
Abstract
Introduction
Clinical isolates of P. aeruginosa producing acquired β-lactamases such as GES-5 have caused outbreaks in several hospitals in different regions of the world, such as China, 33 South Africa, 15 Brazil, 20 and Spain. 32 VIM-family MBLs were first described in Europe in 1999 and since that time, specific reports on VIM-11 19 and VIM-13, 14 as well as on other VIM variants, have been made worldwide, at present reaching a family with 32 subtypes 18 (nomenclature is in accordance with that provided by G. Jacoby on the Lahey website available at www.lahey.org/studies/other.asp#table1]). The simultaneous presence of genes encoding MBLs and/or ESBLs has been described in P. aeruginosa. 32 In Mexico, blaVIM-2 was found in Escherichia coli isolates in 2009 17 and simultaneously with blaIMP-15 in P. aeruginosa clinical isolates in 2010. 24 In addition, a blaVIM-23 variant was identified in an Enterobacter cloacae isolate (GenBank accession number GQ242167).
The aim of this study was to determine the diversity of β-lactamases in a group of multidrug-resistant (MDR) isolates of P. aeruginosa involved in nosocomial infections in Mexico in 2004, demonstrating the novel combination blaGES-5 and blaVIM-11, as well as the previously reported blaVIM-2 and blaOXA-2.
Materials and Methods
Biological material
Fourteen imipenem- and ceftazidime-resistant P. aeruginosa isolates were studied. 5 These isolates were obtained from cases of nosocomial infections at the Intensive Care Service, Hospital of Infectious Diseases, La Raza National Medical Center, Mexican Institute of Social Security (IMSS) in Mexico City, from August to November 2004. Bacteriological identification was performed with the BBL Crystal® system (Becton, Dickinson and Company), and three isolates were selected at random to confirm identification at the molecular level, analyzing their 16S rDNA gene partial sequence (results not shown).
Molecular typing and susceptibility to antibiotics
The clonal relationship of isolates was studied by pulsed-field gel electrophoresis (PFGE), as described by Spencker et al. 29 Genomic DNA was subjected to digestion with SpeI. Macrorestriction patterns of DNA were interpreted under the criteria established by Tenover et al. 30 and the dendrogram-derived degree of genetic relatedness. The dendrogram was derived from the PFGE results from the matrix of correlation distances by using the Jaccard similarity coefficient with the Dendro-UPGMA program (http://genomes.urv.cat/UPGMA). 10
Susceptibility to 21 antibiotics (amikacin, azlocillin, aztreonam, carbenicillin, cefepime, cefoperazone, ceftazidime, ceftizoxime, ceftriaxone, ciprofloxacin, imipenem, levofloxacin, meropenem, mezlocillin, netilmicin, norfloxacin, ofloxacin, piperacillin, ticarcillin, ticarcillin/clavulanic acid, and tobramycin) was assessed by the disk diffusion method utilizing filter paper discs and following the recommendations of the Clinical and Laboratory Standard Institute (CLSI). 6
The MBL phenotype was detected with E-test metallo-β-lactamase strips (E-test MBLs; AB Biodisk) employed according to the manufacturer's instructions. ESBL detection was performed with the double-disk synergy test (DDST) with ceftazidime and amoxicillin/clavulanate with a 10–20 mm separation. 13
Detection, characterization, and cloning of acquired β-lactamases
To have a biochemical approximation to the potentially acquired β-lactamases, crude protein extracts were prepared by sonication for determination of isoelectric focusing (IEF) using Phast gels (pH gradient, 3–9) in a Phast System apparatus (Pharmacia AB). The extracts from four control strains of the collection of the Son Dureta Hospital were subjected in parallel to electrophoresis CTX-M-15 producing Klebsiella pneumoniae (isoelectric point [pI], 8.6), 9 GES-1, GES-5, and OXA-2 producing P. aeruginosa (pI, 5.9, 5.8, and 7.7, respectively), 32 TEM-1 and CTX-M-14 producing E. coli (pI, 5.4 and 8.0, respectively), 9 and SHV-12 producing E. coli (pI, 8.2). 32 For genetic characterization, polymerase chain reactions (PCRs) were performed using as a template the genomic DNA obtained with the DNeasy Tissue kit (Qiagen), as well as specific primer sequences previously reported to identify β-lactamase genes, including GIM, IMP, SIM, SPM, and VIM,12,23 GES, 32 and OXA 27 (Table 1). Amplicons obtained by PCR and cloned DNA fragments were sequenced in ABI PRISM 3100 automated equipment® (PE Applied Biosystems). The resulting sequences were then compared with those available at GenBank (www.ncbi.nih.gov/BLAST).
