
Abstract
Forty-seven vancomycin-resistant Enterococcus (VRE) strains were isolated from clinical samples in 13 Zhejiang hospitals and fecal samples from ICU patients in a large teaching hospital in China. No VRE isolates were detected in healthy human subjects. CC17 was the main clonal complex in clinical Enterococcus faecium isolates but not in isolates from healthy human subjects. Novel vancomycin-resistance transposons were detected among VRE strains. This is the first report demonstrating insertion of tnpA and fosB genes in the vanRS–vanH intergenic region of Tn1546 leading to coresistance to vancomycin and fosfomycin. The four plasmid replicon types (pRUM, pRE25, pEF418, and pB82) were more common in VRE isolates, suggesting their association with vancomycin resistance and nosocomial transmission. The prevalence rate of vancomycin-resistant Staphylococcus aureus–related Inc18-like plasmid, pIP501, in VRE was 21.3%. The prevalence of the esp gene among VRE isolates was high (76.6%). In several VRE strains, the esp and hyl genes were cotransferred with the vanA gene by conjugation. Although the frequency of VRE is low in Chinese hospitals, its association with virulence determinants, the vancomycin-resistance transposon with other resistance gene insertions or plasmids may lead to multidrug resistance and the evolution of pathogenic VRE.
Introduction
VRE has become an important nosocomial pathogen in many countries but it is rare in mainland China, although glycopeptide antimicrobials have been used there for decades. 22 However, recently, VRE colonization and infection rates in several hospitals in China have increased with frequencies of up to 10%.12,25 Horizontal transfer of a resistance gene cluster has also been regarded as an important contributor to this phenomenon. It is known that the vanA gene cluster is part of Tn1546-like elements, and this indicates that the horizontal transfer of Tn1546-like elements plays an important role in the spread of vanA-type VRE17,20 and other species, such as vancomycin-resistant Staphylococcus aureus (VRSA). VRSA is thought to arise by in vivo transfer of an Inc18-like vanA plasmid from VRE to S. aureus.23,24 Currently, VRSA infections are rare. If VRE with Inc18-like vanA plasmids becomes more common, then this will lead to an increase in the rate of serious clinical infections. DNA polymorphisms among Tn1546 variants and the relevant plasmid types can be combined with epidemiological data and exploited for tracing the transmission routes of vancomycin-resistance genes.
The purpose of this study was to characterize the clonal diversity of Enterococci and the incidence of antibiotic resistance, resistance transmission, and virulence factors in Enterococci isolated from clinical samples and human feces.
Materials and Methods
Bacterial isolate collection
A total of 47 VRE strains were isolated from the following sources: (i) 40 VRE isolates were screened from the 370 clinical strains from clinical samples from patients in 13 hospitals in different cities in Zhejiang Province from January to December 2009. Of these 40 VRE strains, 38 strains were Enterococcus faecium, one was Enterococcus faecalis, and one was Enterococcus avium. These VRE strains were isolated from blood (11 strains), urine (16 strains), ascites (7 strains), and feces from patients with diarrhea (6 strains). (ii) Seven VRE isolates identified as E. faecium were screened from 350 fecal samples from hospitalized patients in the ICU in the First Affiliated Hospital of Zhejiang University Medical College from January to June of 2009. (iii) No VRE isolate was screened from 500 fecal samples from healthy human subjects who had visited the Department of Health Examination in the above hospital from January to December 2009.
The 59 clinical vancomycin-susceptible Enterococcus (VSE) isolates (37 E. faecium isolates and 22 E. faecalis isolates), and 38 VSE isolates (11 E. faecium isolates and 27 E. faecalis isolates) were derived from fecal samples obtained from healthy human subjects who were selected at random for comparative study.
