
Abstract
Flavobacterium psychrophilum is the causative agent of the recognized diseases ‘bacterial coldwater disease’ and ‘rainbow trout fry syndrome’ and is found in many farmed freshwater and marine fish species. In Norway, the bacterium has mainly been isolated from Atlantic salmon (Salmo salar L.) and brown trout (Salmo trutta L.). In the present study, 26 isolates from Norwegian farmed salmonids were examined. All isolates were tested for susceptibility towards various antibacterial drugs by the disk diffusion method, and minimum inhibitory concentration values for oxolinic acid and flumequine were established for selected isolates. All isolates from rainbow trout displayed reduced susceptibility towards quinolones, while brown trout and Atlantic salmon isolates were susceptible. The quinolone resistance determining regions (QRDRs) of the gyrA, gyrB, parC, and parE genes were sequenced. Sequence analysis of the QRDR of gyrA in quinolone resistant isolates revealed a threonine:arginine amino acid substitution at position 82 in all 16 isolates from Norwegian rainbow trout and a single reference strain isolated from rainbow trout in Sweden. No evidence for plasmid-mediated quinolone resistance was found in any of the isolates. Pulsed-field gel electrophoresis and phylogenetic analysis of parC and gyrB sequences indicate a clonal relationship between rainbow trout isolates.
Introduction
Quinolones such as oxolinic acid and flumequine have been used in Norwegian aquaculture. 18 Oxolinic acid is a synthetic first-generation quinolone that is an authorized veterinary medicine for cattle, fish, and poultry, while flumequine is a first-generation synthetic fluoroquinolone introduced to treat infections in cattle, swine, poultry, and fish. Flumequine has been reported to be more effective than oxolinic acid, as it is readily absorbed and distributed to various body tissues of domestic animals and fish.18,23
The most common mechanism of bacterial quinolone resistance (QR) is alteration of the quinolone target enzymes: DNA gyrase and topoisomerase IV. Another mechanism utilized by Gram-negative bacteria is reduction of outer membrane permeability to quinolones through decreased expression of porins. Both mechanisms are chromosomally mediated.10,25 DNA gyrase and topoisomerase IV are each composed of two subunits encoded by gyrA/gyrB and parC/parE, respectively. Mutations resulting in amino acid substitutions between positions 67 and 106 of the GyrA protein have been found to be most commonly responsible for resistance against quinolones. This region is regarded as the gyrA quinolone resistance determining region (QRDR).25,35 The point mutations causing amino acid replacement in either of the QRDRs in the two subunits of the target enzymes GyrA and ParC are widespread in many bacteria. The presence of a single mutation in the QRDR of gyrA is usually responsible for resistance to first generation antibiotics, but for a high level of resistance against fluoroquinolones, an additional mutation is usually required in gyrA or in any other target of quinolones such as gyrB or parC. 5
Plasmid-borne qnr was first discovered in 1998 in Klebsiella pneumoniae 17 and, subsequently, in many other bacteria.14,16,17 Three families of qnr genes (qnrA, qnrB, and qnrS) have been reported differing in nucleotide identity by 40% or more. 12 In Scandinavia, qnrA and qnrS were first reported in Denmark with qnrS subsequently reported in Norway. 14
Salmonid farming is an internationally important industry, and Norway is one of the largest producers of these cold water fish species. While isolations of F. psychrophilum have been previously related and limited to fin rot and ulcers in brown trout and Atlantic salmon raised for re-stocking purposes 13 , the number of outbreaks related to disease in Norwegian farmed rainbow trout has increased in recent years. 21
The quinolones oxolinic acid and flumequine currently represent two of the few antimicrobials licensed for use in aquaculture in Norway. The aim of the current study was, therefore, to investigate the genetic basis for the detected QR in F. psychrophilum, in Norway by characterization of the QRDRs of DNA gyrase and topoisomerase IV, and also, in addition, to investigate for the presence of qnr genes.
Materials and Methods
A total of 26 bacterial isolates of F. psychrophilum from diseased fish farmed in Norway were examined. Of these, 16 were from farmed rainbow trout; 6, from farmed Atlantic salmon; and 4, from farmed brown trout. One reference isolate from farmed rainbow trout from a farm in Sweden and the type strain NCIMB 1947T (Table 1) were also included. Escherichia coli Lo qnrA1, K. pneumoniae 15, qnrB1, and Enterobacter cloacae 287, qnrS1 (kindly provided by Dr. L. Poirel, Centre Hospitalier Universitaire De Bicêtre France) were used as qnr positive controls.
