
Abstract
The aim of this study was to investigate the fitness of macrolide resistant Campylobacter coli and Campylobacter jejuni. The in vitro growth, the survival on food matrix, and the in vivo colonization of C. jejuni and C. coli susceptible isolates and their isogenic resistant mutants were studied. In vitro experiments demonstrated that macrolide resistance imposed a fitness cost when the susceptible strains and their isogenic resistant mutants were cultured in competition. When inoculated in food matrix, the resistant C. jejuni mutant was no longer detectable after 3 to 5 days but the susceptible strain remained detectable for over 18 days. No difference in survival in food matrix was observed between susceptible and resistant C. coli. When inoculated in vivo in chickens, the macrolide susceptible and resistant C. coli displayed similar levels of colonization, both in separated inoculations and during competitive assays. Strikingly, when mono-inoculated or co-inoculated into chickens, macrolide susceptible C. jejuni outcompeted the macrolide resistant population. However, a spontaneous mutant that evolved in vivo showed a colonization capacity similar to the susceptible strain. Our findings demonstrate the effect of macrolide resistance on the fitness of Campylobacter but suggest that evolved mutants may be as fit as susceptible strains.
Introduction
Apart from human use, macrolides are also used as therapeutic agents for food-producing animals, which also contributes to the selection of macrolide resistant Campylobacter. 13 The spread of antibiotic resistant bacteria is not only affected by environmental factors or usage of antibiotics but is also influenced by the relative fitness of the resistant microorganism. The stability and the impact of macrolide resistance on Campylobacter fitness, although critical and crucial to our understanding of resistance evolution, are still largely unknown especially in in vivo conditions. It is generally thought that the acquisition of antimicrobial resistance imposes a fitness cost. However, this decrease in fitness may be restored by compensatory mutations without loss of resistance, rendering bacterial populations as fit as the susceptible bacteria. 3 In this study, we investigated the fitness cost of macrolide resistance in Campylobacter coli and in Campylobacter jejuni. As previously studied for fluoroquinolone resistance, 21 the effects of macrolide resistance were examined in vitro in food matrix and in vivo in the chicken digestive tract, which is the natural reservoir of Campylobacter.
Materials and Methods
Bacterial isolates and growth conditions
Two strains previously isolated from healthy chicken caeca, C. coli Cc454 and C. jejuni Cj126, were used as wild-type (WT) susceptible strains for selection of erythromycin (ERY) resistant mutants. Strains had been identified to the species level by polymerase chain reaction (PCR). 9 Strains were routinely cultivated on Mueller-Hinton agar (MHA) plates or in Mueller-Hinton broth (MHB) (Biorad, Marnes la Coquette, France) under microaerobic conditions (85% N2, 10% CO2, and 5% O2) at 42°C. When necessary, the medium was supplemented with ERY (Sigma, St Quentin Fallavier, France) at a final concentration of either 4 mg/L for C. jejuni or 16 mg/L for C. coli.
In vitro selection of macrolide resistant mutants
Macrolide resistant mutants were selected as described previously. 6 Briefly, the WT strains were grown on increasing concentrations of ERY. Mutants were selected on MHA plates supplemented with 160 mg/L ERY and 5% sheep blood. After 48 hr of incubation under microaerobic conditions at 42°C, single resistant colonies were selected, cultured on nonsupplemented MHA plates, and stored at −70°C.
Antimicrobial susceptibility test
Minimal inhibitory concentrations (MICs) were tested using the agar dilution method as recommended by the Clinical Laboratory and Standards Institute (CLSI), M31-A3. 8 According to EUCAST (European Committee on Antimicrobial Susceptibility Testing), 11 C. coli strains with MIC(ERY)>8 mg/L and C. jejuni strains with MIC(ERY)>4 mg/L were considered to be resistant to ERY. MICs of tylosin, azithromycin, lincomycin, ciprofloxacin, ampicillin, and tetracycline were also determined for the parent and their mutant strains.