Used only for sequencing.
When the IEF assay showed bands with a pI corresponding to β-lactamases reported in the literature but whose presence was not detected by PCR, cloning of the involved gene was attempted. Cloning was performed in E. coli XL1-Blue or E. coli DH5-α utilizing the CaCl2 method with ligation reactions of pUCP24 and total DNA digested with EcoRI and BamHI. 26 The transformants were selected on Luria Bertani (LB) agar plates containing 5 mg/L gentamicin and 30 mg/L ampicillin. Since the potential presence of multiple integrons simultaneously in the same isolate may interfere with PCR and sequencing experiments, cloning was carried out to detect integrons and/or genetic regions containing resistance genes. ESBL and MBL activity was analyzed in the obtained transformants as previously described (DDST and E-test MBLs). Minimal inhibitory concentration (MIC) to the antibiotics piperacillin, piperacillin/tazobactam, ceftazidime, cefepime, imipenem, meropenem, gentamicin, tobramycin, amikacin, ciprofloxacin, and colistin was determined with E-test (AB Biodisk).
Characterization of the involved integrons
The search for integrons of classes 1, 2, and 3 that potentially contain β
Characterization of genetic elements carrying the transferable β-lactamases
B203(B), 204(B1), and 208(D) were the only clinical isolates that were shown to have plasmids. Electroporation of the plasmid DNA in P. aeruginosa PAO1 was carried out. 28 The transformants were selected on LB agar plates and added together with ceftazidime at a concentration of 16 mg/L. The analysis of resistance transfer in the transformed clones was tested by E-test (AB Biodisk) for piperacillin, piperacillin/tazobactam, ceftazidime, cefepime, imipenem, meropenem, gentamicin, tobramycin, amikacin, and ciprofloxacin, interpreting the results according to CLSI. β-lactamase phenotypic tests were conducted as previously described.
A series of hybridization analyses was performed to determine the localization of genes encoding β-lactamases in the bacterial genome. Southern hybridization analyses were carried out according to the manufacturer's instructions (GE Healthcare), utilizing as hybridization probes internal fragments of blaGES, or blaVIM and genomic or plasmid DNA obtained through Genopure Plasmid Maxi kit (Roche Diagnostics) from clinical isolates and recombinant clones.
Results
Molecular typing and susceptibility to antibiotics
PFGE analysis allowed grouping the 14 MDR clinical isolates of P. aeruginosa into four types designated A to D (containing from one to six isolates each); and four subtypes: A1, A2, B1, and C1 (Fig. 1). Eleven isolates showed resistance to nearly all (≥20/21) of the antibiotics tested and were distributed among the four PFGE types. Two isolates (from clusters B and D) demonstrated the ESBL phenotype by DDST, whereas the MBL E-test yielded a positive result in seven isolates (Table 2).

Analysis of Pseudomonas aeruginosa clinical isolates by pulsed-field gel electrophoresis (PFGE).
The highlighted text indicates strains representing each of the four clonal groups.
N.D., not detected; PFGE, pulsed-field gel electrophoresis; ESBL, extended-spectrum β-lactamases; MBL, metallo-β-lactamase; AMK, amikacin; AZL, azlocillin; AZT, aztreonam; CAR, carbenicillin; CAZ, ceftazidime; CEF, cefoperazone; CIP, ciprofloxacin; CTX, ceftizoxime; CTZ, ceftriaxone; FEP, cefepime; IMP, imipenem; LEV, levofloxacin; MER, meropenem; MEZ, mezlocillin; NET, netilmicin; NOR, norfloxacin; OFX, ofloxacin; PIP, piperacillin; TIC, ticarcillin; TIC/CLAV, ticarcillin-clavulanate; TOB, tobramycin.
