
Abstract
An increased incidence of enrofloxacin-resistant Escherichia coli associated with septicemic colibacillosis in calves was observed recently in northern Italy. The aim of this study was to investigate this phenomenon. A total of 47 consecutive E. coli isolates exhibiting reduced susceptibility to enrofloxacin (intermediately resistant or resistant) causing septicemic colibacillosis in calves from 45 large-scale farms during 2006–2008, were studied. Phylogenetic group, antimicrobial agents susceptibility, and O serogroup were determined with randomly amplified polymorphic DNA (RAPD) and pulsed-field gel electrophoresis (PFGE) typing, providing additional discrimination. All of the microorganisms carried resistance to two or more additional drugs, with the pattern fluoroquinolone–ampicillin–co-trimoxazole–tetracycline–gentamicin–thiamphenicol being the most represented (18/47; 38.3%). Plasmid-mediated extended-spectrum and AmpC β-lactamases and plasmid-mediated fluoroquinolone resistance genetic determinants were not detected. Third-generation cephalosporins emerged as the most active antimicrobial agents tested (97.9% of susceptible strains). Overall, 37 different RAPD profiles and 18 different O serogroups could be distinguished among the typeable strains, indicating a substantial heterogeneity and suggesting the occurrence of several independent selection events. However, approximately one-fourth (11/47) of the strains belonged to serogroup O78, and PFGE revealed that the great majority (7/11) of these were clonally related, indicating the selection of a O78 clonal group. This is the first report investigating the molecular epidemiology of fluoroquinolone-resistant E. coli in calves and describing the emergence of a fluoroquinolone-resistant E. coli clonal group in these animals.
Introduction
The increased incidence of enrofloxacin-resistant E. coli strains, causing septicemic colibacillosis in young calves observed recently in our area (from 14.3% in 2002 to >40% in 2008) (S. Zoppi, Istituto Zooprofilattico Sperimentale del Piemonte Liguria e Valle d'Aosta, unpublished data) prompted this study with the aim of understanding the basis of this phenomenon through molecular characterization.
Materials and Methods
Bacterial strains
Forty-seven E. coli displaying reduced susceptibility to enrofloxacin (zone diameter ≤18 mm, enrofloxacin disc 5 μg) were selected from 137 strains causing septicemic colibacillosis in young calves [mean age, 9.87 days±9.88 (mean±standard deviation)]. All strains were isolated from blood and tissue samples (spleen, liver, and bone marrow) from septicemic animals bred in 45 widely dispersed large-scale farms in Piemonte, a region in the northwest of Italy. Strains were consecutively isolated during the years 2006–2008 at the Istituto Zooprofilattico Sperimentale in Turin, Italy, and then referred to the Microbiology Section (DISC Department) of the University of Genoa, Italy, where they were reidentified to the species level using the API 20E system (bioMèrieux, Marcy l'Etoile, France) and stored at −80°C until further analysis.
Susceptibility tests
Susceptibility to selected antimicrobial agents was determined by using the disk diffusion test and interpreted according to the Clinical Laboratory Standards Institute (CLSI) criteria for bacteria isolated from animals, or, when not available, according to the CLSI criteria for bacteria isolated from humans. 8 Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 were used as quality-control strains.
β-Lactamase studies
To detect extended-spectrum β-lactamase
O serotyping
O serotyping was performed using E. coli O antisera purchased from Denka Seiken, Co Ltd (Tokyo, Japan), according to the manufacturers' instructions.
Virulence genotyping
All of the isolates were screened for the virulence markers astA, pap, iucD, afaD8/E8, eae, bfpA, cnf1/2, and stx1/2 using previously described primers and amplification conditions.2,15,16
Phylotyping
E. coli phylogenetic group (A, B1, B2, or D) was determined by an established multiplex PCR assay described by other authors and as reported previously. 5
Screening for plasmid-mediated quinolone resistance genes
All isolates were screened for the presence of qnr (qnrA, qnrB and qnrS), aac(6′)-Ib-cr, and qepA genes using previously described primers and amplification conditions.5,15 Plasmids pmG252 (qnrA 1), pmG298 (qnrB 1), pmG306 (qnr S 1), pPSTV28 (qepA), and pBCSK(+) (aac(6′)-Ib-cr) were used as positive controls.
