
Abstract
Staphylococcus aureus produces virulence factors, including various exotoxins and adhesins, which are associated with a variety of symptoms caused by its infections. In the present study, the prevalence of these virulence factors was analyzed for 23 S. aureus strains isolated from wound infections in hospitals, nasal swabs, or vomit from patients and cooks in a food poisoning case and from healthy adults in Yangon, Myanmar. Among these strains, five were methicillin-resistant S. aureus (MRSA) derived from pus (four strains, SCCmec III, ST239) and a healthy adult (one strain, SCCmec-IVa, ST5). The Panton-Valentine leukocidine (PVL) gene was detected in five methicillin-susceptible S. aureus (MSSA) clinical strains belonging to ST121 (CC121). The MRSA clinical strains had only a few or no staphylococcal enterotoxin (SE) genes, whereas PVL-positive MSSA and an MRSA strain from a healthy adult possessed an enterotoxin gene cluster (seg, sei, sem, sen, seo, and selu). Strains from the food poisoning case had either SE genes or only etd and edin-B. Adhesin genes, which are associated with binding to fibronectin, fibrinogen, and elastin, were detected in all the MRSA and most of the MSSA strains examined. However, the bone sialoprotein-binding protein gene (bbp) and the variant form of the elastin-binding protein gene (ebpS-v) with an internal 180 bp deletion were identified only in the MSSA strains harboring the PVL gene. These findings suggest that those genetic traits are characteristic of PVL-positive ST121 S. aureus strains in Myanmar.
Introduction
A variety of symptoms caused by S. aureus infections are associated with exotoxins and virulence factors produced by this organism. Staphylococcal food poisoning, which is characterized by vomiting and diarrhea, is caused by intake of food or beverages contaminated with one or more staphylococcal enterotoxins (SEs). 18 To date, genetically distinct SEs, SEA-SEE, SEG-SEI, and SER-SET have been identified, together with genes encoding SE-like toxins (selj-selq, selu, and selv).35,48,64 Toxic shock syndrome toxin-1 (TSST-1) is considered the most common cause of toxic shock syndrome 59 and has superantigenic activity similar to SEs. 45 Despite lacking superantigen capability, clusters of multiple genes designated as staphylococcal superantigen-like (SSL) proteins were also reported.36,70 Exfoliative toxin (ET) causes cleavage of epidermal cells typically seen in staphylococcal scalded skin syndrome (SSSS) and bullous impetigo, and three types of ETs (ETA, ETB, and ETD) have been described in S. aureus isolates from humans. 71 Epidermal cell differentiation inhibitor (EDIN) is also an exotoxin associated with induction of hyperplasia of the epidermis and is classified into three types (A-C). 15 Panton-Valentine leukocidine (PVL), which has been considered one of the factors related to severe symptoms of CA-MRSA infection,37,72 has cytotoxicity against human leukocytes and is possibly related to the development of necrotizing lesions.
In addition to these exotoxins, S. aureus produces other virulence factors, adhesins, which are composed of different protein families and mediate attachment of S. aureus to different types of cells. These factors are designated also as microbial surface components recognizing adhesive matrix molecules. 21 Some are found in all the S. aureus strains (core adhesins), whereas the others are distributed to particular strains (accessory adhesins).65,68
Prevalence of the virulence factor genes in S. aureus isolates has been mostly investigated in developed countries and was revealed to be different depending on methicillin resistance, sources of isolates, or clones.7,27,49,53,62 In contrast, only limited information is available for isolates in developing countries. In Myanmar, genetic study of S. aureus that can elucidate the SCCmec type and sequence type (ST) of MRSA, as well as the distribution of virulence factors including PVL, has not been yet conducted. In the present report, as a first study in Myanmar, 23 representative strains from different sources were investigated to clarify the prevalence of virulence determinants and their genetic characteristics in S. aureus isolates. The results provide new insights into the virulence factors in PVL-positive S. aureus.
