
Abstract
The trimethoprim resistance gene dfrK has been recently described in Staphylococcus aureus, but so far has not been found in other bacteria. A total of 166 enterococci of different species (E. faecium, E. faecalis, E. hirae, E. durans, E. gallinarum, and E. casseliflavus) and origins (food, clinical diseases in humans, healthy humans or animals, and sewage) were studied for their susceptibility to trimethoprim as determined by agar dilution (European Committee on Antimicrobial Susceptibility Testing) and the presence of (a) the dfrK gene and its genetic environment and (b) other dfr genes. The dfrK gene was detected in 49% of the enterococci (64% and 42% of isolates with minimum inhibitory concentrations of ≥2 mg/L or ≤1 mg/L, respectively). The tet(L)-dfrK linkage was detected in 21% of dfrK-positive enterococci. The chromosomal location of the dfrK gene was identified in one E. faecium isolate in which the dfrK was not linked to tet(L) gene but was part of a Tn559 element, which was integrated in the chromosomal radC gene. This Tn559 element was also found in 14 additional isolates. All combinations of dfr genes were detected among the isolates tested (dfrK, dfrG, dfrF, dfrK+dfrG, dfrK+dfrF, dfrF+dfrG, and dfrF+dfrG+dfrK). The gene dfrK gene was found together with other dfr genes in 58% of the tested enterococci. This study suggested an exchange of the trimethoprim resistance gene dfrK between enterococci and staphylococci, as previously observed for the trimethoprim resistance gene dfrG.
Introduction
The most frequent mechanisms of resistance to trimethoprim are (a) decreased permeability or efflux, (b) natural dfr genes mutated, (c) overproduction of sensitive DHFR enzymes, and (d) the production of an additional, trimethoprim-resistant DHFR. 11 This latter mechanism is frequently detected among a variety of Gram-positive and Gram-negative bacteria and the dfr genes encoding these DHFR enzymes are often found on mobile genetic elements. So far, more than 30 dfr genes have been described, which confer high-level resistance to trimethoprim. A vast majority of the currently known dfr genes were detected in Gram-negative bacteria, often associated with gene cassettes in class 1 and class 2 integrons, whereas only few dfr genes have been identified in Gram-positive bacteria. These include (a) the Tn4003-borne gene dfrS1 (also referred to as dfrA) 24 described in Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus hominis, and Staphylococcus haemolyticus, (b) dfrD identified in S. haemolyticus and Lysteria monocytogenes,4,6 (c) dfrG and dfrF shown to be present in S. aureus, Staphylococcus pseudintermedius, and/or enterococci,3,5,22,25 and (d) the recently described dfrK in S. aureus.12,14 The observation that the same dfr gene, such as dfrG, is present in different staphylococcal and enterococcal species strongly points toward an interspecies and intergenus transfer between different Gram-positive bacteria. Up to now, only a few studies analyzed the mechanisms of resistance to trimethoprim in enterococci.3,5,12 As a consequence, there is only limited knowledge of the presence of dfr genes in enterococci.
The objectives of this study were to analyze the susceptibility to trimethoprim in a collection of enterococci of different species and origins and to investigate them for the presence of dfr genes with particular reference to the presence of the novel gene dfrK and its genetic environment.
Materials and Methods
Bacterial isolates
In total, 166 Enterococcus isolates, which comprised 24 vancomycin-susceptible and 142 vancomycin-resistant isolates (vanA/vanB2: n = 93; vanC1/2: n = 49), were included in this study. The isolates belonged to the following species (no. of isolates): E. faecium (88), E. faecalis (15), E. hirae/E. durans (14), and E. gallinarum/E. casseliflavus (49); they were isolated from food (50), humans with clinical diseases (50), healthy human (34), healthy animal (31), or sewage (1) samples. The isolates were obtained during 1991–2007 in previous studies.16,17,23,28,29
Antimicrobial susceptibility testing
Susceptibility to trimethoprim-sulfamethoxazole (1:19) was determined by the disk diffusion test as previously reported. 7 The minimum inhibitory concentration (MIC) of trimethoprim was also determined for all 166 isolates using the agar dilution method with a twofold dilution series of the agent. The MIC was recorded as the lowest concentration of antimicrobial that inhibited the visible growth after 24 hr of incubation at 37°C. Isolates were classified as resistant or susceptible using the interpretive criteria given by the European Committee on Antimicrobial Susceptibility Testing. 7 These interpretive criteria (susceptible: ≤1 mg/L; resistant ≥2 mg/L) are—according to EUCAST—only applicable to enterococci from urinary tract infections in humans. Because of the lack of additional interpretive criteria for enterococci, a reliable classification of enterococci from other sources as susceptible or resistant to trimethoprim is currently not possible. E. faecalis ATCC 29212 and S. aureus ATCC 29213 were used as quality control strains.