Characterization of the parent and mutant strains
Operon-specific PCR amplification of the three copies of 23S rRNA gene was performed for C. coli and for C. jejuni using primers listed in Table 1. For C. coli, a small internal fragment of domain V of the 23S rRNA gene was amplified using the primer pair 23Scc1–23Scc2. 12 Genes encoding L4 and L22 ribosomal proteins in C. coli and C. jejuni were amplified by PCR using the primers pairs PL4CJ1-PL4CJ2 and PL22CJ1-PL22CJ2, respectively. All PCR products were purified and sequenced as described previously. 21
In vitro competitive assay and survival on food matrix
Growth curves and survival on food matrix (chicken skin fragments) of the susceptible strains and the ERY resistant mutants were obtained separately and by competition between WT and mutant, as described previously. 21
In vivo experiments
Three independent experiments were conducted with 10-day-old Specific Pathogen Free (SPF) Campylobacter spp.–free chickens. The experimental design is detailed in Table 2. The susceptible and the resistant strains were inoculated separately or simultaneously in competition or one strain was inoculated four days before or four days after the other, to evaluate whether the presence of the resistant mutant in the digestive tract could prevent colonization by the susceptible strain or vice versa. Birds were inoculated via oral gavage with 100 μl of an inoculum suspension adjusted to 108 colony forming units (CFU)/ml. After inoculation, individual fecal samples were collected at different times during four weeks from the 10 inoculated chickens in each isolator and from all the contact animals. Samples were serially diluted to 1:10 in MHB, and were cultured following direct plating and enrichment. For direct plating, 100 μl of the appropriate dilutions was plated in duplicate onto MHA with Campylobacter selective agents and onto MHA with Campylobacter selective agents and ERY to detect macrolide resistant Campylobacter colonies. After enrichment in MHB for 24 hr at 42°C, a 100 μl sample of each broth culture was streaked onto MHA plates with Campylobacter selective agents and onto MHA plates with Campylobacter selective agents and ERY. Plates were incubated for 48 hr under microaerobic conditions at 42°C.
CFU, colony forming units.
For the third experiment, three groups of 10 chickens were used. All the birds were mono-inoculated with Cj126, Cj126m, or Cj126m2 (a clone reisolated from a bird during the second experiment). Inoculation, sampling, and cultures were performed as described previously. Assays were performed according to the French animal welfare regulations (Authorization B-22-741-1 decisions 10/04/13-07 and 10/11/09-04).
Statistical analysis and quantification of transmission during in vivo experiments
In the in vivo experiments, when only the enrichment culture was positive, the samples were considered to contain 100 CFU/g of feces, corresponding to the detection limit of the plating method. The geometric mean values of the titers of the WT and mutant strains were calculated and compared using Student's t-test or the Mann and Whitney test depending on the normality of distributions and homogeneity of variances. Differences between the mean values were considered significant at p<0.05.
A susceptible–infectious–susceptible (SIS) type model was used to compare the transmission rate parameter β of the susceptible and the resistant strains. 17
Sequence analysis
Nucleotide sequences were aligned using the Web site http://bioinfo.genotoul.fr/multalin/. C. coli or C. jejuni strain sequences were aligned using C. coli ATCC 33559 (accession no. GQ167698.1) or C. jejuni NCTC 11168 (accession no. AL111168.1). To further analyze the sequences in C. jejuni, the Needle and Water programs from the EMBOSS (European Molecular Biology Open Software Suite) package were used for global and local alignments. 20 Local alignment was used to correlate the positions of the mutations between C. jejuni and Escherichia coli (accession no. J01695) sequences. The canonical secondary structures of 23S rRNA were downloaded from the comparative RNA Web site. 7 The analysis of structural models of ribosome was carried out with the Pymol visualization software. 1 The coordinates were downloaded from the Protein Data Bank (www.pdb.org): PDB ID codes 3OFO, 3OFP, 3OFR, 3OFQ (70S ribosome in complex with ERY). 10
Results
Characterization of Campylobacter WT strains and mutants
MICs of WT and isogenic mutants are shown in Table 3. MICs of ciprofloxacin, ampicillin, and tetracycline did not differ between WT and resistant mutants, whereas MICs of ERY, tylosin, azithromycin, and lincomycin of mutants were increased.