Materials and Methods
Bacterial isolates
A total of 23 S. aureus strains derived from clinical specimens (15 strains), samples from a food poisoning case (5 strains), and nasal swabs from healthy adults (3 strains) were analyzed. The 15 clinical strains were isolated from skin infections (pus) of patients at a general hospital in Yangon during a period from November 2007 to September 2008. In the food poisoning case, which occurred in October 2008 in Yangon, the five strains were isolated from different sources: three isolates from hand swabs of cooks (strains Y27, Y28, and Y29), an isolate from vomit of a patient (Y30), and an isolate from the surface swab of a table contaminated with food (Y31). Three strains from healthy adults were obtained from nasal swabs of food handlers at the three different hotels in Yangon, in May 2008, when a routine inspection for hotel workers was held. All the strains were kept in Microbank (Pro-Lab diagnostics, Richmond Hill, ON, Canada), stored at −80°C, and recovered when they were analyzed.
Antimicrobial susceptibility testing
Minimum inhibitory concentrations of representative antimicrobial agents (oxacillin, ampicillin, cefoxitin, vancomycin, gentamycin, kanamycin, fosfomycin, tetracycline, minocycline, ciprofloxacin, clindamycin, and erythromycin) were measured by a broth microdilution test and interpreted as sensitive or resistant based on the National Clinical and Laboratory Standards Institute guidelines.12,13
Multiplex polymerase chain reaction assays for MRSA and SCCmec typing
Presence of the staphylococcal 16s rRNA gene, nuc, mecA, and PVL gene (lukS-PV/lukF-PV) was investigated by multiplex polymerase chain reaction (PCR) assay as described by Zhang et al. 74 for all the S. aureus isolates. The SCCmec type of MRSA was determined by PCR as previously described. 33 SCCmec type II and IV elements were further analyzed for their subtypes by PCR with the use of primers previously reported.26,74 Presence of the PVL gene was confirmed by PCR using the primers described by Lina et al., 37 and other leukocidine genes lukE-lukD and lukM were examined by PCR with primers as previously described. 29
Coagulase gene typing, agr typing, and multilocus sequence typing
Coagulase genotypes, including subtypes of IV and V, were determined by multiplex PCR assay as previously described. 25 Subtypes of coagulase genotypes VII were determined by PCR and direct sequencing of the coagulase gene D1 region, as previously described. 25 The accessory gene regulator (agr) typing (subgroups I, II, III, and IV) was performed as previously described.53,61 For selected strains, ST based on the scheme of multilocus sequence typing was determined as previously described. 20 ST data were further analyzed by eBURST to determine their clonal complexes (CCs).
Detection of virulence factor genes and drug resistance genes
Genes for various exotoxins and hemolysins were detected by monoplex or multiplex PCR using PCR primers and in conditions previously reported.4,11,22,27–30,34,43,47–49,65,71 The presence of most exotoxin genes was confirmed by PCR using two different primer pairs previously described. The genes analyzed and their references of PCR primer sequences are as follows: 5 hemolysin genes (hla, 29 hlb, 30 hlg, 29 hlg-v, 29 and hld 30 ), 20 genes of SE or SE-like toxins (sea,4,28 seb,4,29 sec,28,29 sed,4,29 see,4,28 seg,29,47 seh,29,43 sei,29,47 selj,27,29 selk,27,28 sell,22,27 selm,28,29 seln,28,29 selo,22,29 selp,22,27 selq,11,27 ser,22,28 ses,30,48 set,30,48 and selu11,28), 3 ET genes (eta,4,71 etb,4,71 and etd 71 ), TSST-1 gene (tst-128,71), and 2 EDIN genes (edin-A 29 and edin-B 71 ). For detection of etd, an additional primer pair etd-n1 (5′-CTAAAGTTATATTCACACCT-3′) and etd-n2 (5′-GTAGAAAAGTTATAAGGGTATC-3′) designed in this study was used. The presence of genes in the SSL protein cluster (set6, set8/9, set10, set13, and set15; nomenclature of genes is based on genomic island nuSa-alpha of MRSA strain N3153) was also examined by PCR with the use of primers described by Holtfreter et al. 27 and a primer pair for set6 designed in the present study (set6-f: 5′-AAAAGCAAGTTTAGCATTGGGA-3′; set6-r: 5′-GTATTTGTCTGCCGTCGATAG-3′).