Median in mm.
No. of strains screened for gyrB=12 (6 RT, 3 SS, 2 BT, NCIMB 1947T); parC=12 (7 RT, 3 SS, 1 BT, NCIMB 1947T); parE=13 (7 RT, 3 SS, 2 BT, NCIMB 1947T).
No. of strains tested for MIC=09 (5 RT, 2 SS, 1 BT, NCIMB 1947T).
RT, rainbow trout; BT, brown trout; SS, Atlantic salmon; W, wild; P, position; AS, amino acid substitution; SM, silent mutation; MIC, minimum inhibitory concentration.
Antimicrobial susceptibility testing
Antibiotic susceptibility was evaluated for all isolates by the disk diffusion method on dilute Mueller-Hinton agar supplemented with 5% fetal calf serum (Sigma Chemical) 4 and on Anacker and Ordal agar (AOA). 1 Disks containing trimethoprim+sulfamethoxazole 5.2+240 μg, flumequine 30 μg, oxolinic acid 10 μg, tetracycline 80 μg, and florfenicol 30 μg (Neo–Sensitabs Rosco® Taastrup, Denmark) were used. The inoculated plates were incubated at 15°C for 3–6 days.6,9 Aeromonas salmonicida subsp. salmonicida ATCC 14174T was included as quality control. The tests were performed in duplicate.
Minimum inhibitory concentration determination
The minimum inhibitory concentrations (MICs) of oxolinic acid and flumequine were determined by the broth micro-dilution method. The stock solutions were made according to the CLSI guidelines 4 and the manufacturer's instructions (Sigma-Aldrich). Stock solutions were further diluted in the micro-titer assay with Anacker and Ordal broth to concentrations from 8 to 0.01562 μg/ml for flumequine and from 16 to 0.03125 μg/ml for oxolinic acid. 21 For each antimicrobial dilution, two replicate wells were inoculated with bacterial suspension and incubated at 15°C for 24–48 h. 4
DNA extraction, polymerase chain reaction amplification, and sequence analysis
DNA was obtained by suspending two colonies of bacterium in 50 μl sterile distilled water. This was incubated at 100°C for 15 min and then centrifuged for 2 min at 14000 rpm. The supernatant was used as DNA template.
The QRDRs were amplified using the primers and annealing temperatures listed in Table 2. Polymerase chain reaction (PCR) amplification was performed in a total reaction volume of 50 μl. The reaction mixture contained 5 μl of DNA, 1.25 μl of dNTP, 5 μl of 10× key buffer (15 mM MgCl2), 1.25 μl of 50 mM magnesium chloride (MgCl2), 25 pmol of each primer, and 3.1 U of Taq polymerase. The PCR products were purified for sequencing using the QIAquick PCR purification kit and were sequenced by GATC Biotech AG, Jakob-Stadler-Platz 7, D-78467 Konstanz. Sequence identities were confirmed using the NCBI blast program: http://blast.ncbi.nlm.nih.gov/blast
Each polymerase chain reaction amplification had initial denaturation at 94°C for 5 min, followed by 30 cycles of denaturation, annealing and extension, and final extension at 72°C for 5 min.
Phylogenetic analysis
gyrB and parC gene sequences were subjected to phylogenetic analysis. Following alignment in ClustalX 32 , neighbor-joining analysis was performed in PAUP 4.0 using the kimura 2 parameter model. 31 Missing bases were excluded, and bootstrap confidence values were obtained with 1,000 re-samplings.
Preparation of bacterial DNA for pulsed-field gel electrophoresis
Cultures incubated for 3 days on AOA at 15°C were scraped off the agar surface and re-suspended in buffer (100 mM Tris: 100 mM EDTA pH 8, 0). The optical density was then measured (610 nm) and adjusted to 1.3. A 400 μl volume was then mixed with 20 μl proteinase K (20 mg/ml) (Invitrogen) and an equal volume of 1% agarose gel (SeaKem® Gold, Cambrex Karlskoga, Sweden):1% SDA in TE-buffer (10 mM EDTA, pH 8.0) and dispensed into a plug mould (Bio-Rad). The plugs were allowed to sit on ice for 10 min, before transfer to a clean 1.5 mL Eppendorf tube containing 5 mL lysis buffer (50 mM Tris pH 8.0, 50 mM EDTA, 1% sarkosyl, 25 μl proteinase K (20 mg/ml)). After incubation at 54°C for 2 h, the lysis buffer was removed, and the plugs were washed with 3× TE-buffer followed by incubation in 15 min at 50°C. The plugs were subsequently stored at 4°C until use.