Characterization of acquired β-lactamases
Strains 212(A2), 204(B1), 206(C), and 208(D) were selected as representative of each PFGE clonal type to detect the production of β-lactamases by the IEF technique. Two bands with a pI of 5.8 and 7.7 suggested the presence of GES-5 and OXA-2 β-lactamases, respectively. Since no pI corresponding to MBLs could be observed, we decided to do specific MBL gene amplification on all the strains. The search was positive for the MBL gene blaVIM for seven isolates, and each blaVIM fragment was sequenced either directly or from recombinant E. coli clones. An analysis of the corresponding amino-acid sequences revealed the presence of VIM-2 and VIM-11 variants (Fig. 2). Interestingly, two strains showed both variants of VIM. Moreover, blaGES-5 was detected by PCR and sequencing in 11 strains, as well as blaOXA-2, which was present in four strains (Table 2).

Comparison of a 168 amino-acid region of the lactamase B superfamily domain of VIM from seven clinical isolates of P. aeruginosa. The reference sequences to compare with were VIM-2 (ACV91880) and VIM-11 (ABL61511). The amino-acid conservation identities are plotted to a VIM-2 as a dot, as well as by numbering. Reference and clinical isolates sequence titles are shown on the left. P. aeruginosa isolates present VIM-2 and VIM-11.
Characterization of the integrons involved
Only six strains of P. aeruginosa showed class 1 integrons, carrying mainly the gene cassettes encoding the resistance to aminoglycosides. However, blaVIM was not detected. The blaGES-5 and blaOXA-2 genes detected in diverse integrons presented a different arrangement. The organization of integrons in 173(A1), 212(A2), 206(C), and 210(C) showed an identity with intI1 [integrase], aacA7 (GenBank accession number: AM988778) [resistant to aminoglycosides], and qacΔE1 [codifying resistance to quaternary ammonium salts]; the integron 204(B1) contained genes intI1, aadA6 (GenBank accession number: FJ752628), and qacΔE1; organization of the integron in 208(D) presented gene sequences corresponding to intI1, aacA4 (GenBank accession number: GQ202693), aadA7 (GenBank accession number: DQ899757), blaOXA-2 [resistant to oxacillin], and qacΔE1 (Fig. 3).