Trimethoprim resistance genotype
The presence of the trimethoprim resistance gene dfrK was confirmed by polymerase chain reaction (PCR) using dfrK-F (5′-GAGAATCCCAGAGGATTGGG-3′) and dfrK-R (5′-CAAGAAGCTTTTCGCTCATAAA-3′) primers and the following conditions: an initial cycle of 94°C for 5 min, followed by 40 cycles of 94°C for 1 min, 58°C for 2 min, and 72°C for 3 min, and a final cycle of 72°C for 7 min. To confirm positive results, sequencing of the dfrK amplicon was carried out for some of the amplicons, and sequences were compared with the dfrK sequences deposited in the databases. 12 To detect the linkage between the genes tet(L) and dfrK, as previously described in S. aureus, 12 a PCR was performed with primers tet(L)-dfrK-F (5′-GATCCCGGATTAGGGAAAAA-3′) and tet(L)-dfrK-R (5′-ATTTGTTCTCCCCCGAAAAT-3′) and all amplicons were sequenced for confirmation. The presence of the other trimethoprim resistance genes previously detected in enterococci, dfrF and dfrG, was also investigated by PCR 3 for all isolates.
Study of the location and genetic environment of the dfrK gene in E. faecium C1723
Southern blot analysis was performed to determine the plasmid or chromosomal location of the dfrK gene in E. faecium isolate C1723. To detect large bacterial plasmids in the presence of genomic DNA, the S1–pulsed-field gel electrophoresis (S1-PFGE) assay was used. Briefly, a small slice of agarose plugs prepared with whole-cell DNA 2 was digested with 14 U of Aspergillus oryzae S1 nuclease (Takara Bio, Inc.) for 45 min. Digested plugs were subjected to PFGE for 16 hr using the following conditions: 6 V/cm2 and 1–30 sec at 14°C. The endonuclease I-CeuI recognizes specific sequences found only in the 23S rRNA genes. As this enzyme digests only bacterial chromosomal DNA, it is also useful to detect the presence of extrachromosomal elements via PFGE and subsequent hybridization. I-CeuI-PFGE (New England Biolabs) digests of enterococcal DNA were performed in agarose plugs for 3 hr at 37°C. Fragments were separated in a 1% (w/v) PFGE agarose gel using the following conditions: 22 hr, 6 V/cm2, and 5–30 sec at 14°C. Subsequently, S1-PFGE and I-CeuI-PFGE digests were transferred to a nylon membrane and hybridization and detection with a dfrK probe, as well as with a 23S rRNA probe in the case of I-CeuI, were done according to the manufacturer's (Roche) recommendations.