The presence of other virulence factor genes including adhesins was also examined by PCR. The targeted proteins (genes) and their references of PCR primer sequences are as follows: biofilm-associated protein (bap 68 ), collagen-binding protein (cna 65 ), laminin-binding protein (eno 65 ), fibronectin-binding protein (fnbA 68 , fnbB7,34,68), elastin-binding protein (ebpS 68 ), clumping factor (clfA 65 , clfB 65 ), fibrinogen-binding protein (fib 65 ), serine-aspartate repeat protein (sdrC7,49, sdrD7,49, sdrE7,49), and bone sialoprotein (bbp 65 ). The icaA and icaD genes, which are associated with biofilm formation, were also examined by PCR using the primers previously described. 69 The fnbB is known to be diverse depending on the domains 6 ; as such, three pairs of PCR primers specific to different domains7,34,68 were used to detect this gene.
Resistance genes to penicillin (blaZ), tetracycline [tet(K), tet(L), and tet(M)], macrolide (ermA, ermB, ermC, and msrA), and aminoglycoside [aac(6′)-Im, aac(6′)-Ie-aph(2″)-Ia, ant(3″)-Ia, ant(4′)-Ia, ant(6)-Ia, ant(9)-Ia, ant(9)-Ib, aph(2″)-Ib, aph(2″)-Ic, aph(2″)-Id, and aph(3′)-IIIa] were detected by monoplex or multiplex PCR using the primers previously reported.39,40,46
Sequence analysis of ebpS
Nucleotide sequences of the ebpS genes were determined by PCR and direct sequencing as previously described. 67 A DNA fragment containing full-length ORF of ebpS was amplified by PCR with primers ebpS-1 (5′-CATAAACATATGTCATGTGGGT-3′) and ebpS-2 (5′-GCACGTTCGTATATAGCAGTTC-3′). Genetyx Version 5.1 (Software Development, Tokyo, Japan) was used to perform pairwise alignment and calculate sequence identity of ebpS genes from different strains. Multiple alignments of nucleotide and amino acid sequences of ebpS were performed by the Clustal W program.
GenBank accession numbers
The nucleotide sequences of ebpS genes from strains Y6, Y12, and Y18 have been deposited in the GenBank database under accession numbers JF706228, JF706229, and JF706230, respectively.
Results
Among the 23 strains analyzed, four strains isolated from pus and one strain from a healthy adult were MRSA (Table 1). The four MRSA clinical isolates had type III SCCmec and belonged to coagulase gene type IVa and agr type I. Among these MRSA, two strains, Y3 and Y18, were assigned to ST239 (CC8). MRSA strain Y6 from an adult had type IVa SCCmec and was classified into coagulase genotype II, agr type II, and ST5 (CC5). A PVL gene was detected in five MSSA strains from pus, which belonged to coagulase genotype Va, agr type IV, and ST121 (CC121) (strains Y12, Y16, and Y21). Five strains from a food poisoning case were MSSA, and only two strains Y27 and Y30 had the same genotype (coagulase type II and agr type III).
Staphylococcal superantigen-like proteins.
Five hemolysin genes were detected in all the strains, but lukM, sed, see, seh, eta, etb, and tst-1 were not identified in any strain (Table 1). Three MRSA clinical strains had no SE genes, and a remaining strain Y3 harbored three SE genes (sea, sek, and seq). Similarly, most of the PVL-negative MSSA strains possessed few or no SE genes. In contrast, five PVL-positive strains and an MRSA from a healthy adult possessed six SE genes, which constituted enterotoxin gene cluster 14 (egc2: seg, sei, selm, seln, selo, and selu). Among the strains from a food poisoning case, SE genes were detected in only two strains (Y28 and Y29) from cooks with different SE gene profiles. In a strain from the vomit of a patient (Y30) as well as a strain from a cook (Y27), etd and edin-B were detected, despite the absence of the SE gene. Any gene in the SSL cluster was detected in all the strains examined, although profiles of detected genes were different depending on the strains.
