can sufficiently reduce the bactericidal activity of fluoroquinolones, demonstrating obvious clinical significance of PMQR determinants. 1
Previous studies have demonstrated that the primary fluoroquinolone resistance mechanism in clinical E. coli isolates is the QRDR mutations in GyrA and ParC. 14 QRDR alterations act as the key factor of fluoroquinolone resistance in clinical isolates. In this study, E. coli exhibited much higher fluoroquinolones resistance rates and increased occurrence of QRDR substitutions than K. pneumoniae (Table 1). qnr-carrying isolates exhibited fewer QRDRs mutations, along with higher CIP susceptibility. Conversely, qnr-negative isolates had more QRDRs mutations and frequently exhibited high-level CIP resistance. Therefore, the high fluoroquinolone susceptibility exhibited by clinical K. pneumoniae isolates might be due to fewer QRDR substitutions resulting from a high host prevalence of qnr alleles. Similar results have also been reported in other Enterobacteriaceae, such as Enterobacter cloacae. 10 In vitro analysis has confirmed that Qnr protects QRDR domains in gyrase and the topoisomerase IV from mutation under fluoroquinolones inhibition by reducing the amount of the enzyme–DNA complex, 4 which are the fluoroquinolones target formed by the enzyme–DNA binding.23,24 Thus, the observation that fewer QRDR alterations occurred in qnr-carrying isolates supports the hypothesis that chromosomal resistance mutations are affected by the presence of Qnr under the fluoroquinolones selective pressure. 22 Qnr may prevent the mutations in gyrase and topoisomerase IV through QRDRs protection. On the one hand, Qnr supplements QRDR mutations-mediated fluoroquinolone resistance and leads to the appearance of high-level fluoroquinolone resistance strains by widening the mutant selection window.21,22 Further analysis is required for a better understanding of the underlying mechanisms causing the conflict.
Clinical ESBLs-producing isolates exhibited significant geographical diversity. 9 In this study, 59.5% of E. coli isolates presented the ESBL-positive phenotype, which is similar to the average ratio of 61% previously recorded in the same area. 25 However, PMQR genes-carrying K. pneumoniae strains exhibited much higher proportion of ESBLs-positive phenotype than the average (73.5% vs. 36.6%). It was reported that few TEM-type ESBL variants have been identified in Enterobacteriaceae and one-fifth of ESBL phenotypes were ascribed to SHV variants. 11 In this study, SHV-type ESBL was the only ESBL genotype among 35.3% of MDR K. pneumoniae isolates analyzed. Thus, the increased ESBL prevalence among K. pneumoniae is primarily conferred by SHV. Because the SHV-type enzymes exhibit stronger hydrolytic activity for ceftazidime than cefotaxime, 20 the high prevalence of SHV-type ESBL also contributes to increased ceftazidime resistance in K. pneumoniae (Table 1). Another remarkable observation in this study was the strong association between qnrB and blaDHA. All (32 of 32) qnrB-positive K. pneumoniae carried blaDHA, and similar results have been previously described. 15 Plasmid-mediated AmpC enzymes not only hydrolyze cephalosporins, such as cefotaxime and ceftazidime, but also affect ESBL phenotype detection and cause an unreliable result. 18
Prior fluoroquinolones use has been shown to be an independent risk factor for infections caused by ESBL-producing E coli 26 and carbapenem-resistant K. pneumoniae. 8 The combination of PMQR genes with the determinants, such as blaCTX-M and blaSHV, may contribute to the appearance of MDR isolates. This may be attributed to the selective pressure caused by fluoroquinolones as driving force to select for strains carrying not only PMQR determinants but also ESBLs genes. Additionally, previous use of antimicrobials such as fluoroquinolones may select for resistance to other antibiotics such as cephalosporins or carbapenems. 19 Thus, reducing use of antibiotics that facilitate selective pressure will contribute much to the control of infections caused by ESBLs- or carbapenemases-producing isolates.
Footnotes
Disclosure Statement
No competing financial interests exist.