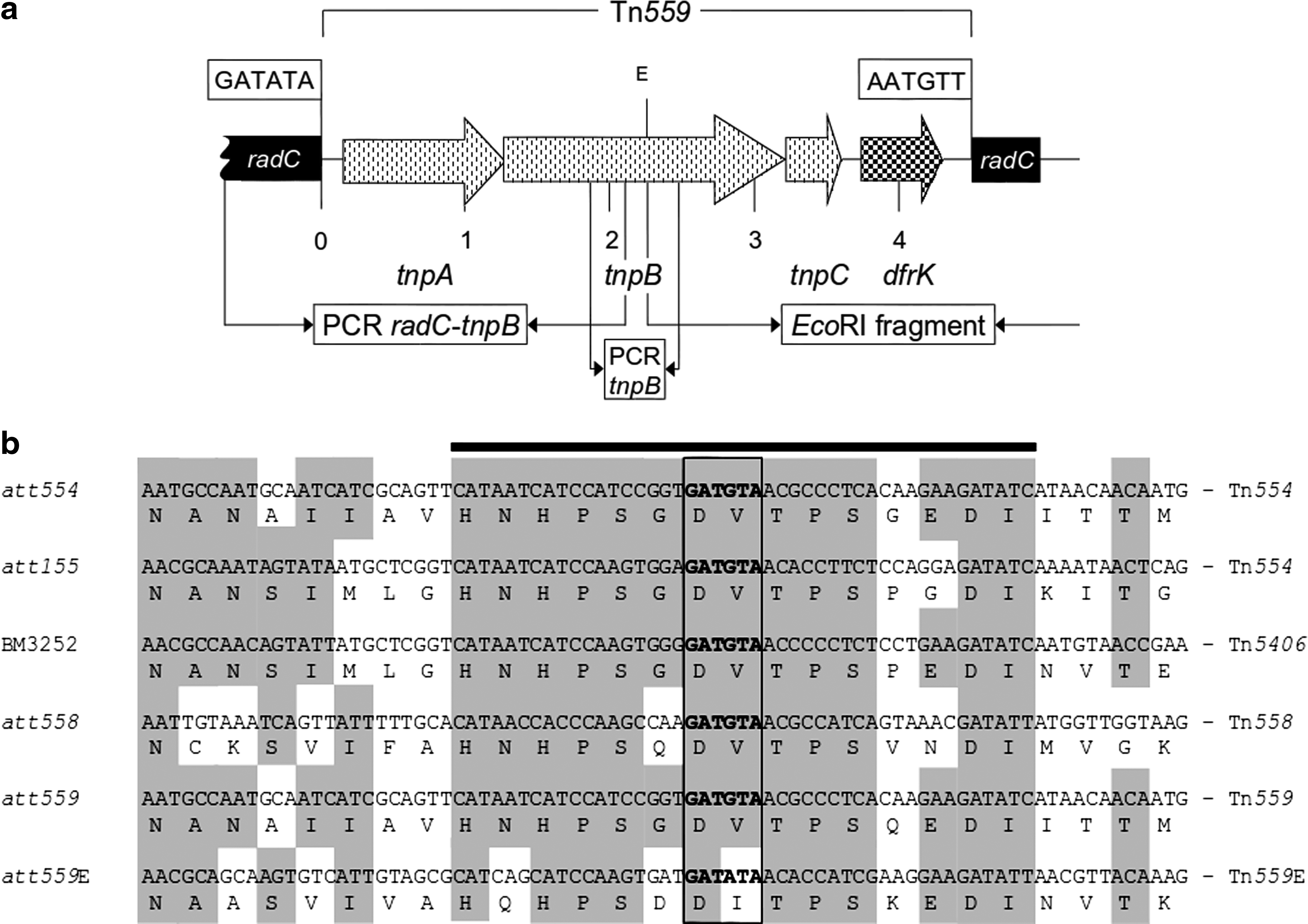
To gain insight into the genetic environment of the dfrK gene in E. faecium C1723, whole-cell DNA was extracted and digested with EcoRI. Resulting fragments were religated (T4 ligase; Fermentas) and a dfrK inverse PCR amplification was carried out. 14 The resulting amplicon was cloned into competent Escherichia coli cells using the Zero Blunt® Topo® PCR cloning kit (Invitrogen). Clones were selected on Luria-Bertani agar plates (Oxoid) supplemented with 50 mg/L kanamycin. Clones were analyzed and the PCR product was sequenced. As EcoRI cuts within the tnpB gene of Tn559, the corresponding reverse PCR amplicons contain only the right-hand end of Tn559 and the downstream-located chromosomal region. The remaining parts of Tn559 including the upstream region were amplified via specific PCR assays using the primer pairs radC_seq (5′-GGAAAGGATGGGGAGAAGAG-3′) with tnpB_seq (5′-TGCTTCAATTTCCACTCTCG-3′) and tnpB-1 (5′-TTGCCCAAAAACTTAAAGAGTTC-3′) with tnpB-2 (5′-TCGTAGCCATATCTTCGGGTA-3′) (Fig. 1a). The corresponding amplicons were completely sequenced using primer walking.
Screening for the transposon Tn559 and its functionality
Specific PCR assays were carried out, with primers derived from the Tn559 sequence, 14 to investigate the enterococcal isolates harboring dfrK for the presence of the Tn554-related dfrK-carrying transposon Tn559. The insertion site of this transposon was also determined. For this purpose, a PCR assay using radC_seq and tnpB-1 primers was performed. As transposition of Tn559 includes the formation of a circular form, inverse PCR using previously described specific primers and conditions 14 were performed to detect this intermediate circular structure in Tn559-positive enterococci.
