
Abstract
Three hundred eighty-three isolates of Moraxella catarrhalis were collected from healthy children aged less than 2 years in China and assessed for antimicrobial resistance. We found that 92.2% (n=353) produced a β-lactamase. Nonsusceptibility rates to erythromycin and azithromycin, determined using Clinical Laboratory Standards Institute (CLSI) breakpoints, were 40.3% and 22.5%, respectively; nonsusceptibility rates determined using pharmacokinetics/pharmacodynamics breakpoints, however, were 59% and 60.1%. The minimal inhibitory concentration (MIC)90 values were >256 μg/ml. Nonsusceptibility rates varied by region from 9.7% in Dongguan to 75.9% in Jinan. Further, concomitant resistance to β-lactam antibiotics was also observed. Pulsed-field gel electrophoresis analysis of 27/37 high-level macrolide-resistant M. catarrhalis isolates showed that closely related pulsotypes dominated, with a total of 11 different pulsotypes being observed. The closely related pulsotypes were observed in isolates originating from all six Chinese cities investigated, possibly as a consequence of the mobility of the Chinese population. Sixteen patterns of 23S rRNA mutations were found among 97 selected isolates using polymerase chain reaction and sequencing, but no known ermA, ermB, mefA, or mefE genes could be detected. Mutations A2982T and A2796T in 23S rRNA were related to high-level macrolide resistance (MICs ranging from 24 to >256 μg/ml), while an A2983T mutation was associated with low-level macrolide resistance (MICs ranging from 0.19 to 16 μg/ml).
Introduction
All macrolides are defined as being composed of a derivatized macrolactone ring. This class of antibiotic targets the ribosome exit tunnel hampering nascent amino acid chain progression during polypeptide synthesis. Crystal structures of ribosome–macrolide interactions show that most ribosome exit tunnel interactions involve nucleotides A2058–A2059 (23S rRNA) as the main constituents of the macrolide-binding pocket, with dimethylation or mutations at these positions leading to macrolide resistance in bacteria. 38 In contrast, macrolide resistance in Streptococcus pneumoniae, also a bacterial pathogen of the upper respiratory tract, is not commonly associated with 23S rRNA mutations. Instead, the main macrolide resistance mechanisms in this organism are the carriage of target-site modification and drug efflux genes, with target-site modification being most frequently mediated by methylases that are encoded by the genes ermB or ermA, and drug efflux being mediated by the genes mefA and mefE.7,11,13,28 In this study, we investigated the presence of both 23S rRNA mutations and the presence of known macrolide resistance genes in macrolide-nonsusceptible M. catarrhalis.
Therefore, the goal of this study was to determine the prevalence of macrolide-nonsusceptible M. catarrhalis isolated from the nasopharynx of healthy children aged less than 2 years in China and to determine the resistance mechanisms of macrolide resistance.
Materials and Methods
Bacterial isolates
A total of 383 nonduplicate isolates of M. catarrhalis were collected from six cities (Beijing, n=65; Shanghai, n=66; Jinan, n=54; Nanjing, n=47; Wuhan, n=48; Dongguan, n=103) in China between May 2009 and May 2010. The isolates were taken from the nasopharynx of 3,622 healthy children less than 2 years of age. Isolates were initially identified using standard microbiological methods such as colony morphology, Gram-stain, oxidase and DNase production, and a nitrate reduction test at the sentinel laboratory in each city. All the culture medium (Thermo Fisher, Oxoid, Basingstoke, United Kingdom) and materials of biochemical reaction (Hangzhou Tianhe Microorganism Reagent Co., Ltd., Hangzhou, China) used for identification were purchased from the same manufacturers. Confirmatory analysis and antimicrobial susceptibility testing was performed at the Department of Clinical Laboratory, Peking Union Medical College Hospital (Beijing, China). Isolates were stored at −70°C until testing with a minimum number of passages.
Antimicrobial susceptibility testing
M. catarrhalis isolates were subcultured from storage onto horse blood agar and incubated overnight at 35°C in a reduced oxygen (5% CO2) environment. Freshly prepared Haemophilus test medium (HTM) was used for antimicrobial susceptibility testing, and growth of Haemophilus influenzae strains ATCC49247 and ATCC49766 were used for quality control. All isolates were tested for susceptibility to erythromycin, azithromycin, and amoxicillin using the E-test (BioMérieux, Marcy l'Etoile, France) method in accordance with the manufacturer's guidelines. To assess resistance to erythromycin and azithromycin, test plates were incubated at 35°C for 20–24 hr in ambient air, 35 while the test plates for amoxicillin were incubated in 5% CO2. Breakpoint interpretations were based on current Clinical Laboratory Standards Institute (CLSI) M45-A guidelines 9 or pharmacokinetics/pharmacodynamics (PK/PD) breakpoints.4,19 In addition, the broth microdilution method was used for comparison of antimicrobial susceptibility results of macrolide-nonsusceptible M. catarrhalis. Production of β-lactamase was performed by the nitrocefin test (Thermo Fisher, Oxoid).