MIC, minimum inhibitory concentration; MICs determined by the agar dilution method. ERY, erythromycin; TYL, tylosin; AZM, azithromycin; LIN, lincomycin; CIP, ciprofloxacin; AMP, ampicillin; TET, tetracycline.
No mutation was detected in the macrolide susceptible strains C. coli and C. jejuni for all sequenced genes. Resistant mutant C. coli harbored the A2075G mutation in each of the three 23S rRNA gene copies but no modifications were observed in rplD or rplV genes. The C. jejuni mutant harbored the A2074C mutation in one copy encoding 23S rRNA (rrlB) out of the three of the 23S rRNA genes and the G221A mutation in the rplD gene. No modifications were observed in the rplV gene.
Evaluation of spontaneous mutation or reversion
Spontaneous macrolide resistant mutants arose only at a very low mean frequency of less than 8.27×10−8 for C. coli Cc454 and less than 2.54×10−8 for C. jejuni Cj126. The frequency of colonies with spontaneous resistance mutations was thus considered to be negligible.
Likewise, to determine whether the number of macrolide susceptible colonies corresponded to the inoculated WT strains or to a mixture of inoculated WT cells and revertants, cultures were always spread in parallel on MHA and on MHA with ERY plates. Given that the number of colonies on the two types of plates was not significantly different, the reversion of resistant strains to susceptibility was considered to be negligible.
Effect of macrolide resistance on the stability and the fitness of C. coli and C. jejuni in vitro
Growth kinetics followed for 48 hr were similar between the WT strain and its mutant either for C. coli or C. jejuni, either in separated or competitive cultures. To assess slight differences in the growth of the susceptible strain and the mutant, sequential passages were conducted. For C. coli and C. jejuni, no detectable differences in growth rates were observed in separated cultures (data not shown). However, in the mixed culture over a long period of time, the population of the mutants clearly decreased in comparison to the susceptible population for C. coli and C. jejuni. As shown in Figure 1, after eight sequential passages, the ratio of CFU/ml (mutant) to CFU/ml (WT) decreased up to 103-fold for C. coli and up to 106-fold for C. jejuni. The internal fragment of the three copies of the 23S rRNA gene as well as rplD and rplV genes was sequenced for 10 C. coli and 10 C. jejuni clones isolated from the fourth and the eight mono-inoculated culture passages. The results revealed identical sequences for all collected isolates and inoculated mutants. MICs of ERY, tylosin, azithromycin, lincomycin, ciprofloxacin, ampicillin, and tetracycline remained unchanged.

In vitro population dynamics of the susceptible strains Campylobacter coli 454 and Campylobacter jejuni 126 and their resistant mutants (454 m and 126 m, respectively) in mixed cultures. Results were expressed as a ratio of CFU (mutant)/CFU (WT) at each passage point. Light gray line: CFU (Cj126m)/CFU (Cj126); dark gray line: CFU (Cc454m)/CFU (Cc454). WT, wild type; CFU, colony forming units.
Effect of macrolide resistance on the survival of C. coli and C. jejuni in food matrix
At 5°C±3°C, no significant differences between the titers of the macrolide susceptible strain and its respective mutant were detected for C. coli either in separated inoculation or in competition. Cc454 and Cc454m remained detectable for more than 18 days. However as shown in Figure 2A, when inoculated separately or in competition, C. jejuni Cj126 was still detected after 18 days on food matrix whereas the corresponding mutant Cj126m was no longer detected after only 5 days or 3 days, respectively, when inoculated separately or in competition (Fig. 2B).

Comparison of macrolide susceptible (Cj126) and macrolide resistant (Cj126m) C. jejuni survival in food matrix at 5°C±3°C.
Experiment I: Effect of macrolide resistance on the fitness of C. coli in vivo
Five days after inoculation, individual titers of C. coli WT strain (Cc454) and its resistant mutant (Cc454m) varied among chickens between 105 and 108 CFU/g of feces and all birds were colonized. No significant differences were observed in the mean levels of Cc454 and Cc454m when mono-inoculated, or when co-inoculated, simultaneously or successively into animals. In chicken hosts placed with mono-inoculated birds, the values of the transmission rate parameter β for Cc454 and Cc454m were, respectively, 2.4 [0.84–6.89] and 2.4 [0.86–6.98]. In chicken hosts placed with co-inoculated broilers, the parameter β for both Cc454 and Cc454m was 1.42 [0.53–3.75]. These results indicate similar colonization (i.e., number of colonized chickens and level of excretion) and transmission for macrolide susceptible and resistant C. coli strains.