Results
Trimethoprim resistance
All 166 enterococci included in this study showed resistance to trimethoprim-sulfamethoxazole by the disk diffusion method. However, a wide range of trimethoprim MICs (from ≤0.25 to ≥512 mg/L) was detected among these isolates (Table 1). According to EUCAST breakpoints, strains with MICs of ≥2 mg/L were considered as resistant (considered for uncomplicated UTI infections). Based on these criteria, 29% of the tested enterococci were classified as trimethoprim resistant. In general, higher percentages of trimethoprim-resistant isolates were observed among isolates of the species E. faecium or E. faecalis (40% and 67%, respectively) when compared with isolates of the species E. gallinarum/E. casseliflavus and E. hirae/E. durans (2% and 14%, respectively).
These isolates carry the vanA gene.
MIC, minimum inhibitory concentration.
Detection of the dfrK gene
Despite different trimethoprim MICs, all enterococci included in this study were tested for the presence of the dfrK gene. The dfrK gene was detected in 81 of the 166 isolates (49%). Taking into account the different species, dfrK was detected in 41 of 88 E. faecium (47%), 11 of 15 E. faecalis (73%), 11 of 14 E. hirae/E. durans (79%), and 18 of 49 E. gallinarum/E. casseliflavus (37%) isolates. Moreover, 48% of the vancomycin-resistant isolates (56% of the vanA-, 40% of vanB2-, and 37% of vanC-positive isolates) and 52% of the vancomycin-susceptible isolates harbored the dfrK gene. In addition, 63% and 42% of the trimethoprim-resistant and -susceptible isolates, respectively, carried the dfrK gene (Table 1).
The linkage of dfrK with the tetracycline resistance gene tet(L), previously described in staphylococci, 12 was investigated in the 81 dfrK-positive isolates. This gene linkage was detected in 17 isolates (21%) that belonged to the following species/genotypes: 7 E. faecium/vanA, 1 E. faecium/vanB2, 8 E. gallinarum/vanC1, and 1 E. hirae/vanA (Table 1). All tet(L)-dfrK amplicons and 35 of the remaining 64 dfrK amplicons were sequenced and compared with the database to confirm their identities. These sequences were 100% identical to those described in S. aureus for dfrK and tet(L)-dfrK (GenBank accession number: FN390948).
Characterization of the genetic environment of dfrK
To gain insight into the sequences flanking the dfrK gene in enterococci, the E. faecium strain C1723 was selected for further analysis. This strain showed a trimethoprim MIC of 8 mg/L and harbored the dfrK gene, but was negative for the tet(L)-dfrK linkage. S1-PFGE and I-CeuI-PFGE hybridizations with the dfrK probe revealed the chromosomal location of dfrK gene.
Sequencing of the inverse PCR product of EcoRI digests and the additional Tn559-specific amplicons identified a complete Tn559 element containing the genes tnpA, tnpB, tnpC, and dfrK (Fig. 1a), which differed only by two nucleotides in the noncoding region downstream of dfrK from the previously described staphylococcal Tn559. This transposon was integrated into the radC gene coding for a DNA repair protein. The hexanucleotide 5′-GATATA-3′ was identified at the left-end and the hexanucleotide 5′-AATGTT-3′ at the right-end junction of Tn559. A comparison of the enterococcal integration site att559E within radC with previously described staphylococcal integration sites of Tn559 and structurally related transposons Tn554, Tn5406, and Tn558 is shown in Fig. 1b. Possible functionality of this transposon was also tested by PCR through the detection of circularized intermediates of Tn559. A circular form was detected in this strain, suggesting that the Tn559 element is also functionally active in enterococci.
The presence of Tn559 was investigated in other dfrK-positive enterococci by PCR and 14 additional isolates (11 E. faecium, 1 E. durans, and 2 E. gallinarum) revealed the presence of amplicons indicative of Tn559 (Table 2).
(S), susceptible; (R), resistant.
Presence of other dfr genes
Different combinations of dfrK, dfrF, and dfrG genes were detected among the tested enterococci and a correlation with the MICs of trimethoprim in these isolates can be seen in Table 2. The dfrK gene was simultaneously present with other dfr genes in 51 of the 81 (63%) dfrK-positive isolates.