copB polymerase chain reaction–restriction fragment length polymorphisms
Polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) patterns of the copB gene have been previously shown to be surrogate markers for the two major genotype lineages of M. catarrhalis, and this technique was used on macrolide-resistant isolates in this study, as described by Verhaegh et al. 34 A touchdown thermocycling program was used for all PCRs. The touchdown protocol used an initial annealing temperature of 70°C, which was reduced by 1°C per cycle over 15 cycles of PCR. The final 20 amplification cycles used an annealing temperature of 55°C. The copB PCR products were digested with RsaI (New England Biolabs, Ipswich, MA). Five units of RsaI were used per reaction mix, incubated at 37°C. The copB PCR-RFLP typing method can distinguish four CopB types: type 0, consisting of 374 and 157 bp; CopB types I/III, consisting of 342 and 157 bp; CopB type II, with bands of 332 and 187 bp; and CopB type IV, consisting of a single band of 519 bp.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) was performed as described by Yano et al. 42 Briefly, overnight cultures were suspended in buffer and mixed with 2.0% low-melting point agarose to form an agar insert. After removing plugs from the suspension buffer they were treated with 1–5 mg of lysozyme/ml (Seikagaku, Tokyo, Japan) of lysis solution (1 M NaCl, 0.1 M EDTA [pH 8.0], 10 mM Tris-HCl, 0.5% Brij 58, 0.2% sodium deoxycholate, and 0.5% Sarkosyl) and incubated at 37°C for 12 hr. Lysis solution was then decanted and replaced with 0.1–1.0 mg of proteinase K/ml (Merck, Sharp & Dohme [China] Ltd., Shanghai, China) of EDTA and Sarkosyl (ES) solution (0.25 M EDTA [pH 8.0] and 1% Sarkosyl) at 56°C for 24 hr. The ES solution was decanted, and the plugs were placed in TE buffer (10 mM Tris-HCl [pH 8.0] and 1 mM EDTA [pH 8.0]) containing 1 mM phenylmethylsulfonyl fluoride at ambient temperature for 4 hr. Plugs were washed in TE buffer for 40 min at ambient temperature. For restriction endonuclease digestion, plugs were incubated in enzyme restriction buffer for 30 min at ambient temperature to remove the EDTA. The plugs were incubated at 37°C in restriction enzyme buffer with 20 U of SpeI (TaKaRa, Tokyo, Japan) for 4 hr.
Electrophoresis was performed using a CHEF Mapper (Bio-Rad, Hercules, CA). Agarose gels were prepared at a 1% concentration in 0.5×TBE buffer (45 mM Tris base, 45 mM boric acid, and 1 mM EDTA [pH 8.0]). DNA fragments were separated using an electric current of 6 V/cm at 14°C for 18 hr. The pulse time, which changed linearly, was 5–8 s. Lambda Ladder (NEB) was used as a molecular size standard. Gels were stained for 30 min in ethidium bromide (1 μg/ml) and decolorized in distilled water for 15 min. Fluorescent bands were visualized using UV transillumination and gel images were captured using Bio-rad GelDocEQ (Bio-rad).
Pulsotypes were analyzed using BioNumerics software (Applied Maths, Sint-Martens-Latem, Belgium). Isolates with a similarity score of<85% were considered to represent different lineages, while a similarity of >85% were considered to represent closely related isolates; only isolates that had a similarity score of 100% (indistinguishable) were considered to be the same clone.
PCR detection of macrolide resistance determinants
PCR was used to detect the presence of genes known to cause macrolide resistance (ermA, ermB, mefA, and mefE) among 171 strains of M. catarrhalis (all of the 153 macrolide-nonsusceptible M. catarrhalis strains, and 18 macrolide-susceptible M. catarrhalis isolates randomly selected from 230 macrolide-susceptible isolates), as described by Mazzariol et al. 25 DNA from M. catarrhalis isolates was extracted using the QIAamp DNA Mini Kit (Qiagen, Dusseldorf, Germany), with DNA purity and concentration being determined using a NanoDrop ND-1000 (NanoDrop, Wilmington, DE). A 260 nm/280 nm absorbance ratio was used to assess the purity of DNA. A ratio of ∼1.8 was generally accepted as pure for DNA. Isolates of S. pneumoniae known to harbor ermA, ermB, mefA, or mefE genes were used as positive controls for PCR, and repeat PCRs were performed using a 2°C–4°C reduction in Tm to try to amplify any erm- and mef-resistance determinants that were originally PCR negative for erm- and mef-related genes.
PCR and sequencing analysis of the 23S rRNA
PCR and sequencing was used to detect the presence of mutations in a 934-bp region of the 23S rRNA gene in 171 isolates of M. catarrhalis (i.e., all 153 macrolide-nonsusceptible M. catarrhalis isolates, and 18 macrolide-susceptible M. catarrhalis isolates randomly selected from the 230 macrolide-susceptible isolates cultured in this study). Our laboratory developed oligonucleotide primer pairs MUT-F, 5′-2685CAGGCTGCTGCAACTGTTTA2704-3′, and MUT-R, 5′-3618CAACCGAAACA CCAGAGGTT3599-3′ (based on the GenBank sequence with accession number FJ410380, from M. catarrhalis KCCM:40056; www.ncbi.nlm.nih.gov/nuccore/FJ410380), for amplification using Primer 3 software (http://frodo.wi.mit.edu/primer3/). PCR was performed in volumes of 50 μl, and 2×TransTaq High Fidelity (HiFi) PCR SuperMix I (TransGen Biotech, Beijing, China) was used for amplification, with an initial denaturation temperature of 95°C for 10 min, followed by 35 cycles of 95°C for 15 sec, 61°C for 15 sec, 72°C for 30 sec, and a final extension of 72°C for 7 min. DNA sequencing was performed using the same primers used for PCR amplification, providing bidirectional coverage. Results were aligned with sequences directly obtained from GenBank (accession number FJ410380). Genomic DNA extracted from each of the 171 isolates was amplified and sequenced three times each time using freshly synthesized primers. The frequency of mutations in the two MIC groups was statistically compared using Pearson's χ2-test (or the Fisher exact test, when appropriate) for categorical data.
Results
Antimicrobial susceptibility testing
The activities of erythromycin, azithromycin, and amoxicillin against M. catarrhalis according to current CLSI M45-A guidelines and PK/PD breakpoints are shown in Table 1. Erythromycin and azithromycin were less active against M. catarrhalis than amoxicillin; MICs against M. catarrhalis ranged from 0.047 to >256 μg/ml for erythromycin and from 0.023 to >256 μg/ml for azithromycin, with an MIC90 value of >256 μg/ml for both antibiotics. Based on the CLSI breakpoints for M. catarrhalis, the nonsusceptibility rates of erythromycin and azithromycin were 40.3% and 22.5%, respectively. By PK/PD breakpoints, the nonsusceptibility rates of erythromycin and azithromycin were 59% and 60.1%, respectively.
No breakpoints were available from the current CLSI M45-A guideline, and the PK/PD breakpoint was used.
