
Abstract
Fluoroquinolones (FQs) have been increasingly used for effective treatment of infections caused by rapidly growing mycobacteria, and resistance to this drug has been predominantly attributed to gyrA and gyrB mutations. Accordingly, this study investigated a total of 36 Mycobacterium massiliense clinical isolates for their susceptibility to ciprofloxacin and presence of gyrA and gyrB gene mutations. The minimal inhibitory concentration (MIC) values, determined by broth microdilution method, of 35 ciprofloxacin-resistant isolates ranged between 4 and 16 μg/mL and a single susceptible isolate was obtained. A total of 31 of 35 (88.5%) ciprofloxacin-resistant isolates presented an amino acid substitution at codon 90 (Ala-90→Val) and no isolate presented mutation at position Asp-94. Moreover, 4 of 35 (11.4%) ciprofloxacin-resistant and one susceptible isolate had no mutation in Ala-90 and Asp-94. No gyrB mutation was observed in all tested M. massiliense isolates. In conclusion, our results have shown that mutations of gyrA codon 90 are frequent and may constitute an important mechanism of resistance to FQ in M. massiliense.
Introduction
Infections caused by RGM are frequently difficult to treat and require specific regimens that include several drugs given over a prolonged period; relatively few drugs are effective against mycobacteria. 2 These organisms are resistant to the conventional antituberculous agents and their susceptibilities to other antimycobacterial agents differ among species.6,37 The increasing use of fluoroquinolones (FQs) in mycobacterial infections has led to emergence of FQ-resistant isolates.10,30 FQs inhibit the bacterial type II topoisomerase DNA gyrase, an ATP-dependent enzyme essential for DNA supercoiling, which is required for DNA replication and gene transcription. This enzyme consists of two proteins, GyrA and GyrB, which are encoded, respectively, by gyrA and gyrB genes. 9 Mutations occurring in a short sequence named the quinolone resistance-determining regions (QRDRs) in gyrA and gyrB genes have been generally involved in resistance to quinolones.14,28,38 Mutations of gyrA gene, mainly at position 90 and 94 (Mycobacterium tuberculosis numbering system), have been found to confer high-level FQ resistance in several bacterial species.3,10,20,38 In contrast, gyrB mutations are frequently clustered at codons 495, 516, and 533; alterations in this gene were rarely observed in mycobacterial clinical isolates. 26
Because of the increasing clinical importance of surgical infections by RGM worldwide, the aim of the present study was to investigate the gyrA and gyrB gene mutation in 35 ciprofloxacin-resistant M. massiliense isolated from Southern Brazil.
Materials and Methods
Bacterial isolates
A total of 36 isolates from patients (one isolate per patient) submitted to surgical procedures at hospitals located in the city of Curitiba, Southern Brazil, from August 2007 to January 2008, were obtained for this study. Procedures were performed in four private hospitals: 28 of 36 patients from hospital 1, 6 of 36 from hospital 2, 1 of 36 from hospital 3, and 1 of 36 from hospital 4. Samples were collected by either biopsy or aspiration of abscess fluids from patients presenting signs of infection clinically suggestive of RGM, such as cutaneous wound, presence of abscess, and delayed wound healing. Samples were cultivated on Lowenstein-Jensen solid medium for up to 4 weeks at 37°C. 31 All M. massiliense were submitted to genotypic analysis by pulsed-field gel electrophoresis (PFGE) (Fig. 1) and compared with M. massiliense BRA 100 clone from an outbreak that occurred in Rio de Janeiro state (CRM 0020). 12 PFGE analysis revealed that all isolates presented indistinguishable patterns, and according to the criteria proposed, 34 the isolates were considered to belong to the same strain from an epidemic of surgical infections that occurred in seven geographically distant states in Brazil. 25 One isolate of Mycobacterium abscessus, Mycobacterium chelonae, and Mycobacterium fortuitum collected at hospital 1 during the same period were included in this study. This study was approved by the Internal Review Board Hospital do Trabalhador Ethics Committee under the number 244690.