Pulsed-field gel electrophoresis
A small (∼2×7 mm) portion of the plug was removed and incubated in a 1.5 mL tube containing the enzyme buffer for 1 h at 37°C, after which the buffer was replaced by 125 μl fresh buffer containing 20 U of enzyme, and incubated for 3 h at 37°C. The restriction enzymes KpnI and XhoI (both New England Biolabs) were used.
Restriction fragments were subjected to electrophoresis in 1% agarose (SeaKem®) gels in 0.5% tris-borate-EDTA buffer using a CHEF-DR III system (Bio-Rad). Running conditions were 6 v/cm 0.5 to 7 s ramp for 19 h at 12°C for the KpnI and 1–12 s for 18.5 h at 12°C for XhoI.
After electrophoresis, the gels were stained in ethidium bromide (2 μg mL−1) for 20 min, de-stained in water for 15 min, and photographed under uv-transillumination (GeneGenius, Syngene Cambridge, UK).
Results
All F. psychrophilum isolates from rainbow trout examined in the present study displayed reduced susceptibility towards quinolones (oxolinic acid and flumequine), while isolates from brown trout and Atlantic salmon were susceptible. However, all isolates were tetracycline, sulfamethoxazole/trimethoprim, and florfenicol susceptible. The quinolone-resistant isolates of F. psychrophilum from rainbow trout yielded disk diffusion inhibition zones ranging between 14 and 46 mm for oxolinic acid and between 28 and 60 mm for flumequine, while for susceptible isolates from brown trout and Atlantic salmon, this range was 54–90 mm for oxolinic acid and 74–100 mm for flumequine. The MICs ranges were 3–6 μg/ml for oxolinic acid and 2–4 μg/ml for flumequine in the rainbow trout isolates (Table 1). On phylogenetic comparison of gyrB and parC sequences, all rainbow trout isolates revealed identical sequences, while isolates from brown trout and salmon were different and quite heterogeneous (Fig. 1a, 1b). The sequence of the QRDR of gyrA of the quinolone-resistant isolates demonstrated a single point mutation in the DNA sequence, resulting in substitution of C for G at bp 245, resulting in the substitution of threonine by arginine at position 82 in all 16 isolates from rainbow trout. The Swedish rainbow trout isolate also displayed the same amino acid substitution. All resistant isolates also displayed silent mutations. Identical QRDR sequences were identified from resistant and susceptible strains in gyrB, parC, or parE, although 5 different silent mutations were observed in parC, 10 in gyrB, and 4 in parE. All sequences obtained have been submitted to Gene Bank (Table 3). The plasmid mediated qnr genes (A, B and S) were not found in any of the isolates.
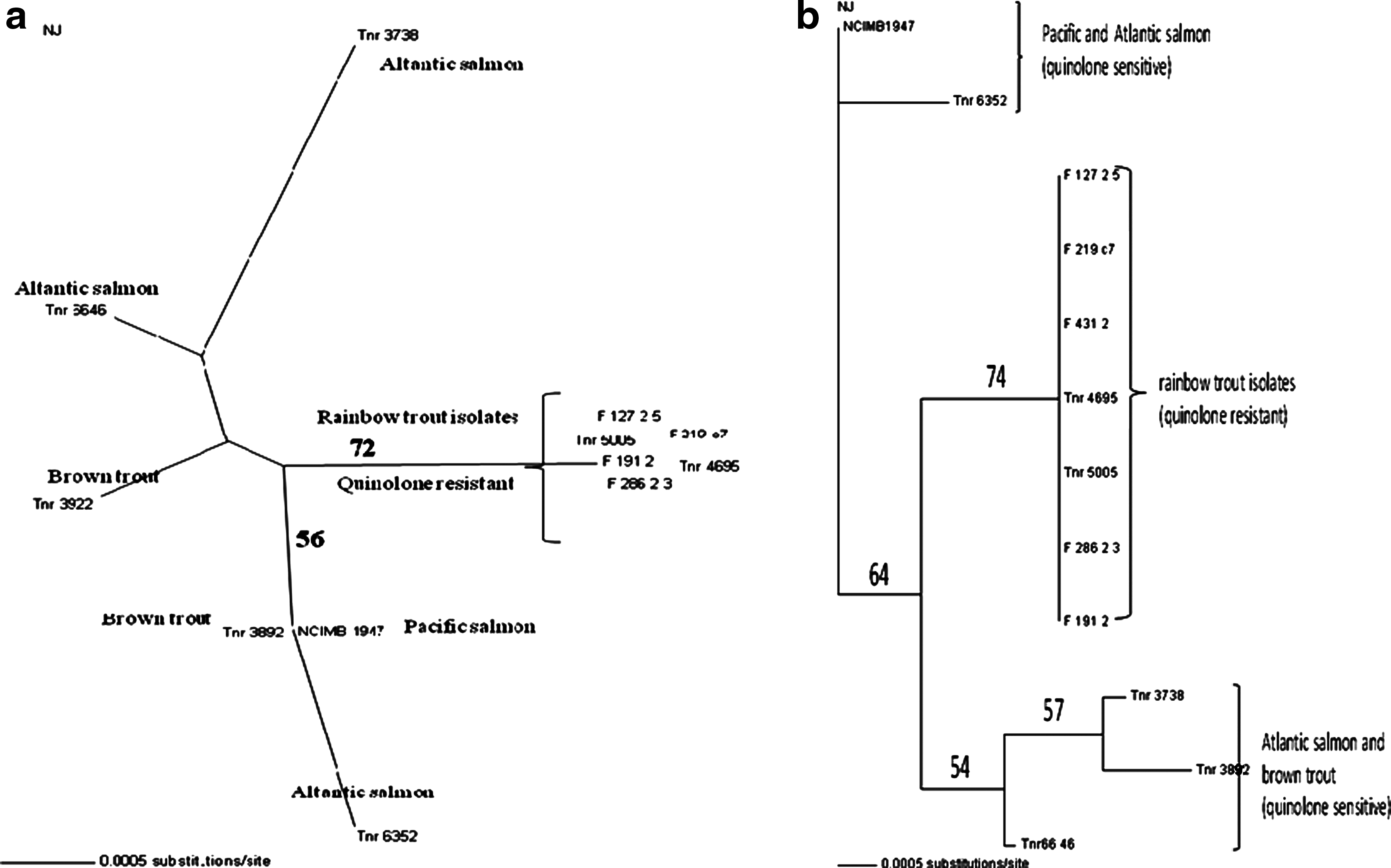