By PK/PD breakpoints: S≤0.25 μg/ml, erythromycin; S≤0.12 μg/ml, azithromycin.
CLSI, Clinical Laboratory Standards Institute; PK/PD, pharmacokinetics/pharmacodynamics; MIC, minimal inhibitory concentration; R, resistant; I, intermediate; S, susceptible.
Of 383 isolates of M. catarrhalis, ∼92.2% (353/383) isolates produced a β-lactamase. Among these β-lactamase-producing M. catarrhalis, the nonsusceptibility rates of erythromycin and azithromycin were 43.1% and 24.3%, respectively (CLSI breakpoints). However, based on the PK/PD breakpoints, the nonsusceptibility rates were 63.2% for erythromycin and 60.6% for azithromycin. Interestingly, among the 30 remaining isolates of M. catarrhalis with no detectable β-lactamase, erythromycin and azithromycin displayed potent activities. Based on CLSI breakpoints, the nonsusceptibility rates were 6.7% for erythromycin and 0% for azithromycin, with an MIC90 value of 0.25 μg/ml. Interpreting the antimicrobial resistance using PK/PD breakpoints suggested that the nonsusceptibility rates of erythromycin and azithromycin were 10% and 53.3%, respectively. Thus, our data indicate that MIC distributions and nonsusceptibility rates of erythromycin and azithromycin were higher in β-lactamase-producing versus β-lactamase-nonproducing M. catarrhalis using both CLSI and PK/PD breakpoints. Further, for isolates with increasing MIC values of amoxicillin, we observed an increase in erythromycin nonsusceptibility rates from 20.9%/28.4% to 66.7%/76.2% using either the CLSI or the PK/PD breakpoints (Fig. 1). Significant differences between erythromycin nonsusceptibility rates were observed when comparing the amoxicillin-susceptible and amoxicillin-nonsusceptible groups (erythromycin, CLSI: χ2=12.915, p<0.001; erythromycin, PK/PD: χ2=33.373, p<0.001).
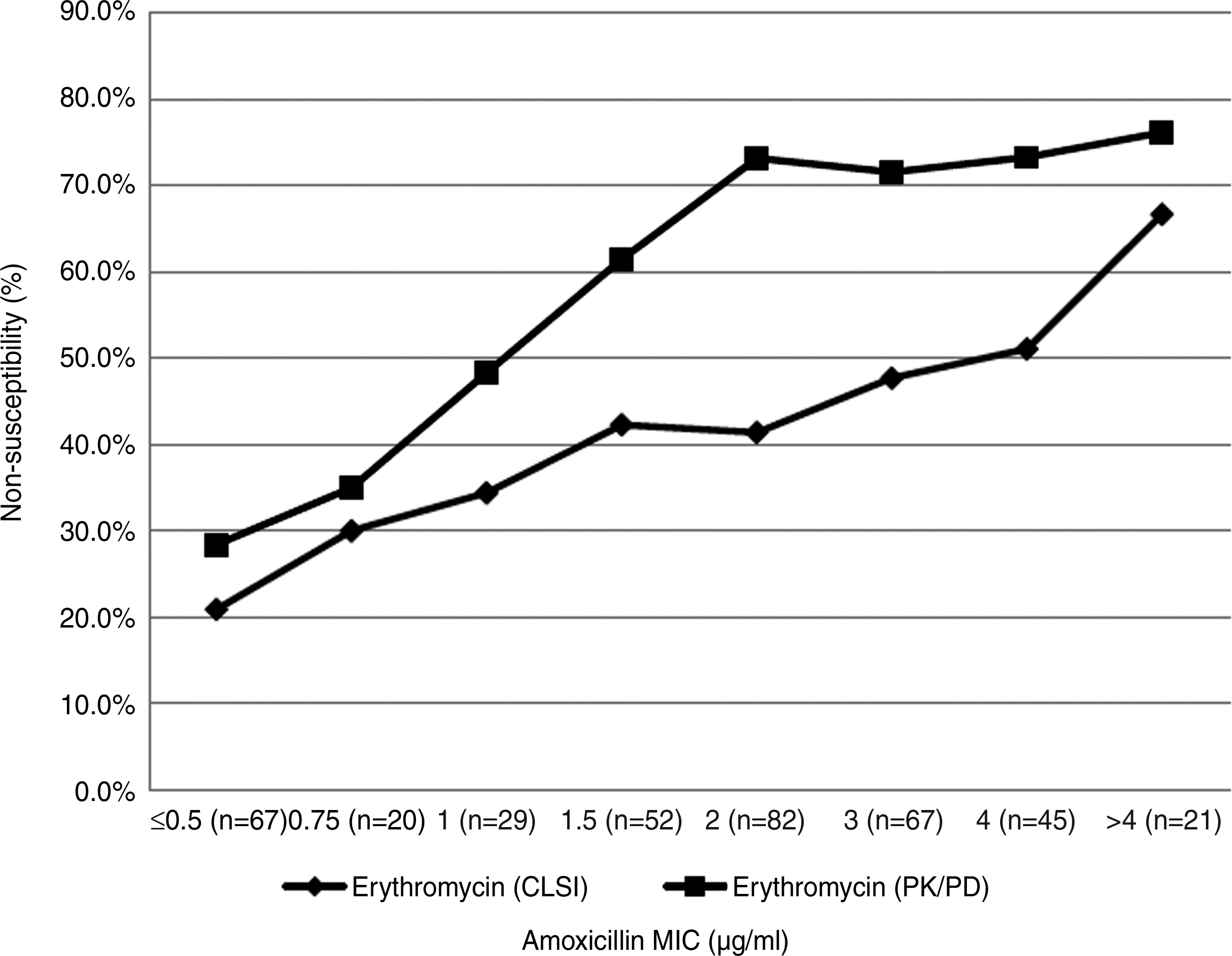
The relationship between MIC values of amoxicillin and the erythromycin nonsusceptibility rates analyzed using CLSI breakpoints and PK/PD breakpoints. CLSI, Clinical and Laboratory Standards Institute; PK/PD, pharmacokinetics/pharmacodynamics; MIC, minimal inhibitory concentration.