Discussion
Trimethoprim-resistant bacteria, including enterococci, have been isolated worldwide after the introduction of trimethoprim into clinical use in 1962 or of the combination trimethoprim-sulfamethoxazole in 1969. Although the clinical importance of these antimicrobial agents has gradually declined during the last decades, the lack of oral treatment alternatives for enterococcal infections has led to a renewed interest in them.5,11,30 Nevertheless, only a few previous studies have focused on trimethoprim resistance in enterococci, and none of them in other species than E. faecium or E. faecalis. 3
Phenotypic susceptibility results demonstrated a wide range of trimethoprim MICs (≤0.25 to ≥512 mg/L) in our collection of enterococci, although only E. faecium and E. faecalis showed high-level resistance to trimethoprim (≥512 mg/L). These results are in agreement with previous data on enterococci, wherein a wide range of trimethoprim MICs (0.06–2,048 mg/L) was also detected. 3 However, the percentage of resistance detected in our study was lower than in the study by Cattoir and coworkers (29% vs. 63%). 3 This could be due in part to the different species distributions in both studies and/or also to the different origins of the isolates. The data of the present study also do not show a clear relationship between vancomycin resistance genotypes and dfr genes, although it is interesting to point out that the seven isolates harboring all three dfr genes tested (dfrK+dfrF+dfrG) were vancomycin resistant (two vanB2 and five vanA). In a previous study, it was noted that the dfrF gene was more frequently detected among vanB-containing E. faecium isolates and dfrG gene was more frequently identified among vanA-positive isolates. 3 This finding could not be confirmed in our study, because we detected 46 vanA- and 8 vanB-positive isolates with dfrF and 6 vanA- and 8 vanB-positive isolates harboring dfrG.
In the present study, the dfrK gene was detected for the first time in members of the genus Enterococcus. This gene was detected in trimethoprim-susceptible (42%) and trimethoprim-resistant isolates (63%). The difference in the percentage observed among the two groups might implicate a contribution of this resistance gene to the increase in MIC for trimethoprim. However, its function as a trimethoprim resistance gene in Enterococcus needs to be clarified and future transcriptional studies of this gene in enteroccocci with different trimethoprim susceptibilities would help to understand its role in trimethoprim resistance. Although all sequenced dfrK amplicons exactly corresponded to the staphylococcal dfrK sequence, it remains to be answered which factors play a role in the expression of dfrK in enterococci and may account for its apparent functional inactivity in trimethoprim-susceptible enterococci. In other bacteria, the MIC of trimethoprim increased from 0.25 mg/L to ≥512 mg/L when the dfrK gene was transferred to Escherichia coli 12 and from 0.5 mg/L to ≥512 mg/L when transferred to S. aureus. 13 It is also noteworthy that dfrG, which is next related to dfrK, was detected only in resistant isolates. The dfrG gene was identified in a previous study in E. faecium strains with moderate trimethoprim resistance (2–64 mg/L). 3
The in-depth analysis of the dfrK environment in E. faecium C1723 revealed that the dfrK gene was part of a Tn559 element, which was virtually identical to the one found in S. aureus. 14 Tn559, like Tn554, integrates site-specifically by using an integration site within the radC gene.14,18–20 The hexanucleotide core sequence within this integration site detected in enterococci, 5′-GATATA-3′, is very closely related to the one present in the radC gene from S. aureus, 5′-GATGTA-3′ (Fig. 1b). As E. faecium harbored a radC gene with a suitable integration site, E. faecium may be able to accept resistance-mediating transposons such as Tn554, Tn5406, Tn558, or Tn559 from staphylococci. As all these transposons are nonconjugative, they need to be transferred to an E. faecium host first. Such transfer processes are mainly mediated by plasmids. Although Tn559 has not been yet detected on plasmids, other transposons of the Tn554 family, such as Tn554 and Tn558, have been detected on plasmids.15,20
The observation that the trimethoprim resistance gene dfrG was detected in E. faecium, S. aureus, and S. pseudintermedius in combination with the present finding of dfrK in E. faecium and S. aureus confirms a trimethoprim resistance gene flux between enterococci and staphylococci.5,22,26 Although the dfr genes may not be essential for trimethoprim resistance in enterococci, the presence of dfr genes—particularly as part of a functionally active transposon—may have implications for the further dissemination of these genes to other bacteria that are primarily susceptible to trimethoprim. Enterococci have been previously identified as donors and acceptors of resistance genes and play an important role in the dissemination of resistance genes within and beyond the Gram-positive gene pool.10,21
Footnotes
Acknowledgments
This work was supported in part by Project SAF2009-08570 from the Ministry of Education and Science of Spain and FEDER. M. López received a fellowship from the Gobierno de La Rioja of Spain. The authors thank Kerstin Meyer for excellent technical assistance.
Disclosure Statement
No competing financial interests exist.