Seven adhesin genes (eno, fnbA, ebpS, clfA, clfB, fib, and sdrC), icaA, and icaD were detected in all the strains, but no strain harbored the bap gene (Table 2). fnbB and sdrE genes were detected in all strains, except for a single MSSA strain. The sdrD gene was detected in all MRSA strains and MSSA strains derived from the food poisoning case and healthy adults, but was not found in most of MSSA from wounds, including those with the PVL gene. In contrast, the bone sialoprotein-binding protein gene (bbp) was detected in only PVL-positive strains. Collagen-binding protein gene cna was detected in MRSA clinical strains and PVL-positive MSSA, but was absent in most of the other strains.
The ebpS gene with internal deletion (ebpS-v).
Although the elastin-binding protein (EbpS) gene (ebpS) was detected in all the isolates examined, PCR products from all five PVL-positive strains were shorter (∼470 bp) than the expected size (652 bp), with the primers described by Tristan et al. 65 (data not shown), suggesting genetic alteration in these strains. Accordingly, full-length sequences of the ebpS gene were determined for PVL-positive strains Y12, Y13, Y16, and Y21 and PVL-negative strains Y6 and Y18. The ebpS genes of the Y6 and Y18 strains had the same length (1,458 bp, 486 amino acids) as those previously reported 19 and showed 97%–100% nucleotide (nt) sequence identities to those of representative S. aureus strains Cowan and Newman (GenBank accession nos. U48826 and AP009351, respectively). However, the ebpS sequences of the four PVL-positive strains were 1281 nt in length, which may encode the 427-amino acid (aa) product, showing absolute sequence identity among the four strains. These ebpS genes were provisionally designated ebpS-v. Sequence alignment revealed that in comparison with the ebpS sequence of the Cowan strain, ebpS-v has 3 nt (GCG) insertion after nt 288 of Cowan ebpS, causing an additional amino acid between positions 96 and 97 of EbpS (Fig. 1). Further, a 180-bp sequence corresponding to nt 586–765 of ebpS was deleted in ebpS-v, causing a 60 aa deletion from the intact EbpS, 19 in comparison with those of other representative strains (Figs. 1 and 2). The deleted portion contained the H1 region, one of the three hydrophobic domains of EbpS. It was of note that a 57-nt sequence at the 5′-end side of the deleted region (nt 586–642 of ebpS) was similar to the 57-nt sequence located just after the deleted portion (nt 766–822) (Figs. 2 and 3). The 57-nt sequence of the strain Y12 corresponding to ebpS nt 766–822 had a 96.5% identity to those in this site of other strains and 87.7%–94.7% to sequences at the nt 586–642 of these strains. Two conserved nucleotides at positions 795 (T) and 804 (C) were different from the corresponding nucleotides in the nt 586–642 portion, that is, nt 615 (A or C) and 624 (T), respectively.

Alignment of EbpS amino acid sequences from strains Cowan, Newman, and three strains isolated in Myanmar (Y6, Y18, Y12). EbpS sequences of strains Y13, Y16, and Y21 were omitted, because they are identical to that of strainY12. Amino acid numbers based on EbpS of strain Cowan are shown above the sequence. Asterisk indicates conserved amino acid among all the strains, and dash denotes gap. A closed circle and a shaded region indicate an additional amino acid and deleted portion in Y12-EbpS (EbpS-v), respectively. LBD near the N-terminus represents ligand-binding region of EbpS, and H1, H2, and H3 denote three hydrophobic domains.

Alignment of partial nucleotide sequences of ebpS (nt 551–850) from representative Staphylococcus aureus strains (Cowan, Newman, TW20, TCH1516, and Mu3) and three strains in Myanmar (two strains Y6 and Y18 with intact ebpS, and strain Y12 having the ebpS variant, i.e., ebpS-v). Nucleotide numbers based on ebpS of strain Cowan are shown above the sequence. Asterisk below the alignment indicates conserved nucleotide among all strains. Deleted region in the strain Y12 is shown by dashes (nt 586–765). Underlined 57-bp region of ebpS-v (corresponding ebpS nt 766–822) is arranged in a box at the 5′-end portion of the putative deleted region (nt 586–642) to show sequence similarity; asterisk below the box indicates identical nucleotide to strains shown above. Two conserved nucleotides at positions 795 (T) and 804 (C) indicated by # are not observed in the corresponding positions (615 and 624, respectively) in the nt 586–642 region.