Randomly amplified polymorphic DNA and pulsed-field gel electrophoresis
Randomly amplified polymorphic DNA (RAPD) and pulsed-field gel electrophoresis (PFGE) analysis, and the interpretation of results (banding patterns) was carried out as previously described. 5
Outer membrane protein profiles
To ascertain if resistance to third-generation cephalosporins and cefoxitin (strain E. coli Eco 28) was due to impermeability, outer membrane protein (OMP) profiles analysis was carried out. OMP preparations were obtained after sonic disruption of bacterial cells grown in nutrient broth, followed by selective solubilization of cytoplasmic material with sodium N-lauroyl sarcosinate (2% w/v) and ultracentrifugation as previously described. 5 The preparations were run on sodium dodecyl sulfate–polyacrylamide gels (12%) and stained with Coomassie Blue. Escherichia coli strain HB 101 and E. coli ATCC 25922 were used as fluoroquinolone-susceptible controls.
PCR amplification of the E. coli Eco 28 ampC promoter
Primers AB1 (5′-GATCGTTCTGCCGCTGTG-3′) and AmpC2 (5′-GGGCAGCAAATGTGGAGCAA-3′) were used to amplify a 271-bp fragment from the E. coli ampC promoter, as described by Caroff et al. 6 The PCR products were purified and sequenced as previously described. 5
PCR amplification and DNA sequencing of gyrA and parC
The gyrA and parC gene fragments of four O78 E. coli strains, representing four nonidentical PFGE profiles and four different levels of fluoroquinolone resistance [ciprofloxacin minimum inhibitory concentration (MIC) of 0.5, 4, 32, and >128 mg/L, respectively] were amplified by using previously described PCR conditions and primers. 5 PCR amplimer purification and sequencing were carried out as described above.
Conjugation experiments
Conjugal transfer of ampicillin, tetracycline, co-trimoxazole, gentamicin, and thiamphenicol resistance among all strains and E. coli K12 J53 Rif R was attempted by filter mating. Transconjugants were selected on rifampicin (256 mg/L) and either ampicillin (100 mg/L) or tetracycline (10 mg/L), or chloramphenicol (20 mg/L) or gentamicin (2 mg/L) or trimethoprim (5 mg/L).
Plasmid extraction
Plasmid DNA was extracted by the alkaline lysis method. 19
Results
Susceptibility tests
Table 1 shows the disk diffusion susceptibility patterns of E. coli strains with reduced susceptibility to enrofloxacin. Third-generation cephalosporins (cefotaxime and ceftazidime) emerged as the most active drugs tested (97.9% of susceptible strains). Only one strain (Eco 28) was resistant to ceftazidime and cefotaxime. Overall, >90% of the strains were susceptible to cefoxitin. In contrast, the activity of the remaining antimicrobial agents was dramatically reduced (<50% of susceptible strains).
S, susceptible; I, intermediate; R, resistant.
All microorganisms studied carried resistance to two or more additional drugs, with the pattern fluoroquinolone–ampicillin–co-trimoxazole–tetracycline–gentamicin–thiamphenicol being the most represented (18 out of 47 strains; 38.3%), (Table 2). All strains belonging to the O78 clonal group were simultaneously resistant to enrofloxacin, ampicillin, co-trimoxazole, tetracycline, and thiamphenicol.
AMP, ampicillin; SXT, co-trimoxazole; TC, tetracycline; TH, thiamphenicol; GEN, gentamicin; FOX, cefoxitin.
β-Lactamase studies
The only strain simultaneously resistant to cefotaxime, ceftazidime, and cefoxitin (Eco 28) was positive by the CLSI ESBL initial screen test, but displayed the AmpC phenotype. This strain was PCR negative for AmpC plasmid-mediated genes. Molecular characterization by PCR showed that none of the strains carried bla SHV , bla CTX-M , genes, whereas 41 strains carried a bla TEM gene.