Antibiotic resistance
The minimum inhibitory concentrations (MICs) of glycopeptides (vancomycin and teicoplanin) were determined by Etest (AB Biodisk, Solna, Sweden) and susceptibility to ampicillin, gentamicin (for high-level gentamicin resistance), fosfomycin, tigecycline, rifampin, quinupristin-dalfopristin, and linezolid was determined by the Kirby Bauer (K-B) method. E. faecalis ATCC 29212 was used as a control. The results were interpreted according to Clinical Laboratory Standards Institute (CLSI 2010).
Multilocus sequence typing
Multilocus sequence typing (MLST) was performed as described by Homan et al. and Ruiz-Garbajosa et al.5,15 Sequence types (STs) were assigned using http://efaecium.mlst.net/ and http://efaecalis.mlst.net/. eBURST, available at http://linux.mlst.net/burst.htm, was used to analyze MLST data.
Polymerase chain reaction amplification of virulence factors
Total DNA was extracted as previously described 19 and used as template in polymerase chain reactions (PCRs) to detect the presence of genes involved in the expression of aggregation substance (agg), gelatinase (gel), enterococcal surface protein (esp), cytolysin/hemolysin (cyl), and a protein with homology to hyaluronidases (hyl).
DNA sequence analysis of VRE-resistance determinants and transposons
The vanA, vanB, vanC1, and vanC2, 3 genes were detected in VRE isolates by PCR as previously described. 2 Tn1546-like elements harboring vanA in these isolates were more fully characterized by overlapping PCR and nucleotide sequencing, also as previously described. 18
Conjugation
Filter matings were performed using the VRE strains as donors and E. faecium BM4105RF (resistant to rifampin and fucidin) as the recipient strain as previously described. 16 Transconjugants were selected on BHI agar plates containing vancomycin (32 μg/ml) and rifampin (256 μg/ml).
Plasmid analyses
PCR assays were optimized to amplify 10 characterized Enterococcal plasmid replicon groups. 7 The replicon grouping (pS86, pRUM, pMBB1, pCF10, pAM373, pRE25, pEF418, pIP501, Pb82, and pHTβ) is based on phylogenetic relationships among the replication genes (repA) of Enterococcal plasmids.
Statistical analyses
The statistical significance of differences in the prevalence of plasmid replicon types between VRE and VSE isolates was calculated by the Pearson test, Chi-squared test, or Fisher's exact test using SPSS 15.0 (SPSS Institute, Chicago, IL). A p-value less than 0.05 was considered statistically significant.
Results
Antimicrobial resistance
The 47 VRE isolates showed a high degree of resistance (MIC 256 μg/ml) to vancomycin and varying degrees of resistance to teicoplanin (MIC 16–256 μg/ml). Five vancomycin-resistant E. faecium (VREF) isolates showed resistance to quinupristin-dalfopristin. All of the 47 VRE isolates showed susceptibility to tigecycline and linezolid. The resistance rates of VRE to other antibiotics including ampicillin, rifampin, high-level gentamicin, and fosfomycin were 97.9%, 87.2%, 74.5%, and 57.4%, respectively.
Multilocus sequence typing
The E. faecium MLST data in this study showed that the 45 VREF isolates had 14 STs. The predominant ST78 (19 strains) belonged to the clonal complex CC17, and 27 of the 37 vancomycin-susceptible E. faecium (VSEF) isolates also belonged to CC17. The 11 VSEF isolates from healthy human fecal samples showed different STs not belonging to CC17 (Fig. 1).