We observed that M. catarrhalis isolates from six cities showed differences in nonsusceptibility rates to macrolide (Fig. 2). Significant differences were observed in different geographical regions (erythromycin: χ2=45.483, p<0.001; azithromycin: χ2=35.295, p<0.001). The highest nonsusceptibility rates of erythromycin (75.9%) and azithromycin (48.1%) were found in Jinan, followed by Beijing, with nonsusceptibility rates of 50.8% for erythromycin and 29.2% for azithromycin. Lower nonsusceptibility rates were observed in relatively southern cities, such as Nanjing, Shanghai, Wuhan, and Dongguan.
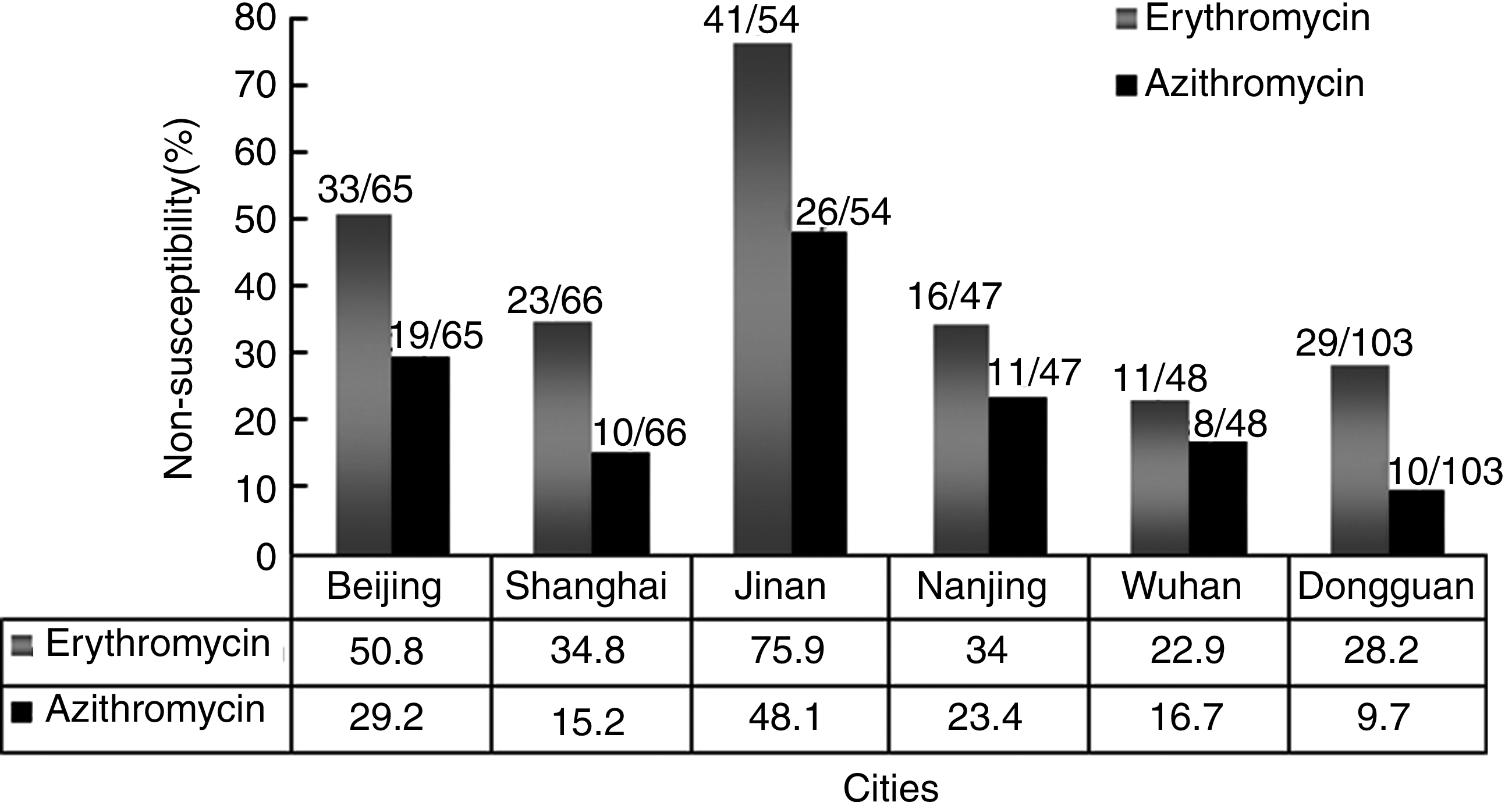
The macrolide-nonsusceptibility rates in Moraxella catarrhalis isolated from six cities.
copB PCR-RFLP
All high-level (MICs >256 μg/ml) macrolide-resistant isolates obtained from Jinan (n=22) and 15 high-level (MICs >256 μg/ml) macrolide-resistant isolates randomly selected from the five other cities (Beijing, n=3; Shanghai, n=3; Nanjing, n=3; Wuhan, n=3; Dongguan n=3) were examined using the copB PCR-RFLP method. The results showed that the majority of isolates, 62.6% (23/37), belonged to copB PCR-RFLP type II, followed by type I/III (27.0%, 10/37) and type IV (10.8%, 4/37); type 0 was not found in our study. These results indicate that the majority of macrolide-resistant isolates belonged to the M. catarrhalis Lineage 1 genotype, a lineage previously shown to be associated with increased virulence potential and serum resistance. 14
Pulsed-field gel electrophoresis
We utilized PFGE analysis to determine the clonal relationship of 27 out of the 37 high-level macrolide-resistant isolates (Fig. 3). Ten of the 37 isolates were found to be untypable using this method. We observed 11 pulsotypes among the 27 isolates, with a single pulsotype appearing to predominate (isolates M1-80, M1-82, M1-69, M1-74, M1-75, M2-128, M1-81, M2-65, M2-77, M2-78, M2-203, M2-229, M2-184, and M1-15). Isolates obtained from the same city appeared clonal, such as M2-65, M2-77, and M2-78 (Shanghai); M2-203 and M2-229 (Dongguan); and M1-80 and M1-82 (Jinan). There was some evidence of clonal dissemination of isolates across the cities; isolates M1-69, M1-74, and M1-75 from Jinan were indistinguishable from M2-128 isolated from Nanjing, and M1-25 isolated from Beijing was indistinguishable from M1-68 isolated from Jinan. However, pulsotypes were not significantly associated with CopB RFLP and 23S rRNA sequence types. For example, M1-81 (pulsotype 3), M1-71 (pulsotype 5), M2-231 (pulsotype 7), M1-86 (pulsotype 9), and M2-136 (pulsotype 10) isolates had the same CopB RFLP and 23S rRNA sequence but were all unique pulsotypes.