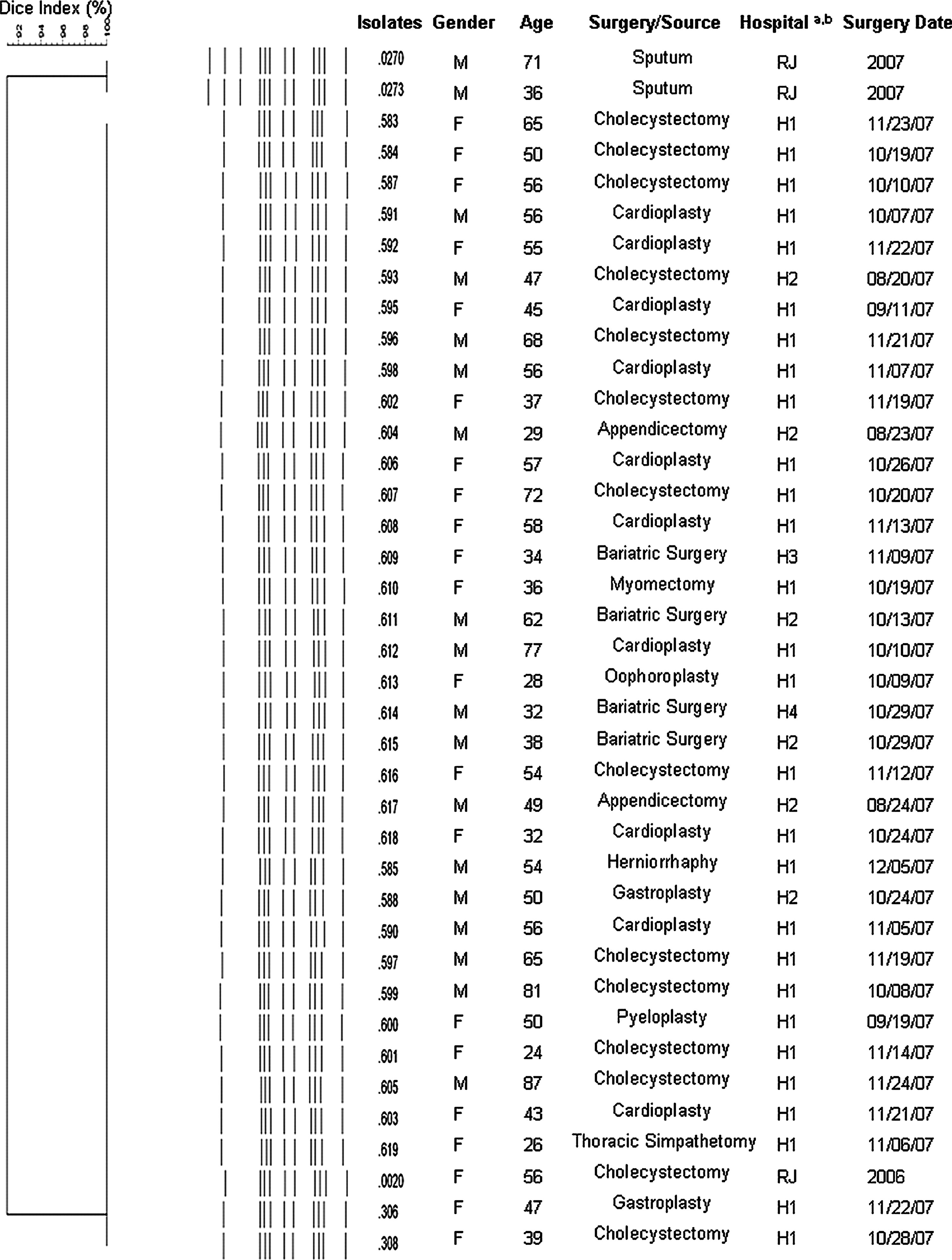
Dendrogram of PFGE patterns of Mycobacterium massiliense isolates from Curitiba outbreak. Identical patterns clustered the isolates from surgical patients. Comparison was performed with a molecular analysis fingerprinting GelCompar software (Applied Maths). The Dice coefficient was used for calculating the percentages of similarity among the profiles. The PFGE patterns of M. massiliense isolated from Rio de Janeiro outbreak (CRM 0020) and two epidemiologically unrelated strains from Rio de Janeiro patients (CRM 0270 and CRM 0273) were included in the analysis. aHn, Mycobacterial isolate obtained from patient submitted to surgery at hospital number n; bRJ, mycobacterial isolate recovered from Rio de Janeiro City. PFGE, pulsed-field gel electrophoresis.