The internal fragment of the three copies of the 23S rRNA gene as well as rplD and rplV genes was sequenced for 10 clones collected 28 days post-inoculation from 10 different Cc454m mono-inoculated animals. The analysis of sequences in the representative isolates revealed identical sequences. All MICs remained unchanged. Macrolide resistance resulting from the A2075G substitution in C. coli was found to be stable in the chicken digestive tract.
Experiment II: Effect of macrolide resistance on the fitness of C. jejuni in vivo
A significantly higher ratio of culture-positive samples was obtained in birds inoculated with the susceptible strain (67/82) compared with birds inoculated with the resistant one (60/89). As shown in Figure 3A, significant differences (p<0.001) in the colonization level between Cj126 and Cj126m were observed in mono-inoculated animals with higher titers for the WT strain Cj126 up to 23 days after inoculation. Similarly, in animals co-inoculated, the macrolide susceptible population outcompeted the macrolide resistant Campylobacter (Fig. 3B). This difference in colonization level was determined in terms of CFU/g of feces, as well as in terms of the number of chickens colonized by both strains.
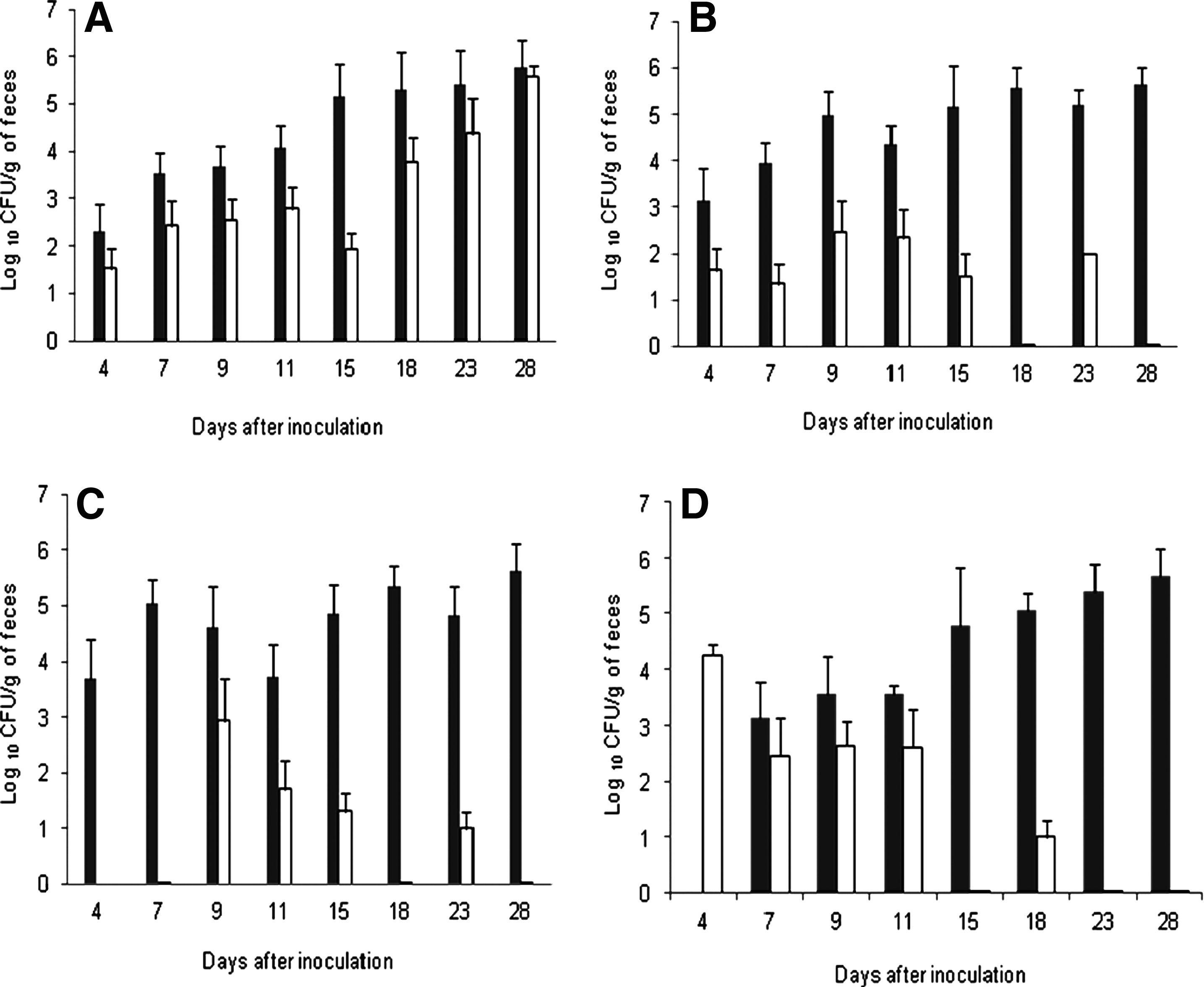
Experiment II: Colonization of macrolide susceptible and resistant C. jejuni in the chicken host.