Phylogenetic analysis of
Pulsed-field gel electrophoresis (PFGE) band patterns using restriction enzymes KpnI and XhoI are shown in Figs. 2 and 3. The patterns obtained from isolates of F. psychrophilum from rainbow trout isolated on the western coast of Norway in 2008 are highly similar using both these enzymes, while other strains from rainbow trout in Norway obtained in 2004 and 2007 show a slightly different yet highly related pattern. Strains from brown trout and salmon show patterns with a high degree of diversity and separate from the rainbow trout strains.
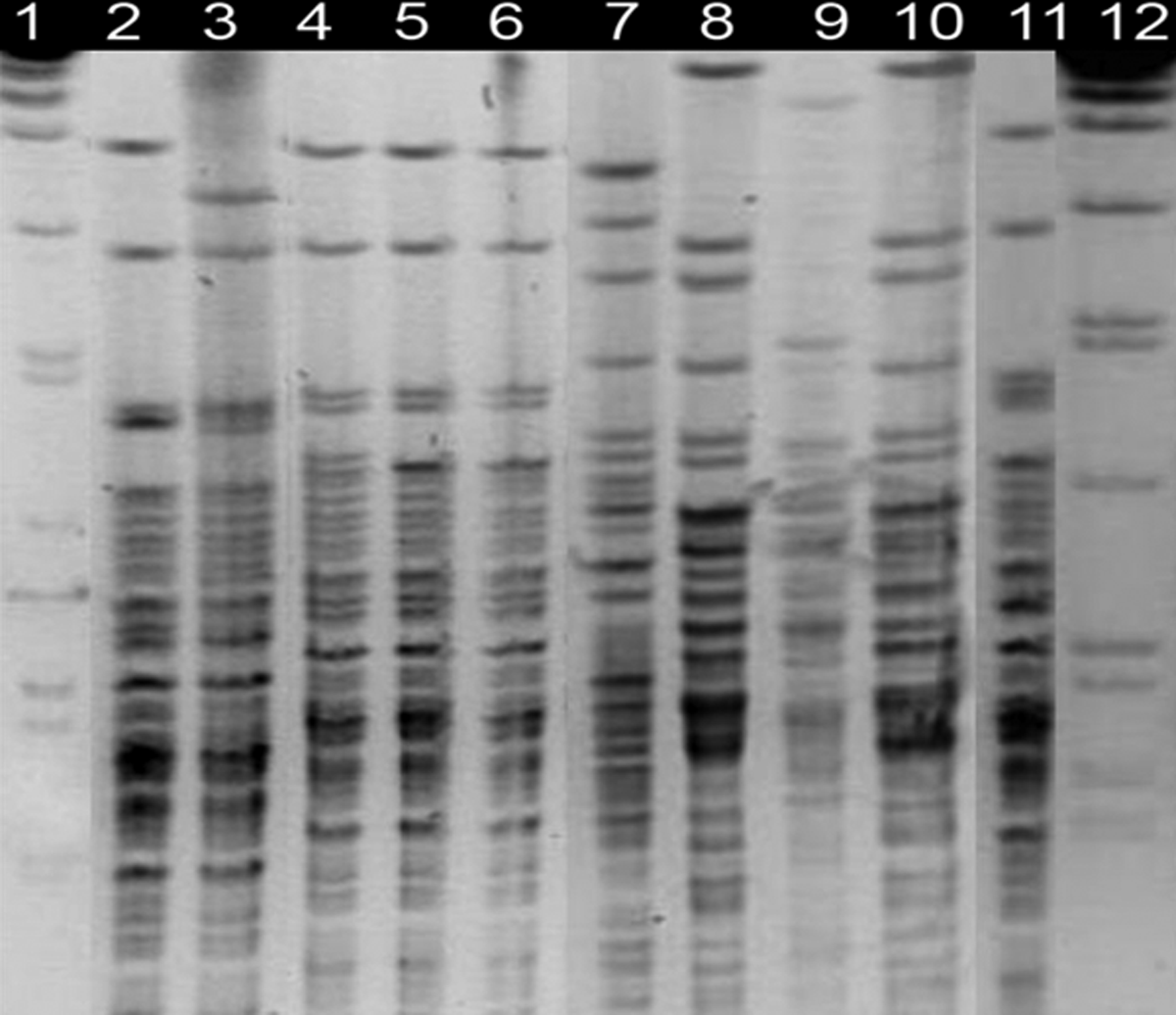
Strains of F. psychrophilum digested with restriction enzyme KpnI. Lane 1–12. Salmonella braenderup H9812, F.nr 5005, F.286-2-3, F.127-2-5, F.165-1-6, F.219-C7, F.251-1-6, F.nr 4115, F.nr 3922, F.nr 3987, F.nr 5049, Salmonella braenderup H9812.

Strains of F. psychrophilum digested with restriction enzyme XhoI. Lane 1–12. Salmonella braenderup H9812, F.nr 5005, F.286-2-3, F.127-2-5, F.165-1-6, F.219-C7, F.251-1-6, F.nr 4115, F.nr 3922, F.nr 3987, F.nr 5049, Salmonella braenderup H9812.
Discussion
The present study shows that resistant F. psychrophilum isolates from rainbow trout have an amino acid substitution in the QRDR of gyrA at position 82 in which threonine is replaced by arginine. This is consistent with many previous studies associating QR in Gram -positive and -negative bacteria including F. psychrophilum.11,24,35 F. psychrophilum isolates from Denmark, Japan, and the United States have been reported to be resistant to oxolinic acid as a result of mutation in the A subunit of DNA gyrase.9,11 In the present study, no mutation was detected at amino acid position 87 in any of the isolates of F. psychrophilum, although a mutation at this position has been previously reported in this bacterium. 11
Quinolone antimicrobials act by inhibiting the activity of the enzymes DNA gyrase and topoisomerase IV. The main target for quinolones in Gram-negative bacteria is DNA gyrase, whereas in Gram-positive bacteria, it is the topoisomerase IV. Amino acid substitutions at positions 67, 81, 82, 83, 84, 87, and 106 of gyrA; 426 and 447 of gyrB; 78, 80, and 84 of parC; and 445 of parE have been reported to be responsible for resistance to quinolones in different bacteria. 24 The mutations causing amino acid replacement in either of the QRDRs in the two subunits of the target enzymes GyrA and ParC are widespread in many bacteria. While both gyrA and gyrB substitutions have been shown to lead to QR, 25 gyrA mutations appear to be more commonly involved in Enterobacteriaceae. 