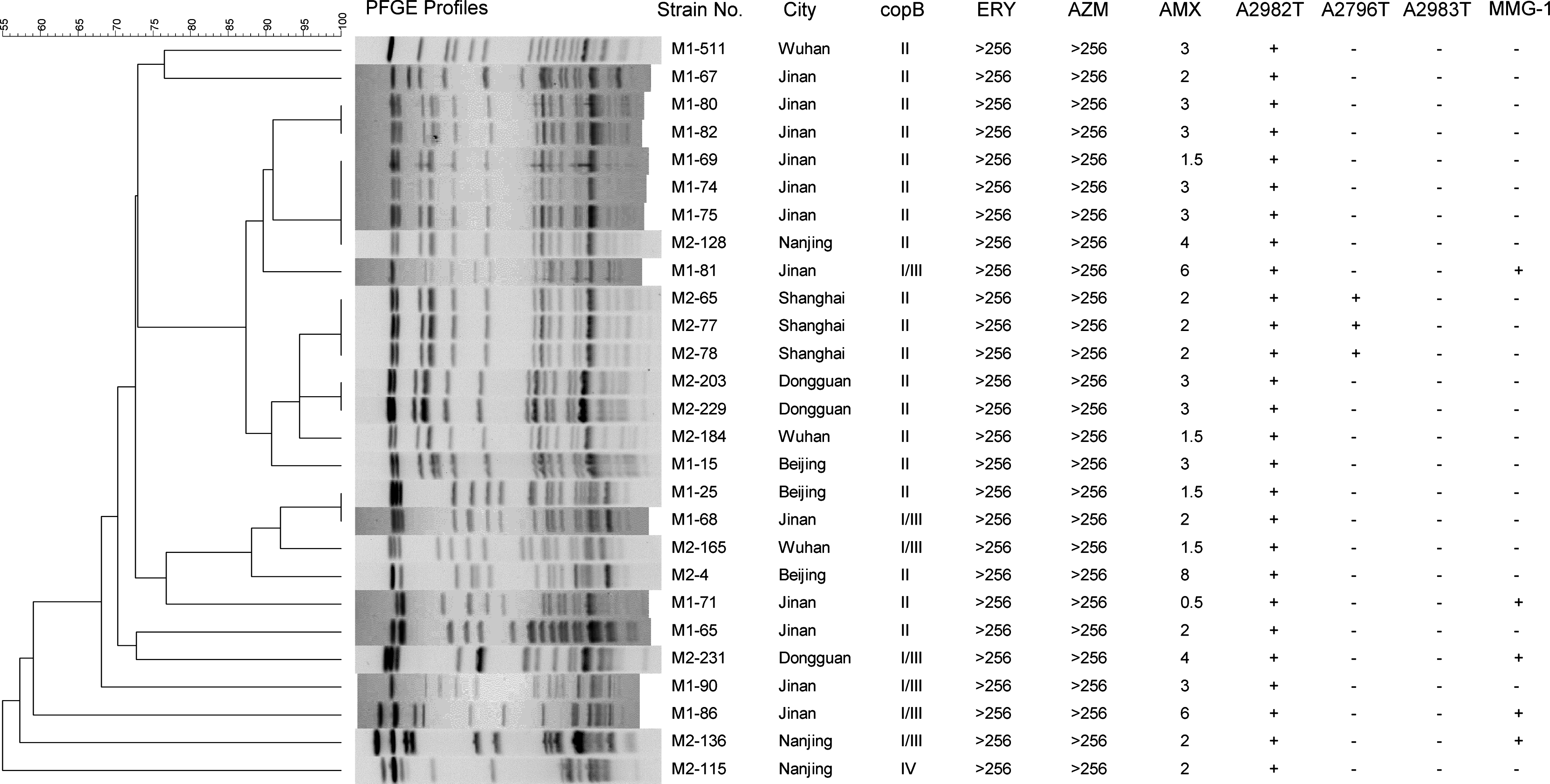
Dendrogram of 27 PFGE profiles of M. catarrhalis isolated from six cities. M2-65 was used as the reference strains for different PFGE types. MMG-1: mutations located at G3129A, A3130T, T3131A, A3137G, A3138T, T3139A, and C3140T coexist in the same strain of M. catarrhalis. A similarity of<85% at dendrogram analysis was considered to represent different PFGE types, while a similarity of >85% was considered to represent PFGE pattern subtypes. Only a similarity of 100% was considered to be the same clone. ERY, erythromycin; AZM, azithromycin; AMX, amoxicillin; PFGE, pulsed-field gel electrophoresis; MMG-1, multiple mutations group-1.
PCR detection of macrolide resistance determinants
Based on the mechanisms of macrolide resistance that exist in other species,25,28,39 we screened our isolates for known ermA, ermB, mefA, and mefE genes. We were unable to find resistance genes (ermA, ermB, mefA, and mefE) in the 171 isolates examined (data not shown), although our positive control was detected and confirmed by sequencing. We altered annealing temperatures from 52°C (mef), 48°C (ermA), and 56°C (ermB) to 50°C (mef), 45°C (ermA), and 52°C (ermB), and while nonspecific bands were detected, none were confirmed by sequencing. These results suggest that other determinants or mechanisms are likely responsible for the macrolide-nonsusceptible phenotype observed in M. catarrhalis, such as mutations in the 23S rRNA.