Serotyping
Thirty-nine out of the 47 strains were typeable, 6 were autoagglutinating, and 2 were untypeable. A high variability was found among our collection (18 different serotypes/39 typeable strains). Overall, the most common serogroup was O78 (11 strains), followed by O111 (4) and O63 (3) (Table 2).
Detection of virulence-associated genes
The isolates produced one or more of the following potential virulence factors: The astA gene was detected in 18 strains, 19 possessed the pap gene, 31 the iucD gene, and 14 the afaD8/afaE8 genes. No strain was found positive for the other virulence-associated genes tested.
Phylogenetic groups
Approximately 70% of the strains (72.3 %) belonged to the less-virulent commensal phylogenetic group A and the remaining 27.7% to virulent phylogenetic group D, as shown in Table 2.
Quinolone plasmid-mediated resistance
By PCR analysis, isolates carrying qnr-like or aac(6′)-Ib-cr and qepA genes were not detected.
RAPD PCR profiles
All 47 strains underwent RAPD PCR with two different primers. Overall, 37 different profiles could be distinguished by this method, The most-represented RAPD profile, designated A, was related to serogroup O78 and was found in seven strains. Three strains belonging to serogroup O63 had the same RAPD profile (Table 2).
PFGE profiles
Finer resolution of the clonal relationship among the 11 O78 E. coli strains and the 3 O63 E. coli strains was obtained by PFGE analysis. Seven O78 strains were related (differing from 0 to 6 bands). The remaining 4 O78 strains had a different unrelated profile. Of the 3 O63 E.coli strains, 2 had a related profile (Fig. 1).
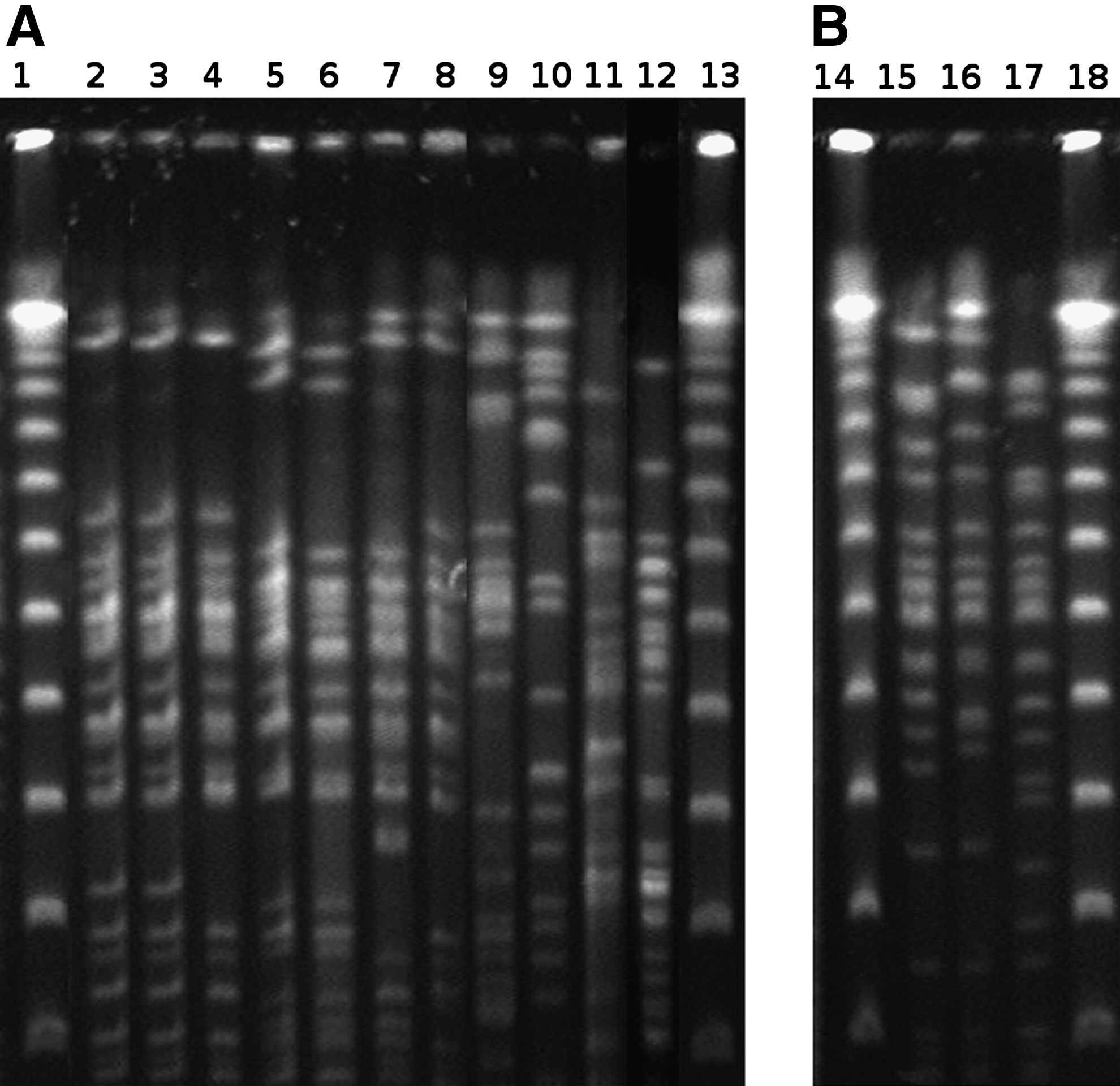
Pulsed-field gel electrophoresis (PFGE) patterns of O78 (
OMP profiles
The only cefoxitin-resistant E. coli (Eco 28) isolate expressed both the OmpC and OmpF proteins. Because OMP loss was not observed in this strain, its phenotype was compatible with hyperproduction of chromosomal cephalosporinase.
PCR amplification of the E. coli Eco 28 ampC promoter
The promoter of E. coli Eco 28 differed from strain K 12 at bases −88, −82, −42, −18, −1, and +58. All of these mutations have been already described in E. coli strains characterized by increased ampC transcription and hyperproduction of chromosomal cephalosporinase. 6