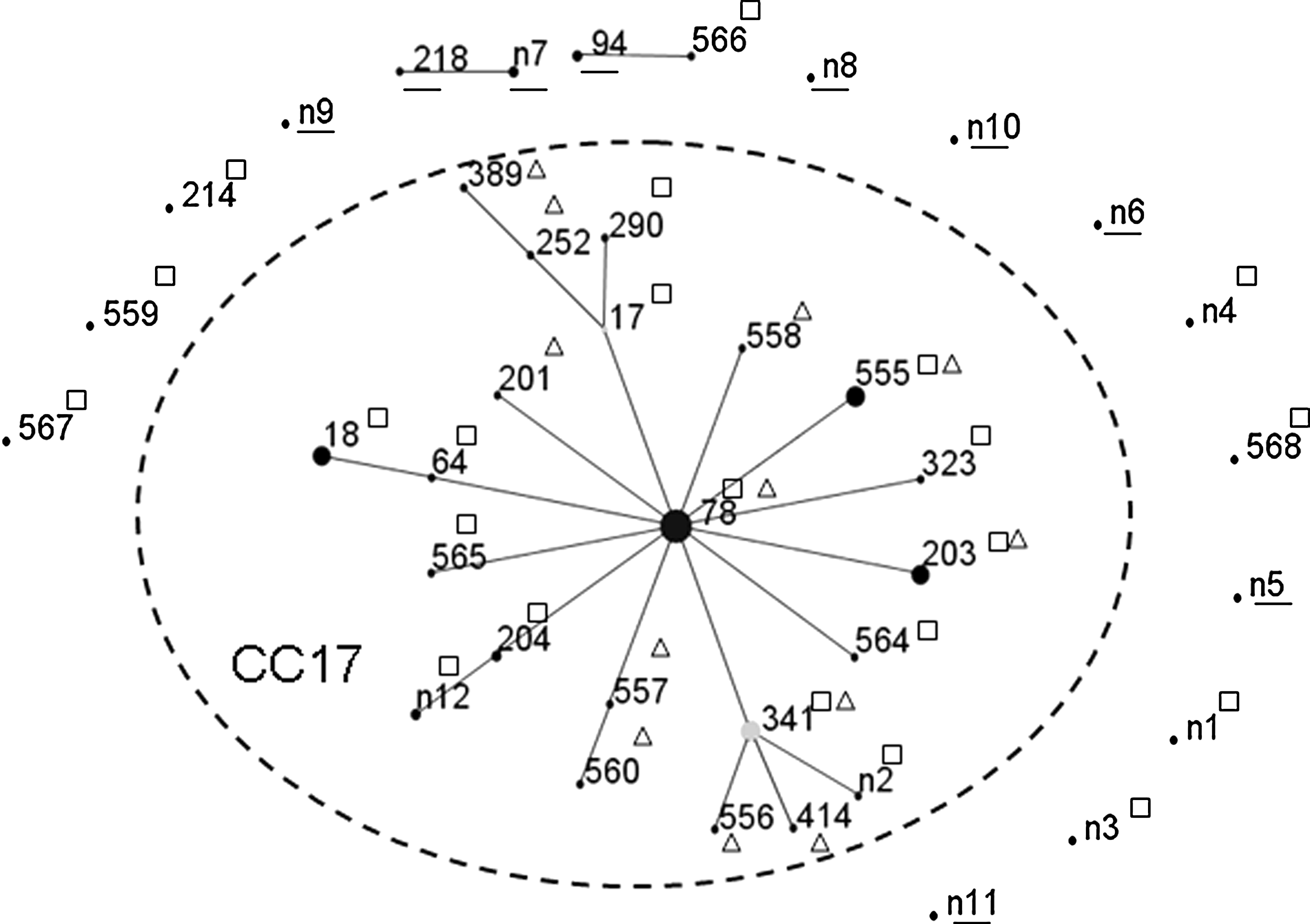
Population snapshot of Enterococcus faecium derived from this study by the eBURST algorithm. The radial diagram reflects the predicted evolutionary descent from the founder ST. A circle represents an ST, and its size corresponds to the number of isolates. The number indicated by a single underline represents STs of VSEF strains from healthy human feces, the number indicated by “□” represents clinical VSEF strains, and the number indicated by “Δ” represents clinical vancomycin-resistant E. faecium strains. The broken line indicates CC17. VSEF, vancomycin-susceptible E. faecium; STs, sequence types.