PCR and sequence analysis of the 23S rRNA gene
We observed 23S rRNA sequence changes in 97 of the 171 isolates of M. catarrhalis as detected by PCR and sequencing. Relative to the reference isolate (M. catarrhalis KCCM:40056), sixteen 23S rRNA sequence patterns were found in the 97 isolates of M. catarrhalis. The accession numbers for the representative 23S rRNA sequences as characterized by isolates M2-25, M2-61, M2-91, M1-5, M1-106, M2-107, M2-77, M2-82, M2-126, M2-231, M2-4, M2-101, M2-103, M2-147, M1-115, and M2-178 were HQ420307, HQ420306, HQ420308, HQ420305, JN164687, JN164688, JN164689, JN164690, JN164691, JN164692, JN164693, JN164694, JN164695, JN164696, JN164697, and JN164698, respectively. Among the 84 isolates of erythromycin and azithromycin-nonsusceptible M. catarrhalis sequenced, 83 (98.8%) were found to have an altered 23S rRNA sequence compared with the reference isolate. Fifteen patterns of 23S rRNA sequence were identified with nucleotide position alterations at T2779A, A2796T, C2811T, A2910C, G2911A, A2982T, A2983T, A3029G, T3037C, T3055A, C3084T, T3108C, A3127G, G3129A, A3130T, T3131A, C3132T, A3133C, A3137G, A3138T, T3139A, C3140T, A3142G, and a T insertion at position 3130 [FJ410380.1(23S rRNA):g.3130insT]. The A2796T mutation always colocated with the A2982T mutation. Among the 69 isolates of erythromycin-nonsusceptible and azithromycin-susceptible M. catarrhalis, 9 isolates (13.4%) deviated from the reference isolate. Within these isolates, four patterns of 23S rRNA sequence were found containing A2983T, A3029G, T3037C, T3055A, C3061T, A3064G, T3065G, A3074T, T3075C, T3076C, G3078A, C3084T, T3108C, G3129A, A3130T, T3131A, C3132T, A3133C, A3137G, A3138T, T3139A, C3140T, A3142G, and a T insertion at position 3130 [FJ410380.1(23S rRNA):g.3130insT]. However, among the 18 isolates of erythromycin- and azithromycin-susceptible M. catarrhalis sequenced, only one change, C3132T, was found in 5 (27.8%) isolates; the MIC values of these isolates ranged from 0.125 to 0.19 μg/ml, which was very close to the PK/PD breakpoints of 0.25 μg/ml for erythromycin and 0.125 μg/ml for azithromycin.
All of the results mentioned above indicated that mutations in the 23S rRNA gene are implicated in the development of nonsusceptibility to macrolide in M. catarrhalis. To help standardize results, we have also generated an alignment table (Table 2) that documents the alignment of the 23S rRNA changes outlined in the current publication with the recently sequenced M. catarrhalis isolate RH4.10,15
The relationship of nucleotide changes in M. catarrhalis to erythromycin and azithromycin MIC values is shown in Fig. 4. Using either the CLSI or the PK/PD breakpoints, the A2982T and A2796T changes conferred high-level erythromycin and azithromycin resistance to all isolates, with MIC values ranging from 24 to >256 μg/ml. As previously mentioned, the A2796T alteration was always combined with A2982T. We observed that isolates possessing either the substitution A2983T, or the multiple mutations group 1 (MMG-1 mutations located at G3129A, A3130T, T3131A, A3137G, A3138T, T3139A, and C3140T in the same isolate of M. catarrhalis) exhibited different MIC values depending on the breakpoint system (CLSI or PK/PD) used (Fig. 4); divergence was notable for azithromycin, but there was less difference for erythromycin. Based on PK/PD breakpoints, we observed that isolates possessing the A2983T mutation conferred low-level erythromycin and azithromycin resistance, with MIC values ranging from 0.38 to 16 μg/ml and from 0.19 to 16 μg/ml, respectively. Isolates possessing MMG-1 also had comparable erythromycin and azithromycin resistance, with MIC values ranging from 0.38 to >256 μg/ml and from 0.19 to >256 μg/ml, respectively. However, differences in the significance of detection of the A2983T and MMG-1 mutations and erythromycin and azithromycin nonsusceptibility were observed when applying CLSI breakpoints, as both A2983T and MMG-1 mutations could be detected in azithromycin-susceptible M. catarrhalis isolates (Fig. 4). The frequency of mutations in the two MIC groups was compared using a χ2-test, and the data indicated that the presence of the A2983T mutation between the two MIC groups (susceptible group,≤2 μg/ml; low-level-resistant group, 3–16 μg/ml based on CLSI breakpoints) was statistically different (χ2=30.231, p<0.001); the presence of the MMG-1 set of changes between the two MIC groups (susceptible,≤2 μg/ml; nonsusceptible, 3 to >256 μg/ml based on CLSI breakpoints) was also statistically different (χ2=8.971, p<0.001). This analysis indicated that an increased number of isolates carrying the A2983T mutation or the MMG-1 mutation are found in the low-level-resistant group or the nonsusceptible group as defined by CLSI breakpoints.