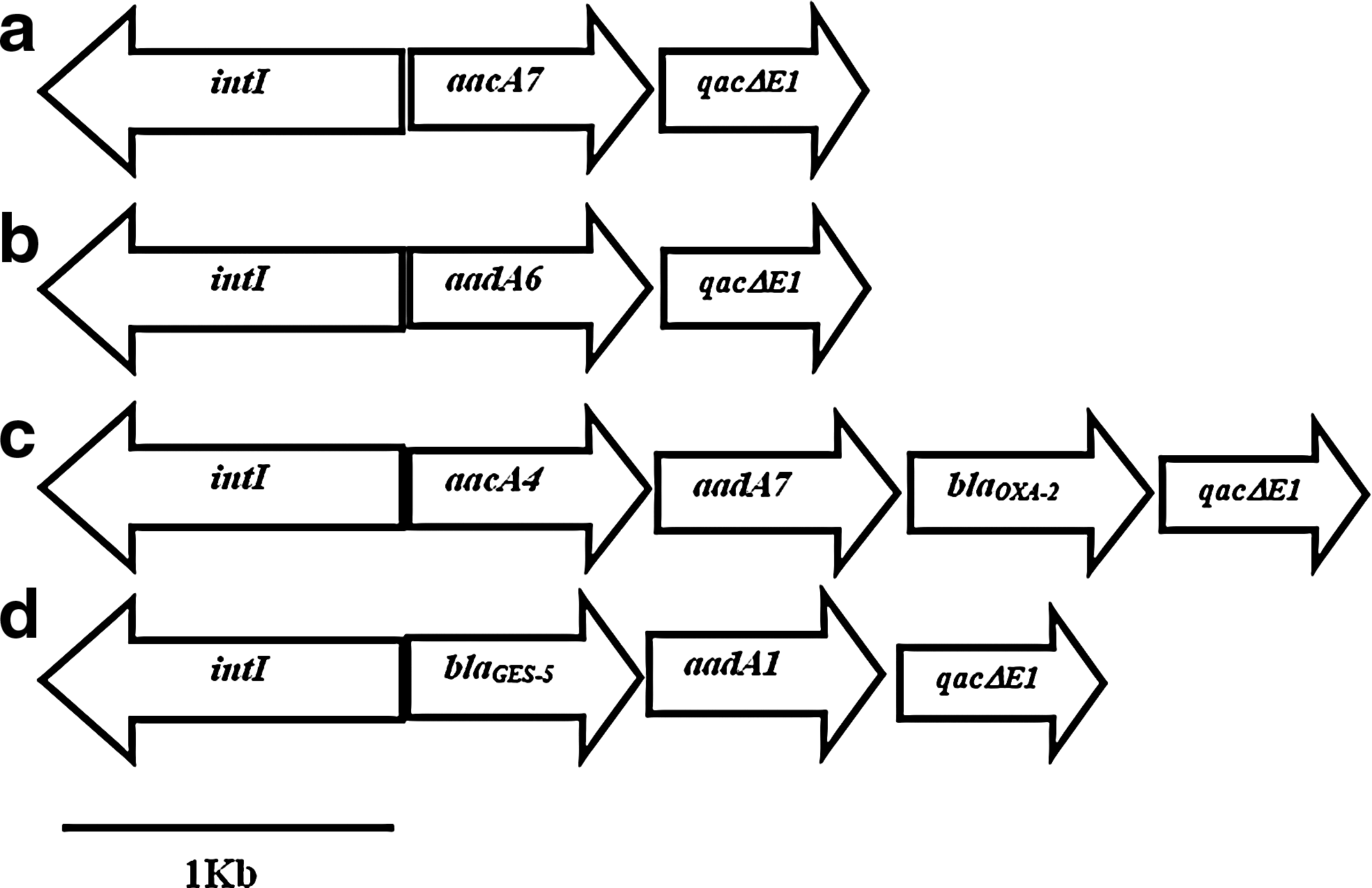
Schematic representation of the class 1 integrons identified in strains of P. aeruginosa.
To study the genetic regions associated with the resistance, total DNA obtained from the isolates 173(A1), 212(A2), B203(B), 204(B1), 206(C), 210(C), and 208(D) was digested with BamHI and EcoRI and ligated to pUCP24 for cloning in E. coli; however, we only obtained the recombinants pUC203B and pUC204B, with both a DNA insert of ∼3.4 kb and an ESBL-positive MBL-negative phenotype (Table 3). The recombinant plasmids were used as templates for the search for class 1 integrons. After amplification and sequencing of pUC204B, a class 1 integron was identified as carrying genes intI1, blaGES-5, aadA1, and qacΔE1.
Cutoff values (μg/ml) for antibiotics: CAZR≥32; FEPR≥32; PIPR≥128; PIP-TZR≥128; IMPR≥16; MEMR≥16; CIPR≥4; COLR≥8; TOBR≥8; AMKR≥64. GEN was the selection marker.
MIC of CAZ, FEP, PIP, PIP-TZ, IMP, MER, CIP, COL, GEN, TOB, AMK, were determined by E-test.
MIC, minimal inhibitory concentration; PIP-TZ, piperacillin-tazobactam; CIP, ciprofloxacin; COL, colicistine; GEN, gentamicin.
blaGES and blaVIM distribution in MDR strains of P. aeruginosa
P. aeruginosa PAO1 was electroporated with native plasmids to obtain three recombinants B203PA, 204PA, and 208PA, which showed a positive phenotype for ESBL, and a negative phenotype for MBL, in addition to presenting resistance to ceftazidime and the transfer of other resistance markers, and susceptibility to imipenem (Table 3).
A Southern hybridization assay was performed to localize blaGES in the chromosome and/or plasmid transferred to P. aeruginosa PAO1. Eleven strains previously positive by PCR to blaGES-5 demonstrated a chromosomal location; in addition, the study on native plasmids obtained from B203(B), 204(B1), and 208(D) confirmed the plasmidic localization of the integrons. On the other hand, blaVIM was found to have chromosomal localization in all the strains (Fig. 4).