The E. faecalis MLST results showed that the only VRE E. faecalis was ST4 (CC4). Of the 22 VSE E. faecalis clinical isolates, CC16 (11 strains) and CC4 (7 strains) were the main STs. The E. faecalis strains from healthy human subjects showed separate STs.
Incidence of genes encoding for potential virulence factors
The prevalence rates of the virulence genes (esp, hyl, cyl, agg, and gel) among 47 VRE strains were 76.6% (36 strains), 21.3% (10 strains), 6.4% (3 strains), 6.4% (3 strains), and 4.3% (2 strains), respectively. Furthermore, multiple virulence genes coexisted in the Enterococci isolates.
VRE-resistance determinant and transposon
The vanA gene of the glycopeptide-resistance determinant was found in all of the 47 VRE isolates. Three types of Tn1546-like elements were found by overlapping PCR and sequences were compared with previously published variations in Tn1546 (Fig. 2). In total, 22 of 47 VRE isolates possessed the Tn1546-like element with IS1485 in the vanX–vanY intergenic region as described in our previous report (GenBank EU599211).12,13 Ten VRE isolates possessed the unique molecular variations characteristic of a Tn1546-like element, with tnpA and fosB (involved in the fosfomycin resistance) genes inserted in the vanRS–vanH intergenic region (Fig. 2). The locations of the insertions in this region were identical, corresponding to nucleotide 5813 bp of Tn1546 (GenBank M97297) with an 8-bp duplication of the target sequence (ATAATTTT). These nucleotide sequences for the Tn1546-like element were submitted to GenBank (GenBank JN982328). The other 15 VRE isolates possessed the typical Tn1546 element.
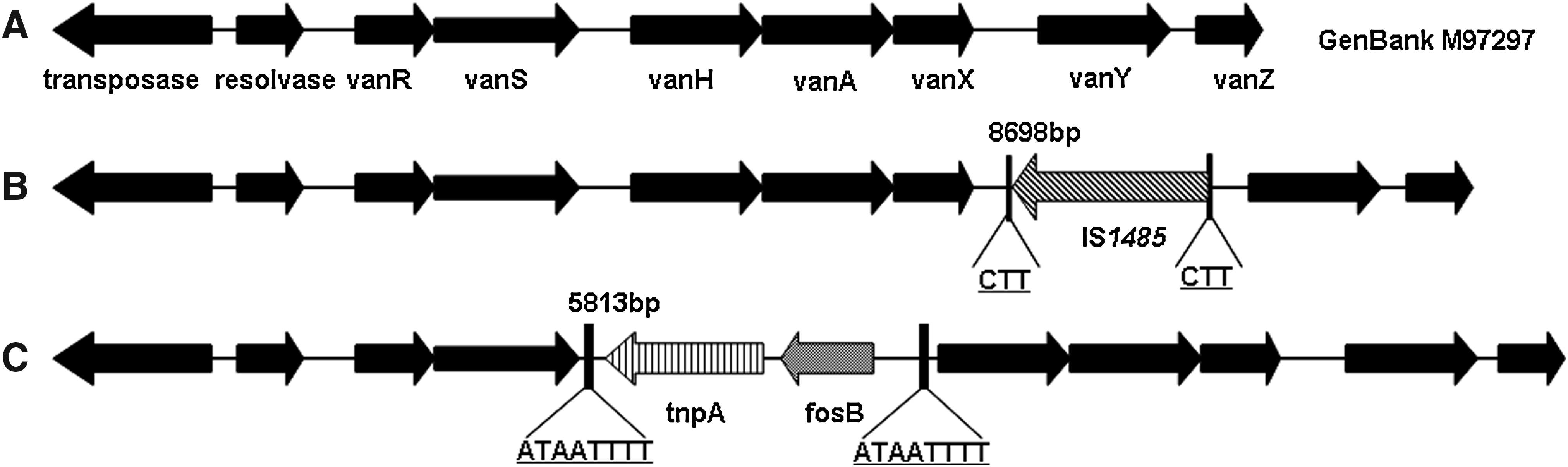
Genetic maps of Tn1546 types of E. faecium isolates in this study. The open reading frames and the direction of transcription are marked by boxes with arrows.