24 The mutation in the DNA sequence of the gyrA gene resulting in the threonine:arginine substitution at position 82 in the present case most probably, therefore, explains the reduced susceptibility towards quinolones in the Norwegian rainbow trout F. psychrophilum isolates. This finding is in accordance with the results of Izumi et al. 11 , who identified a threonine:alanine substitution at the same position in cold-water disease associated F. psychrophilum isolates from Japan. There are no specific guidelines available for evaluation of inhibition zone diameters and/or MIC values for antibiotic drugs and relation of these results to breakpoints for susceptible and resistant fish pathogenic bacteria. However, by using the recommendations by Kronvall 15 and Smith et al. 29 , and data of all strains in the present study, we calculated inhibition zone diameter breakpoints for oxolinic acid and flumequine to be 42 and 49 mm, respectively. The MICs of resistant and susceptible strains also correspond with previously studied isolates of F. psychrophilum. 22 We also identified numerous silent mutations (with the NCIMB 1947 sequence used as a basis for comparison) in the four genes encoding DNA gyrase and topoisomerase IV in Norwegian isolates of F. psychrophilum (Table 1). These synonymous point mutations may indicate a degree of specificity between strains of F. psychrophilum and host fish species in Norway, as all isolates from each of the different species of fish, gathered over a number of years, displayed identical point mutations in these four genes. Since a proportion of Norwegian rainbow trout eggs are imported, we suspect the pathogen may have been imported with infected eggs and then spread in the Norwegian population. The phylogenetic analysis also shows a close similarity between the Swedish strain and the Norwegian strains, which may indicate an association between the characterized QR and high virulence for rainbow trout.
Plasmid-borne qnr has mainly been associated with bacteria in Enterobacteriaceae but has also recently been identified with varying prevalence rates in a variety of plasmid-bearing bacterial genera.14,16,30 No isolates of F. psychrophilum have so far been identified with qnr genes including the results of the present study.
Quinolones are widely used in human medicine, aquaculture, and veterinary medicine. Monitoring for development of microbial resistance to both currently approved antimicrobial agents is important, in relation to both the preservation of the efficacy of these agents and minimization of transfer of resistance to human pathogens. 19