Location of the genes of the lactamases VIM and GES in total and plasmid DNA in strains of P. aeruginosa.
Discussion
Typing of P. aeruginosa by PFGE led to the identification of four different types circulating in the Intensive Care Unit for a period of 3 months, showing the coexistence of different strains within a short period, which is similar to reports of outbreaks by MDR P. aeruginosa in other countries. 8
A plasmid containing a class 1 integron with a cassette containing the blaGES-5 sequence was isolated from the 204PA transformant clone type. In addition, in the same clinical strain-type source, 204(B1), a copy of this gene, was found in the chromosome, suggesting that blaGES-5 could be found at first to be associated with a plasmid and subsequently integrated into the bacterial chromosome. The blaGES-5 gene has been reported in South America 20 and Canada, 22 thus the results presented here indicate an expansion of the presence of this gene to another country of the Americas.
The VIM-2 MBL was first described in P. aeruginosa in Europe by Poirel et al. in 2000 23 and since then, there have been reports in different regions of the world. 31 In Mexico, the detection of the blaVIM-2 gene in isolates of P. aeruginosa was documented in 2009 11 and 2010. 24 On the other hand, the VIM-11 MBL has undergone a very different distribution, because it was first described in Argentina by Pasteran et al. in 2005 19 and to date, there is solely one additional report of this type of MBL in strains of P. aeruginosa collected in India. 3 Interestingly, in two of the seven strains, more than one blaVIM variant was observed, which included a blaVIM-2 and a blaVIM-11. To date, P. aeruginosa strains coproducing two variants of VIM β-lactamase have only been reported by Bogaerts et al. in 2008. 2
The β-lactamase genes are mainly associated with class 1 integrons localized in plasmids or chromosomes. This study did not detect blaVIM genes linked with integrons and plasmids; however, to rule out the presence of blaVIM genes in these elements, it would be necessary to determine the regions flanking these genes in the P. aeruginosa strains studied. Nevertheless, and exceptionally, resistant cassettes could be found without a transmissible element, as has been shown by Toleman et al., 31 who demonstrated the absence of a transmissible element in upstream or downstream regions of the blaSPM-1 gene.
The presence of bacteria carrying ß-lactamases that can cause outbreaks in hospitals in Mexico appears to be an emerging problem. In a previous study, Garza-Ramos et al. 11 reported the isolation of VIM-2 and OXA-2 β-lactamases in P. aeruginosa from the hospital environment. However, in this study, the simultaneous presence of the genes blaOXA-2, blaGES-5, blaVIM-2, and blaVIM-11 is shown. The isolates of this study were recovered in 2004, which is close to 2005 when Pasteran et al. 19 recovered strains with the gene blaVIM-11 in Argentina. The results indicate that β-lactamase-carrying bacteria have been circulating in Mexico since several years.
To our knowledge, this is the first study that reports the presence of blaGES-5 in P. aeruginosa strains from Mexico and Central America, in addition to the particular finding that several P. aeruginosa clinical strains simultaneously produced blaVIM-2 and/or blaVIM-11.The coexistence of several and variable β-lactamases in a single strain is, to our knowledge, reported here for the first time, adding further complexity to the epidemiology of transferable resistance in P. aeruginosa. In addition, the detection of two VIM variants through the approach followed suggests the possibility that this circumstance might be more common than thought, because the detection of a VIM variant may mask the presence of other, additional β-lactamases. It is important to highlight the epidemiological risk posed by these strains, due to the potential spread of resistance genes, especially when these strains possess β-lactamases of a different family that can be transferred horizontally.
Footnotes
Acknowledgments
J.C.-V. received a graduate studies fellowship and support to carry out a research stay at Hospital Universitario Son Dureta, Palma de Mallorca, Spain, from CONACyT, México and I.P.N., México. R.M.R.-A. and G.A.-O. are recipients of COFAA-IPN and EDD-IPN fellowships. This work was supported by SIP-IPN-México grants 20091358 and 20101151. C.J. and A.O. are supported by the Ministerio de Ciencia e Innovación of Spain and Instituto de Salud Carlos III, through the Spanish Network for the Research in Infectious Diseases (REIPI C03/14 and RD06/0008).
Disclosure Statement
No competing financial interests exist.