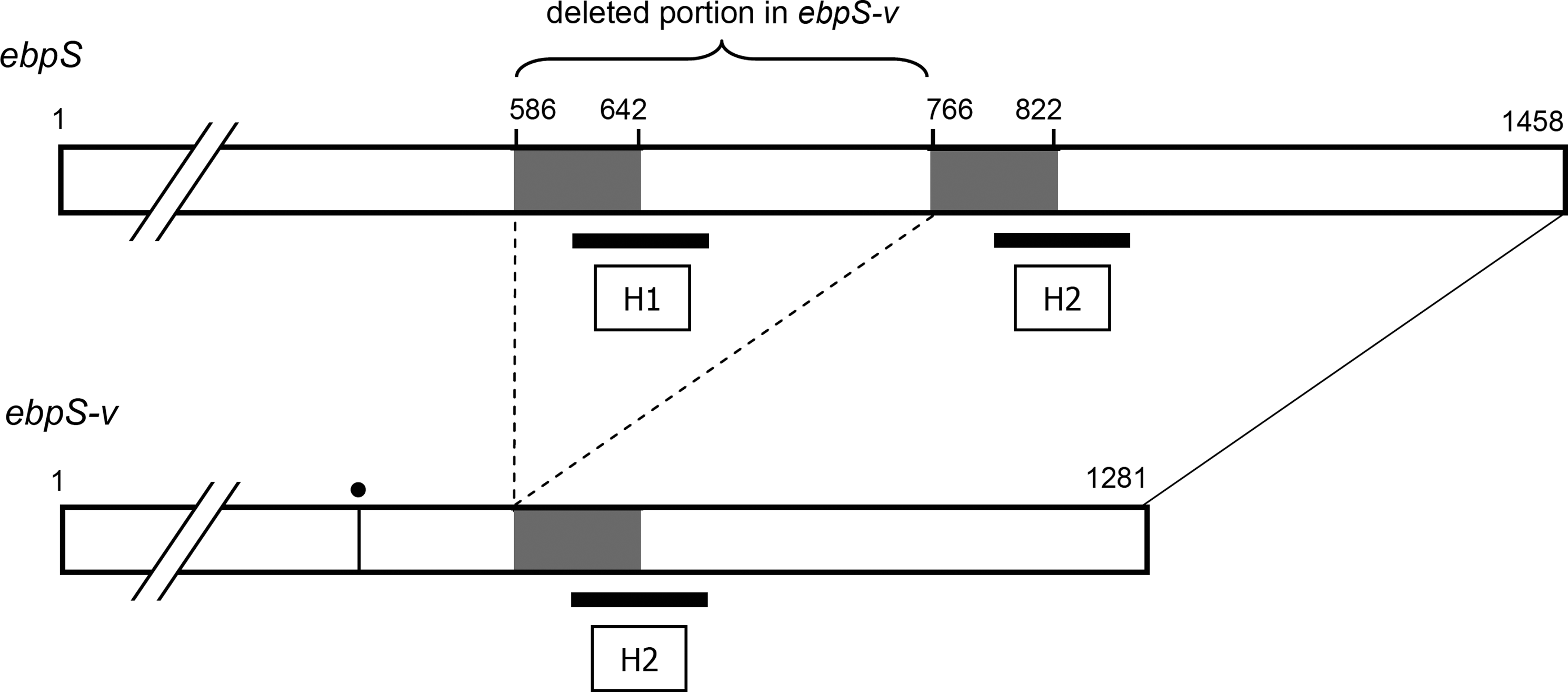
Schematic representation of structures of the intact ebpS gene and the ebpS-v with an internal deletion. Nucleotide numbers are indicated above the genes. The 57-bp regions in ebpS, which show similar sequences (Fig. 2), are shaded. Putative sites of rearrangement in ebpS-v and both ends of the deleted portion, which is present in the intact ebpS, are connected by dotted lines. Hydrophobic regions H1 and H2 are shown by thick horizontal lines. Vertical line with dot in the ebpS-v represents an additional codon at nt position 289–291.