PCR amplification and DNA sequencing of gyrA and parC
Compared to the amino acids in wild-type E. coli K12, one out of the four isolates analyzed (ciprofloxacin MIC 0.5 mg/L) had a single mutation in gyrA, resulting in alterations at positions Ser-83 (Ser→Leu). Two isolates with ciprofloxacin MIC 4 and 16 mg/L, respectively, possessed two mutations in gyrA and one mutation in parC, resulting in alterations at positions Ser-83 (Ser→Leu) and Asp-87 (Asp acid→Asn) in GyrA and positions Ser-80 (Ser→Ile) in ParC, respectively. The fourth strain (ciprofloxacin MIC >128 mg/L) had two mutations in gyrA (the same observed in the other strains studied) and two mutations in parC, resulting in alterations at position Ser-80 (the same observed in the other strains studied) and position Glu-84 (Glu→Val).
Plasmid analysis
Conjugation experiments and antimicrobial susceptibility testing of the transconjugants revealed that only 4 out of 47 (8.5%) strains were able to transfer ampicillin and tetracycline resistance to the recipient. Ampicillin resistance and tetracycline resistance were transferred simultaneously. No detectable transfer was observed for the other resistance markers tested (trimethoprim, gentamicin, and chloramphenicol). The plasmid profile analysis of the four transconjugants revealed three distinct profiles.
Discussion
Septicemic colibacillosis in calves, sustained by E. coli, is usually the main infectious problem of large-scale farms and is responsible for significant losses. 12 Fluoroquinolones and other “new-generation” antimicrobial agents (third- and fourth-generation cephalosporins and new aminoglycosides) are often used as the first choice among veterinarians for neonatal enteritis treatment in Italy. 4 Moreover, the incorrect use of these antimicrobial agents is also common for the prevention of neonatal enteritis in our country. 4
Recently, an increased incidence of enrofloxacin-resistant E. coli strains associated with septicemic colibacillosis in young calves was observed in Italy. 3 Serotyping, phylogenetic grouping, RAPD, and PFGE results showed that in the currently observed area several independent selection events occurred, including the selection of a multidrug-resistant O78 clonal group.