Transfer of resistance and virulence genes
Vancomycin resistance was transferred by filter matings at rates of 10−5–10−7/donor CFU from 31 of the 47 donor strains. All transconjugants expressed resistance to vancomycin and teicoplanin at an MIC that was the same as or lower than that for the donor strains. These 31 conjugative VRE strains were vanA positive. The esp and hyl genes were cotransferred with the vanA gene by conjugation in three VRE strains and one VRE strain, respectively.
Occurrence and distribution of plasmid replicon types
The distribution of the 10 plasmid replicon types in VRE and VSE isolates from clinical samples and human feces is shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertonline.com/mdr). Among the 47 VRE isolates, a total of 46 (97.9%) strains were PCR positive for one to four plasmid replicon groups in this study while pAM373, pMBB1, and pHTβ were not observed. Among the 31 VRE conjugants, replicon types pB82 (67.7%), pEF418 (64.5%), pRUM (38.7%), pRE25 (38.7%), and pIP501 (3.2%) were observed. Five VRE conjugants were negative for the 10 plasmid replicon groups.
The prevalence of the four replicon plasmid types (pRUM, pRE25, pEF418, and pB82) was higher in the VRE isolates compared with the VSE isolates (p<0.05) and was also higher in clinical VSE isolates compared with healthy human fecal VSE isolates (p<0.05). The prevalence of the above four plasmids was higher in CC17 E. faecium strains compared with non-CC17 E. faecium strains and was also higher in the CC17 VREF strains than the CC17 VSEF strains.
The PCR products of the rep amplicons of pS86, pRUM, pCF10, pRE25, pEF418, pIP501, and Pb82 were further identified by DNA sequence analyses. Replicons were identical to the reference sequences.
Discussion
In this study, the 47 VRE strains screened were isolated from clinical samples from hospitalized patients in 13 Zhejiang hospitals and from fecal samples from ICU patients in a large teaching hospital in China. No VRE isolates were detected in feces from healthy human subjects. The occurrence of VRE in China is suggested to occur mainly in clinical environments, which is likely to be the result of antibiotic use in hospitals and clinics. In Europe, the high rate of VRE is mainly due to the use of avoparcin. Moreover, 95.7% of the VRE isolates were E. faecium. We observed high resistance rates of VRE to other antibiotics including ampicillin, rifampin, high-level gentamicin, and fosfomycin. Tigecycline and linezolid were effective against all Enterococci strains investigated.
In this study, MLST profiles showed that all 45 VREF isolates belonged to one clonal complex, CC17. CC17 was also the main CC in clinical VSEF isolates but not in VSEF isolates from healthy humans, suggesting CC17 confers a selective advantage and facilitates adaption in the clinical environment. E. faecium CC17—which is characterized by ampicillin resistance, a pathogenicity island (containing esp), and an association with hospital outbreaks—has disseminated worldwide. 21 Our results also support the hypothesis that Enterococci gains a selective advantage and is able to exploit the clinical environment through acquisition of multiple adaptive mechanisms, including the sequential acquisition of resistance genes (such as vanA) and putative virulence genes (such as esp and hyl), which is facilitated by horizontal gene transfer and recombination ST17 (the presumed founder of CC17). 9 The spread of CC17 in China will have serious clinic consequences, especially if it acquires resistance to glycopeptides. Two clonal complexes, CC16 and CC4, were the main CCs of clinical E. faecalis isolates. As was the case for E. faecium, this finding suggested that an adaptation to the clinical environment had occurred in E. faecalis.