Four MRSA clinical strains were resistant to more antimicrobials and possessed more resistance genes than MSSA strains including PVL-positive strains (Table 3). tet(K) and tetracycline resistance was found in most MSSA strains, but some resistance genes, for example, tet(M), ermA, and aph(3′)-III, were detected only in MRSA clinical strains. MRSA strain Y6 from a healthy adult was susceptible to gentamicin and ciprofloxacin and possessed ermC and ant(6)-Ia, which were distinct characteristics from MRSA clinical strains.
MIC, minimum inhibitory concentration.
Discussion
Because the spread of HA- and CA-MRSA is a global concern, their prevalence and genetic characteristics need to be investigated in both developed and developing countries. However, in Myanmar, to date no study has been conducted for either prevailing S. aureus strains or MRSA. In the present study, which is the first of its kind in Myanmar, representative S. aureus strains isolated from different sources were analyzed in terms of genetic characteristics and virulence factors.
The four MRSA clinical strains from pus were found to have type III SCCmec and agr-1 allele and belong to ST239 (two strains). These traits are known for Hungarian/Brazilian HA-MRSA clone widely distributed in Asia, particularly China and Southeast Asia.31,72 Especially in China and Malaysia, ST239 was reported as a predominant clone.9,23 Therefore, this clone was suggested to be common also in Myanmar as HA-MRSA. However, unlike ST239 MRSA (Hungarian clone) in Malaysia, three MRSA strains in Myanmar did not harbor the sea gene, suggesting the presence of genetic variants in this clone. MRSA strain Y6 from a healthy adult belonged to ST5 with SCCmec type IVa and agr-2 allele and had sem and seo. These traits have been typically described as a “classical” pediatric clone, one that is a minor and historically early MRSA clone, and reported to be distributed in Europe, the United States, Canada, and some South American countries.16,24,41,58 In Asia, the confirmed ST5-SCCmec-IV clone was reported only in Korea as HA-MRSA. 31 The strain Y6 is the first pediatric clone isolated from a healthy adult, which may suggest the significance of this clone as CA-MRSA.
In the present study, PVL-positive strains were all MSSA and were classified into ST121-agr-4 allele (three strains). These types of S. aureus strains were reported as one of the major genotypes of PVL-positive MSSA in Germany. 44 Similarly, ST121-MSSA with the PVL gene was also isolated in other European countries2,42 and Asian countries, including Cambodia, Thailand, and Indonesia.10,55,60 Therefore, ST121 may be a widely distributed S. aureus clone harboring the PVL gene. Like PVL-positive MRSA and MSSA reported,1,57,62 PVL-positive MSSA strains in Myanmar harbored egc.
Five strains isolated from a food poisoning case showed different profiles of exotoxin genes. It was of note that strain Y30 isolated from the vomit of a patient had no enterotoxin genes, but harbored etd and edin-B genes, and the same toxin gene profile was found in strain Y27 derived from a cook. The finding that the strains Y27 and Y30 belonged to the same genotypes (coagulase genotype II, agr-III allele) and showed identical adhesion protein gene profiles suggests that they are derived from the same clone, despite differences in the SSL protein gene profiles. Although the strain that was responsible for food poisoning was not conclusive, toxin profiles of strains Y27 and Y30 raise the possibility that ETD may be involved in the occurrence of food poisoning. Coexistence of the etd and edin-B genes was confirmed for Y27 and Y30, which is in agreement with the finding that etd is located in tandem with edin-B in the same locus of chromosome, that is, the etd pathogenicity island. 71 The ETD induces intradermal blister formation by cleavage of desmoglein 1, which may be implicated in cutaneous abscesses and furuncles. 73 However, strains with etd have been detected from patients with bullous impetigo or SSSS at a low rate, but are also found in isolates from sources of other infections, suggesting that etd is related to a broad spectrum of diseases.71,73 Therefore, further investigation on the pathogenic role of ETD, and its possible involvement in food poisoning, may be necessary.