Species identification
Species identification was based on partial sequencing of the rpoB gene. Extraction of DNA from clinical isolates was carried out using a commercially available kit (Nuclisens Basic Nasba Diagnostics; bioMérieux) based on a published method. 5 DNA amplification and sequencing of polymerase chain reaction (PCR) products was performed with primers MycoF (5′-GGCAAGGTCACCCCGAAGGG-3′) and MycoR (5′-AGCGGCTGCTGGGTGATCATC-3′) as previously reported. 1 The resulting sequences were aligned with a free software (BioEdit, version 7.0.5.3) 17 using M. tuberculosis H37Rv (GenBank accession number: BX842574.1) as the reference sequence.
Antimicrobial susceptibility test
All isolates were evaluated for their susceptibility to ciprofloxacin. Minimal inhibitory concentration (MIC) values were determined by broth microdilution method and interpreted according to the tentative guidelines established by the National Committee for Clinical Laboratory Standards (NCCLS). 27 Bacterial suspensions were prepared by sweeping the confluent portion of growth and adjusted to a final inoculum (5 × 105 colony-forming units [CFU]/mL) in Mueller-Hinton broth (Difco) with 0.02% Tween 80 (Difco). The medium used in microdilution was cation-supplemented Mueller-Hinton broth without Tween 80. The final organism concentration was 1 × 104 to 5 × 104 CFU per tube. Serial double dilutions of antimicrobial agents were prepared, and their concentrations in the tubes followed the NCCLS recommendation. The inoculated tubes were incubated at 30°C in ambient air and read after 72 hr. The MIC was defined as the lowest concentration of the drug that inhibited visible growth. Staphylococcus aureus ATCC 29213 was used as a control strain.
Fisher's exact test was used to correlate the MIC values with the four hospitals involved. The level of significance was established at a p-value of 0.05.
gyrA and gyrB PCR sequencing
DNA was obtained from a loopful of culture resuspended in 200 μL of Tris-EDTA (TE) buffer (10 mM Tris-HCl and 1 mM ethylenediaminetetraacetic acid [pH 8.0]), heat inactivated at 100°C for 10 min, and centrifuged at 10,000 g at 4°C for 20 min. Amplification of the DNA fragments corresponding to the gyrA and gyrB QRDRs were performed as previously described using the degenerated oligonucleotides Pri9 (5′-CGCCGCGTGCTG/CATGCA/GATG-3′) and Pri8 (5′-C/TGGTGGA/GTCA/GTTA/GCCC/TGGCGA-3′) for gyrA 15 and using GyrbA 11 (5′-GAGTTGGTGCGGCGTAAGAGC-3′) and GyrbE 16 (5′-CGGCCATCAA/GCACGATCTTG-3′) for gyrB. The amplification reactions consisted of the following steps: 1 denaturation cycle at 94°C for 10 min and 40 cycles of amplification at 94°C for 1 min, 55°C for 1 min, and 72°C for 1 min, followed by 1 elongation cycle at 72°C for 10 min. The sizes of gyrA and gyrB amplified fragments were 216 and 268 bp, respectively. The nucleotide sequences of gyrA and gyrB PCR products were independently determined on both strands and performed as previously described. 16
Nucleotide sequence accession number
The nucleotide sequences reported here are available at GenBank database under accession numbers JF966195, JF966196, JF439303, and JF439304.
Results
Analysis of the rpoB gene sequences of M. massiliense isolates gave identical results and showed 100% similarity (695/695) to the sequence of M. massiliense strain INCQS 594 (GenBank accession number: EU117207). M. abscessus sequence was similar (695/695) to M. abscessus type strain P02 (GenBank accession number: FJ590436.1); M. chelonae was identical (695/695) to M. chelonae ATCC 19237 type strain, retrieved from GenBank under the accession number AY262740.1; and M. fortuitum presented 100% similarity (695/695) to M. fortuitum type strain CIP 104534T (GenBank accession number: AY147165.1).