PFGE is considered a sensitive method for detailed molecular typing, and earlier reports of patterns of F. psychrophilum for the enzymes XhoI and KpnI have shown that patterns obtained using these enzymes provide a high degree of resolution.2,33 The consistency among the 2008 isolates from rainbow trout confirms the sequencing results showing evidence for a clonal relationship between these isolates and supports the hypothesis that a new virulent clone emerged in Norway that year. 19 Isolates from other rainbow trout strains tested show related but slightly different PFGE patterns, which may indicate that these represent different strains. These results are supported from their known history (isolated from outbreaks in rainbow trout either earlier or from other parts of the country).
Change in PFGE pattern between original strains and their progeny has been reported. 7 However this has not been shown for F. psychrophilum. 2 It cannot be excluded that such minor change has occured in F. psychrophilum of this study. However, sequencing results separating brown trout/salmon from rainbow trout strains correlated well with the PFGE results indicating no or minimal DNA sequence change in progeny isolates.
Altogether, the results obtained both from the sequencing (Table 1) and the PFGE patterns (Fig. 2 and 3) are consistent with earlier reports about species-specific infections of different genotypes within F. psychrophilum, exemplified by the QR in rainbow trout strains and the silent mutations in the salmon and trout strains.
In conclusion, the reduced susceptibility of F. psychrophilum isolated in Norway to quinolones appears to be related to a single point mutation in the QRDR of gyrA. All such strains were isolated from rainbow trout. This feature distinguishes these strains from other Norwegian isolates and supports other evidence indicating that these isolates are clonal. 21
The gyrB and parC phylogenetic analysis also provides further support for a clonal relationship between these strains.
In Norway, two quinolones; oxolinic acid and flumequine, are licensed for use in aquaculture. 5 Since only a limited number of antibiotics are licensed for use in Norwegian aquaculture, development of resistance against any of these antimicrobials is highly undesirable. There is a need, therefore, for a conservative approach in use of antibacterials and further development of effective vaccines against fish pathogenic bacteria, including F. psychrophilum.
Footnotes
Acknowledgment
We thank Nora Tandstad Martinussen, Norwegian Veterinary Institute, Oslo for assistance with the PFGE.
Disclosure Statement
Nothing to disclose.