In the present study, unique profiles and genetic characteristics of adhesin genes were revealed for PVL-positive MSSA strains, that is, the presence of bbp and genetic variants of ebpS (ebpS-v). Bone sialoprotein-binding protein (Bbp), which is an allelic variant of SdrE, 66 is a cell wall glycoprotein and mediates binding of S. aureus cells to the bone matrix protein, that is, bone sialoprotein is involved in the pathogenicity of osteomyelitis. It has been documented that bbp is associated with osteomyelitis, and in patients with this disease, a serum antibody against Bbp was detected.49,54,65 In a study of the prevalence of adhesins in PVL-positive CA-MRSA, bbp was present only in pandemic CA-MRSA clones (ST30-SCCmec IVa/IVc) and not in CA-MRSA of continent- or region-specific type including ST1, ST8, ST59, and ST80 clones. 49 However, no information is available for bbp in PVL-positive MSSA clones. As the bbp may possibly enhance the pathogenicity of PVL-positive strains, it will be of significance to examine the prevalence of this gene in MSSA with PVL.
All the PVL-positive MSSA strains examined in this study had an altered EbpS (EbpS-v) encoded by ebpS-v, which differs from intact ebpS in that it lacks an internal 180-bp sequence and has an additional single codon. Although EbpS is a cell surface molecule and has been shown to bind soluble elastin peptides, 50 actual binding of the S. aureus to the elastin tissue is mediated by fibronectin-binding proteins A and B. 56 Thus, the biological function of EbpS and its role in virulence remain to be clarified. EbpS was predicted to have three hydrophobic domains (H1, H2, and H3), among which the H1 and H3 were presumed to span the cytoplasmic membrane. 19 The N-terminal region is exposed on the outside of the cytoplasmic membrane and contains a ligand-binding domain.19,51 The EbpS-v identified in the present study was predicted to lack an entire H1 region, whereas other regions including the N-terminal ligand-binding domain were intact. The results from the study suggest that the EbpS-v may be structurally altered regarding its association with the cell membrane, although functional differences between EbpS and EbpS-v are not evident.
We identified a long direct repeat-like 57-bp sequences in the ebpS gene, at positions nt 586–642 and nt 766–822, which are located at the beginning (5′-end side) of the deleted portion and a portion just after the deletion in ebpS-v. Therefore, because of similarity of these two sequences, it is also possible to presume that the deleted portion may be nt 643–822. However, the 57-bp sequence in ebpS-v was more closely related to nt 766–822 than nt 586–642 of ebpS. Accordingly, we considered that the 57-bp sequence in ebpS-v may have originally existed at the position nt 766–822, and thus the deleted portion may be nt 586–765 (Fig. 3). Presence of the long direct repeat at the beginning and following portions of the deleted region suggests occurrence of template switching of DNA polymerase during nucleotide synthesis as a possible mechanism of the internal deletion, 52 and a similar finding has been previously reported in bacterial and viral genes.32,63 Campoccia et al. reported the 180 bp deletion in ebpS in the strains isolated from implant-related infections in Italy, speculating that a region between cores of H1 and H2 domains may be deleted. 8 However, they analyzed only partial sequence of ebpS (nt 359–1010) (sequence data are not available in any database), and no information is available on methicillin susceptibility, presence of the PVL gene, and ST of those strains. 8 Our present study revealed for the first time the relatedness of the ebpS-v to the PVL-positive MSSA clone ST121. Further investigation will be necessary to elucidate whether the ebpS-v may be intrinsic to specific clone or PVL-positive strains as well as its role in pathogenicity of S. aureus.
Footnotes
Acknowledgment
This study was supported by a Grant-in-Aid for Scientific Research (no. 23590746) from the Ministry of Education, Culture, Sports, Science, and Technology, Japan, and the Heiwa Nakajima Foundation.
Disclosure Statement
The authors of this article have no commercial associations that might create a conflict of interest in connection with the submitted manuscript.