This study also explored the dissemination of the virulence markers gel, agg, cyl, hyl, and esp among VRE isolates. The prevalence of the esp gene in VRE isolates was high. It has been reported that esp was an important virulence factor in Enterococcus infection. The hyl gene is described as part of a genomic island and was recently identified on a plasmid that increased the ability of Enterococci to colonize the gastrointestinal tract.1,8 As previously reported, vancomycin-resistance genes cotransferred with the hyl-containing plasmid and the gene cluster, suggesting that these elements maybe physically linked.1,8 In this study, the esp and hyl genes cotransferred with the vanA gene by conjugation in a plasmid or different plasmids in several VRE strains. This phenomenon has seldom been reported. 1
Vancomycin resistance disseminates not only clonally but also by horizontal transfer of resistance elements to different clones of Enterococci. VRE isolates were confirmed to be vanA positive. Identical Tn1546-like types were found among the vancomycin-resistant isolates, suggesting the possibility of horizontal transmission of the vancomycin-resistance transposon. In this study, IS1485 was detected between the vanX and vanY genes in the 22 clinical VRE isolates, as we previously reported. 12 Additionally, this is the first study to identify the presence of the tnpA and fosB genes in the vanRS–vanH intergenic region of the vanA gene cluster among the 10 VRE isolates, in contrast to reports based upon studies conducted in other countries.4,6,11,18 Fosfomycin is often the antibiotic of choice to treat VRE infection in clinics. However, the insertion of the fosB gene into Tn1546 will lead to coresistance to vancomycin and fosfomycin. The dissemination of this Tn1546-like element and the insertion of antibiotic-resistance genes will have serious clinical consequences.
PCR-based replicon typing, linked to the detection of other important plasmid-encoded traits, seems to be a feasible tool for tracing disseminating resistance plasmids. 7 The four replicon plasmid types (pRUM, pRE25, pEF418, and pB82) were more common in VRE than VSE even within the same clonal complex (CC17) and also in clinical isolates compared with fecal isolates from healthy human subjects. The evidence presented in this report showed that plasmids pRUM, pRE25, pEF418, and pB82 were associated with the acquisition of vancomycin resistance and nosocomial transmission. However, our results also suggested that pAM373, pMBB1, and pHTβ were not prevalent in the Enterococci in this study. In five VRE conjugants, none of the 10 plasmid replicon groups were observed. It was suggested that there was the genetic linkage of vanA to unknown transferable replicons in this study. Inc18 incompatibility plasmids are a family of broad-host-range conjugative plasmids that occur naturally in Enterococcus and Streptococcus spp. A previous report suggested that the Inc18-like vanA plasmid may be more likely than other vanA plasmids to transfer from Enterococcus spp. to S. aureus and that this plasmid was associated with VRSA. 23 The pIP501 replicate belongs to the Inc18 group of plasmids. 7 The pIP501-related broad-host-range plasmids were involved in the spread of high level VanA-type glycopeptide resistance, as described previously. 14 In this study, the prevalence rate of the pIP501 plasmid in VRE had no significant difference from that in VSE (21.3% vs. 16.9%). Only one VRE conjugant was detected in our study that contained the pIP501 plasmid. These findings may be useful in predicting future VRSA occurrence.
In summary, CC17 was the main clonal complex in clinical E. faecium isolates, especially VREF strains, in Chinese hospitals. Our findings suggested that identical Tn1546-like elements harboring the vanA gene and the four plasmid replicon types (pRUM, pRE25, pEF418, and pB82) were associated with vancomycin resistance and nosocomial transmission. The main virulence genes in VREF, esp and hyl, were occasionally cotransferred with the vanA gene. Although the frequency of VRE is low in clinics, its association with virulence determinants, clonal spread, and the vancomycin-resistance transposon or plasmids may lead to the evolution of pathogenic VRE and serious clinical consequences.
Footnotes
Acknowledgments
This work was supported by the research grant from the National Natural Science Foundation of China (no. NSFC30800035), the Ministry of Health of the People's Republic of China (no. 201002021), and the Education Department Foundation of Zhejiang Province (no. Y200805344).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.