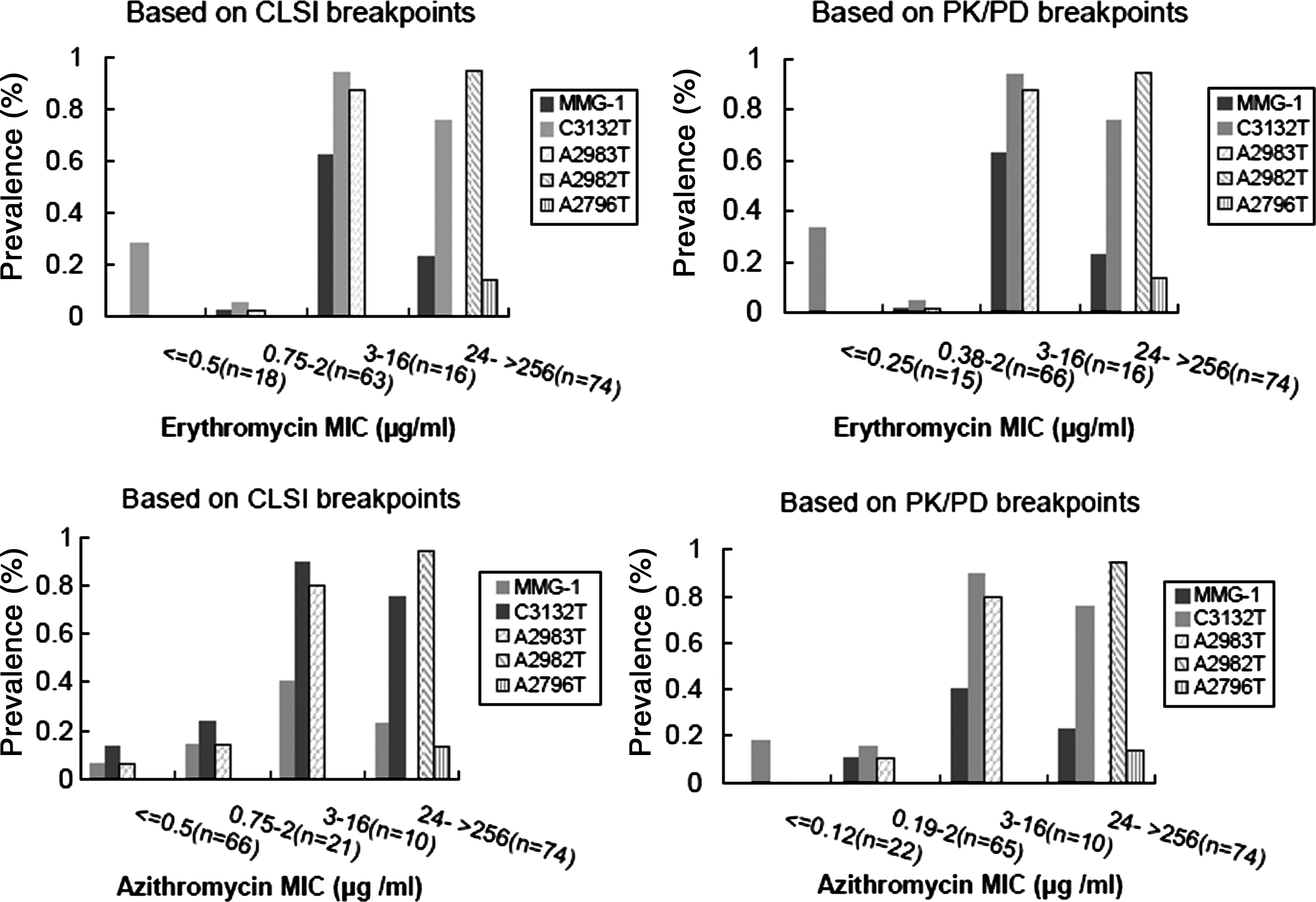
The relationship of nucleotide changes in M. catarrhalis (n=171) to erythromycin and azithromycin MIC values, based on CLSI and PK/PD breakpoints. MMG-1: mutations located at G3129A, A3130T, T3131A, A3137G, A3138T, T3139A, and C3140T of FJ410380 coexist in the same isolate of M. catarrhalis. The CLSI breakpoints: S≤0.5 μg/ml, erythromycin; S≤2 μg/ml, azithromycin. The PK/PD breakpoints: S≤0.25 μg/ml, erythromycin; S≤0.12 μg/ml, azithromycin.
Discussion
Our findings, based on isolation and antimicrobial susceptibility testing, confirm that there is a high prevalence of macrolide-nonsusceptible M. catarrhalis carried in the nasopharynx of healthy children in some regions of China, especially in Jinan and Beijing. Similarly, cross-resistance can be detected between β-lactam and macrolide antibiotic classes among these isolates, with isolates having high MIC values of amoxicillin and being significantly more likely to be erythromycin nonsusceptible. Closely related pulsotypes, being observed in isolates originating from all six Chinese cities investigated, dominated among the macrolide-nonsusceptible M. catarrhalis isolates tested. In this respect, further research is necessary to determine whether the mobility of the Chinese population has contributed to the spread of macrolide-nonsusceptible M. catarrhalis between Chinese cities.
In this study, we also found that many changes in the 23S rRNA relative to a reference isolate (FJ410380) of M. catarrhalis found in GenBank may be associated with macrolide resistance. We selected FJ410380 as a reference sequence based on the fact that M. catarrhalis KCCM: 40056 is often used as a reference strain, 43 and positions at the sequence of FJ410380.1 were easily defined for the small size of 3,681 bp. A recently complete genome, Cp002005.1 (M. catarrhalis RH4, complete genome), 15 contained four operons that we also used to define the positions of the 23S rRNA. We created an alignment of all of our 23S rRNA mutations (FJ410380.1) against the four different operons of this complete genome (Table 2). RH4 is currently the gold standard for M. catarrhalis sequence comparisons. Macrolide resistance in M. catarrhalis was associated with the detection of A2982T, A2796T, A2983T, and MMG-1 mutations in 23S rRNA, but the number of changes did not correlate with increased MIC values; this result suggests that stepwise alterations in the 23S rRNA may not be responsible for M. catarrhalis resistance. In addition, the C3132T mutation may not be as important for resistance as other mutations because it was found in some susceptible isolates; this situation is similar to C2611T in S. pneumoniae. 28 Finally, our findings have also suggested that the conservative PK/PD breakpoints may be much better than CLSI breakpoints when determining the relationship between macrolide resistance and mutations in 23S rRNA. Therefore, we predict that some clinical isolates with mutations in 23S rRNA might be considered as susceptible to macrolide based on CLSI breakpoints, which may lead to treatment failure due to the reduced antimicrobial activity of macrolide against M. catarrhalis. However, using the PK/PD breakpoints could potentially avoid this problem. Additional experimentation and observation are necessary to confirm this hypothesis.