Thirty-five M. massiliense isolates (35/36, 97.2%) were ciprofloxacin resistant and a single isolate was characterized as susceptible. Table 1 shows the MIC values obtained and the drug resistance profiles correlated with mutations in gyrA. There was no significant difference between MIC values of M. massiliense and the hospitals where the samples were collected (p > 0.05). Isolates of M. abscessus (MIC = 16 μg/mL) and M. chelonae (MIC = 16 μg/mL) were resistant to ciprofloxacin, whereas M. fortuitum was susceptible (MIC <0.125 μg/mL).
QRDR, quinolone resistance-determining region; R, resistant; MIC, minimal inhibitory concentration.
Peptide sequences of GyrA and GyrB (amino acids 74–113 and 495–533, respectively) were deduced from the nucleotide sequences. An overall 31/35 (88.6%) ciprofloxacin-resistant isolates presented an amino acid substitution at codon 90 (Ala-90→Val) and no isolate presented mutation at position Asp-94. However, 4/35 (11.4%) ciprofloxacin-resistant isolates had no mutation in Ala-90 and Asp-94 (Table 1). Ciprofloxacin-susceptible M. massiliense presented only an amino acid substitution at position Thr-80→Ala, which was not observed in resistant isolates.
Ciprofloxacin-resistant M. abscessus had an Asp-94→Gly substitution and M. chelonae presented two mutations, Ala-90→Val and Asp-94→Asn; no mutation was found in GyrA peptide sequences of M. fortuitum susceptible to ciprofloxacin.
No mutation was detected in gyrB gene of M. abscessus, M. chelonae, M. fortuitum, and all M. massiliense studied.
Discussion
GyrA QRDR sequences from resistant mycobacterial species have an alanine residue at position 90 (Ala-90) in the A subunit of DNA gyrase, whereas susceptible species have a serine. 16 In our study, a total of 4 of 35 (11.4%) ciprofloxacin-resistant isolates presented an Ala-90 in gyrA; and these results are in agreement with that previously reported. 16 This resistance to quinolones could be due partly to the lack, at this position, of the hydroxyl function carried by the side chain of serine, which is replaced by the methyl group of alanine. The presence of an alanine at this position could decrease the interaction between the gyrase A complex and the quinolone, because the amino acid at position 90 is the most common residue involved in quinolone resistance in M. tuberculosis and other bacterial species.3,7,20,33,38
Resistance to FQ in Mycobacterium species has been predominantly attributed to gyrA mutations, which are frequently clustered at codons 90 and 94.26,29 We found a high mutation frequency (88.6%) in gyrA gene at position 90 in 31 ciprofloxacin-resistant M. massiliense, and all mutated isolates carried an Ala-90→Val, the same substitution reported in M. tuberculosis and M. smegmatis strains.10,30 This amino acid substitution does not change the function of the side chain but only alters the bulk at this location. Thus, the steric hindrance related to the valine residue could explain the resistance found in isolates presenting Ala-90→Val mutation. M. chelonae showed the same substitution (Ala-90→Val) found in M. massiliense and presented a high-level resistance to ciprofloxacin.
Point mutation (GCG→GTG) in our isolates presenting Ala-90→Val substitution did not create a new restriction site on PFGE analysis, so the pattern was the same found in BRA100 strain; however, this strain does not present an Ala-90→Val substitution. Thus, we presume that in this outbreak there is a single strain presenting genetic diversity (Val-90 isolates and Ala-90 isolates) that was spread throughout the country; the high frequency of an Ala-90→Val substitution found in our isolates could be a characteristic mutation present in strains from Southern Brazil.
Ciprofloxacin-susceptible M. massiliense carried a Thr-80→Ala substitution in gyrA, which has been previously reported in M. tuberculosis isolates. 36 This substitution at position 80 seems not to play a relevant role in resistance to quinolones and was not observed in resistant isolates. So, according to hypothesis previously raised, 12 we suppose that some isolates, as ciprofloxacin-susceptible isolate found here, has been already in environment and were not brought from other states; the selective pressure before the outbreak may be responsible for some mutations (Thr-80→Ala), but was not sufficient to change its susceptibility profile.