Recently, plasmid-mediated quinolone resistance has been described both in human and poultry isolates. 21 To our knowledge, no report concerning plasmid-mediated fluoroquinolone resistance mechanisms in bovine isolates is available, and, in our isolate collection, these mechanisms were not found to contribute to the reduced susceptibility to fluoroquinolones observed. In different hosts (i.e., humans, calves, and chicken), different antimicrobial selective pressure, due to different antimicrobial agent dosage and consumption habits and the circulation of different clones, may all have contributed to the selection of different resistance mechanisms with variable frequency.
Strains belonging to serogroup O78 were quite common in our collection (11/47; 24%), confirming the important pathogenic role played by this serogroup in animals.1,11,13,17 E. coli serogroup O78 strains seem to have the ability to gain and express several virulence properties and act bifunctionally as the causative agents of septicemic as well as of intestinal infections. 11 In addition, phylogenetic studies have demonstrated clonal overlap between pathogenic O78 isolates from humans and animals, suggesting that animals are a possible source of serogroup O78 E. coli infections in humans.1,11
The emergence of multiresistance involving fluoroquinolones among E. coli strains belonging to such a versatile serogroup is of great concern and needs to be monitored. Further studies are needed to clearly determine the zoonotic potential of these strains.
To our knowledge our study is the first investigating the molecular epidemiology of fluoroquinolone-resistant E. coli strains in calves, and it represents the first report describing the emergence of a multiresistant fluoroquinolone-resistant E. coli clonal group causing septicemic colibacillosis in calves. The selection of main clusters of fluroquinolone-resistant O78 avian pathogenic E. coli has been described in Japan and France.13,17
Among the members of the O78 clonal group studied, we found the same mutations in gyrA and parC genes already described for avian strains.13,17 The number of strains analyzed was limited; however, this additional information can contribute support to the hypothesis of a common origin for O78 isolates.1,11
Simultaneous resistance to ampicillin (ascribable to broad-spectrum TEM β-lactamase), tetracycline, co-trimoxazole, gentamicin, and thiamphenicol was widely diffused in our collection. Only in a few cases were resistance traits found to be transferable, suggesting a limited role of conjugative plasmid in the spreading of multiresistant E. coli strains.
Although other authors have found plasmid-mediated AmpC and ESBLs able to hydrolyze third-generation cephalosporins in isolates of animal origin, 9 no strains carrying the genes related to these phenotypes were found. It is reassuring that cephalosporins have maintained their activity against our collection of strains, thus supporting their use for the treatment of diarrhea in calves. However, due to the recent introduction of third- and fourth-generation cephalosporins in veterinary medicine and the detection of one strain hyperproducing chromosomal cephalosporinase in this collection, specific surveillance studies are recommended in the near future.
The adoption of strict hygienic measures at farms (the veterinarians were asked to always use gloves and other disposables to visit the animals) and the avoidance of animal exchanges among different farms have successfully prevented the spread of a single plasmid or a single resistant clone. However, further efforts to improve veterinary prescribing habits in our area are needed.
Footnotes
Acknowledgments
We are indebted to: Dr. C. Grattarola, Istituto Zooprofilattico Sperimentale del Piemonte, Liguria e Valle d'Aosta (IZS PLVA), Turin Italy, for her technical help for necroscopic examinations of calves; Professor G.A. Jacoby, Lahey Clinic, Burlington, Massachussets, for providing us with E. coli J53 pmG 252, E. coli J53 pmG 306, and E. coli J53 pmG 298; Professor Kunikazu Yamane Laboratory of Antimicrobial Agents and Resistance Department of Bacterial Pathogenesis and Infection Control National Institute of Infectious Diseases, Tokyo, Japan for providing us with plasmids pPSTV28 (qepA) and pBCSK(+) (aac(6′)-Ib-cr).
Disclosure Statement
All of the authors disclose that they have no commercial associations that might create a conflict of interest in connection with this article.