Compared globally, China appears to have low macrolide-susceptibility rates. Recently, a study from the Community-Acquired Respiratory Tract Infection Pathogen Surveillance (CARTIPS) from 2009 to 2010 reported that 5.8% of M. catarrhalis isolates were resistant to macrolides and that these isolates were only found in mainland China. 36 Further, the exact mechanisms facilitating this resistance is still unknown. In our study, we confirmed the MIC values obtained by E-test using the broth microdilution method. Although Hoogkamp-Korstanje et al. 20 reported that significant disagreements in the interpretation (fourfold or greater MIC differences) in M. catarrhalis resistance to clarithromycin depended on whether E-test or broth microdilution was used, these disagreements were not observed for the MIC values obtained in this study using related macrolide antibiotics. In addition, all of the HTM plates containing erythromycin or azithromycin were incubated in ambient air because CO2 can elevate macrolide MICs. 12
Our results confirm that the frequency of nonsusceptibility to macrolide by M. catarrhalis is greater than previously reported in China, though the reason for this is not exactly clear. In fact, many factors may contribute to the increased frequency of nonsusceptibility to macrolide observed in this publication.
The problem of frequent and inappropriate use of antibiotics is very serious in China, with broad-spectrum antibiotics, new antibiotics, and even combinations of two or three kinds of antibiotics often being prescribed for RTI. This prescribing policy compares unfavorably with the narrow-spectrum, older, and single antibiotic prescriptions that tend to be prescribed in northern European countries.3,24 Further, the empirical use of antibiotics for RTI is common in China, even though the majority of infections are caused by viruses. 23 Also, different doses, routes, duration, and combinations of different types of antibiotics are used in clinical practice, which all favor the selection of antimicrobial-resistant organisms. Finally, antibiotics can be purchased from private retail pharmacies, private clinics, and even the public sector, either with or without a prescription, meaning that antibiotics are frequently taken by patients who are unaware or unconcerned of the growing problems associated with antimicrobial resistance. 5 For example, erythromycin can be purchased from nearly all private clinics or retail pharmacies, and azithromycin has wide community use. Unfortunately, some of these prescribing practices are also shared by other countries, with ∼80% of antibiotics being used in the community, and the rest in more restricted hospital environments.8,40
The migration of people and increasing population density can lead to increased person to person contact, facilitating the spread of resistant bacteria and their antimicrobial resistance genes. 31 At least one study has shown that there is a strong correlation between the defined daily dose/1,000 inhabitants/day of antimicrobials and the prevalence of resistance when population density is taken into consideration. 6 Further, socioeconomic status is also an important factor with regard to antibiotic resistance. For example, a study from India 22 demonstrated that a higher socioeconomic status (educated, professional, white-collar, and skilled individuals) is associated with higher antibiotic use/abuse.
Another important factor facilitating the possible spread of antimicrobial resistance in China is the lack of an antimicrobial surveillance system for the use/abuse of antimicrobials in the community. Accurate statistical data are urgently required to fully assess any current problems and future trends with regard to antimicrobial resistance.
Finally, the authors speculate that all of the above phenomena are operating in China at the present moment in time, all possibly contributing to an increased incidence of antimicrobial resistance among its >1 billion inhabitants.
A second question from our work is how the 23S rRNA changes observed in this publication confer macrolide resistance to M. catarrhalis. From previous research, it is known that nascent peptides emerge out of the ribosome through an exit tunnel, a universal feature of the large ribosomal subunit, which is the target of macrolide antibiotics. Further, the entrance to this tunnel is adjacent to a peptidyl transferase center (PTC). 1 Macrolide have been shown to block the tunnel that channels the nascent peptides away from the PTC, hampering the progression of nascent amino acid chains. In the process, the macrolide can form hydrogen bonds with the peptidyl transferase cavity at three positions: N6 and N1 of A2058Ec (corresponding to A2982 in FJ410380) and N6 of A2059Ec (corresponding to A2983 in FJ410380). This explains why these nucleotides are essential for macrolide binding, thus shedding light on the importance of these positions in the development of macrolide nonsusceptibility. Changes at the equivalent of A2058Ec and A2059Ec in M. catarrhalis have the potential to disrupt hydrogen bonding patterns at the PTC, thereby impairing the ability of macrolides to interfere with the ribosomal exit tunnel, and rendering bacterium nonsusceptible to macrolides. 32 In fact, the two mutations A2982 and A2983 in the 23S rRNA of our M. catarrhalis isolates are equivalent to positions located in Escherichia coli (A2058 and A2059), Helicobacter pylori (A2142 and A2143), S. pneumoniae (A2058 and A2059), Mycoplasma pneumoniae (A2063 and A2064), and others.28,30,41 Although we are not able to provide a definitive explanation for the exact mechanism facilitating these 23S rRNA changes and macrolide nonsusceptibility, it is most likely that mutation alters the conformation or shape of the 23S rRNA, thereby affecting the ability of macrolide to block amino acid synthesis.
In conclusion, we have shown a high prevalence of macrolide nonsusceptibility in M. catarrhalis isolated from the nasopharynx of healthy children aged less than 2 years, which underscores the need for surveillance of macrolide-nonsusceptible M. catarrhalis, especially in China. Although PCR screening was performed to detect known genes that encode enzymes associated with modifying macrolide antibiotics, none of these genes were actually detected. However, several changes in the 23S rRNA sequence such as A2982T, A2983T, A2796T, C3132T, and MMG-1 mutations were detected in macrolide-nonsusceptible M. catarrhalis, and these changes suggest that ribosomal mutations are critical for creating macrolide-nonsusceptible M. catarrhalis.
Footnotes
Acknowledgments
We thank John Klena (International Emerging Infections Program, U.S. CDC, China office) and Evan Sander (University of Tennessee Health Science Center College of Medicine) for their careful revision of our article. We also thank Zhigang Cui, Jianzhong Zhang, and Yuan Hu (National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention, China) for their analysis on PFGE profiles using BioNumerics software.
Disclosure Statement
No competing financial interests exist.