It is interesting to note that gyrA mutations in codon 94 were not observed in all M. massiliense isolates; the absence of gyrA mutation at this position is not in accordance with previous reports in M. tuberculosis, which have described a relatively equivalent frequency of such mutation when compared with codon 90.36,39 In contrast, mutations in codon 94 were found in ciprofloxacin-resistant M. abscessus (Asp-94→Gly) and M. chelonae isolates (Asp-94→Asn). The same mutation pattern was previously found in M. tuberculosis and the authors observed that different substitutions of amino acid 94 resulted in different levels of drug resistance. 39 This occurrence could be due to the differences of amino acid in chemical properties determining the binding of FQ and DNA gyrase. However, in our study, M. abscessus and M. chelonae showed similar MIC values, despite different mutations in codon 94.
In this study, all samples were collected for microbial culture before initial antibiotic treatment of patients and none of the patients has received quinolones for at least 4 weeks before the surgical procedures. Thus, we presume that the incidence of ciprofloxacin-resistant M. massiliense may be due to selective pressure caused by drug abuse before the occurrence of the present cases. Additionally, a possible explanation for the presence of different MIC values obtained in our isolates is that, as in S. aureus, the development of high-level resistance in quinolones occurs during therapy in sequential steps, with a first event resulting in a moderate increase in MIC depending on the dose administered. 18 So, the ciprofloxacin dosage regimen received by patients in previous outbreaks also could be responsible for different MIC values observed in the same strain.
It was observed that substitution of amino acid 90 resulted in different levels of drug resistance. M. massiliense presenting Ala-90→Val mutation showed high values of MIC (8 and >16 μg/mL). In contrast, the lowest MIC (4 μg/mL) was found in isolates that had an Ala-90. The MIC values of FQ in isolates that contained an amino acid change (Ala-90→Val) were 2–3 times higher than that against isolates presenting Ala-90. This value is lower than those found in a previous study with M. tuberculosis, in which the MIC of quinolones against Ala-90→Val mutated strains was eight times higher than Ala-90 strains. 24 Although such findings indicate that gyrA mutations may be associated with high-level ciprofloxacin resistance in M. massiliense, differences in cell wall permeability 21 and natural efflux pumps 4 could account for the variances in FQ susceptibility. The presence of double-codon mutation in M. chelonae isolate was not correlated to high-level resistance to FQ, which has been previously described. 39
In our study, mutation in gyrA gene was highly observed, but mutation was not found in gyrB gene; this fact was similar to higher mutation rate observed in gyrA when compared with gyrB gene in M. tuberculosis isolates.19,32 The low prevalence of gyrB gene mutation may suggest that such alterations are secondary to that of gyrA gene regarding FQ susceptibility.
Analysis only based on rpoB gene was reported as not being sufficient to differentiate closely related mycobacterial species 25 and molecular method targeting erm(41) has been described to be efficiently used for the differential identification of M. massiliense from M. abscessus and M. bolletii. 22 In this study, nucleotide sequences of M. massiliense gyrA (120 bp) and gyrB (117 bp) QRDRs were identical with sequences presenting Ala-90 and sequences showing Ala-90→Val substitution, and discriminate M. massiliense from available sequences of other mycobacterial species,15,11 even if our gyrA and gyrB sequences have shown a high similarity of 98.3% and 97.4%, respectively, when compared with M. abscessus sequence. 11
The presence of a single RGM strain 25 in Brazilian hospital surgical environment allows us to assume that a high frequency of an Ala-90→Val substitution found in our isolates could be a characteristic mutation in M. massiliense strain INCQS 594.
In conclusion, we have found a relatively frequent point of mutation (Ala-90→Val) in gyrA gene and no mutation in gyrB gene in ciprofloxacin-resistant Mycobacterium massiliense isolates. Further studies should be conducted to fully establish the M. massiliense susceptibility to FQ.
Footnotes
Disclosure Statement
The authors of this article, Fernanda Monego and Alexander Welker Biondo, declare that this study has no conflict of interest.