When SPF chickens were inoculated with the susceptible strain 4 days before or after the resistant mutant (Fig. 3C, D), a significant difference was observed between the colonization of Cj126 and Cj126m (p<0.001), regardless of the order of inoculation. Fifteen days after inoculation, mean titers of Cj126m remained between 101 and 102 CFU/g of feces, whereas mean titers of Cj126 were greater than 104 CFU/g of feces and reached 106 CFU/g of feces 28 days after inoculation.
In chickens placed with mono-inoculated animals, the values of parameter β for Cj126 and Cj126m were, respectively, 0.7 [0.28–1.77] and 0.53 [0.28–1.03]. In chickens placed with co-inoculated animals, the values of parameter β for Cj126 and Cj126m were, respectively, 0.65 [0.3–1.43] and 0.97 [0.58–1.6].
However, 23 to 28 days post-inoculation, when mono-inoculated into the host, the susceptible and the resistant C. jejuni had approximately the same titers (p>0.05). To better understand whether the absence of a fitness cost at the end of the experiment was due to compensatory mutations, the internal fragment of the three copies of the 23S rRNA gene as well as rplD and rplV genes was sequenced for 10 clones randomly collected 28 days post-inoculation from the 10 Cj126m mono-inoculated animals. Eight isolates presented a new mutation in one copy of the 23S rRNA gene (rrlB) without losing the mutations conferring macrolide resistance. Interestingly, this new acquired mutation was found in the same copy of the mutated 23S rRNA gene. No changes were detected in MICs of ampicillin, tetracycline, and ciprofloxacin in any of these clones. However, the level of macrolide MICs in all collected clones decreased very slightly, but the clones were still resistant to macrolides. The mutation C2551G in rrlB was also present in 25%, 66%, and 77% of the isolates collected, respectively, at 15, 18, and 23 days post-inoculation. These isolates conserved their resistance to macrolides and no changes were observed for MICs of ampicillin, tetracycline, or ciprofloxacin (Table 3). The mutation C2551G in rrlB was not observed in 10 isolates obtained from Cj126 inoculated chickens 28 days after inoculation
To further understand the effect of the C2551G mutation, this substitution was localized on the 70S ribosome. In comparison with E. coli, the crystalline structure of the 70S ribosome showed that the C2551G mutation lies in the vicinity of the L16 ribosomal protein. However, a distance of ∼60 Å appeared between the C2551G and A2074C mutations. These findings suggest that there is no co-relation between C2551G and ERY resistance, but enhanced ribosomal activity may be present due to the impact of C2551G on L16 protein.
Experiment III: Colonization and morphology of Cj126m2
To assess the in vivo fitness of one of the reisolated clones (Cj126m2) harboring the A2074C and C2551G mutations in the 23S rRNA gene (rrlB) and the G221A mutation in the rlpD gene, three isolators were used with 10 SPF chickens in each. Animals were mono-inoculated with the isolate Cj126m2, the susceptible strain Cj126, or the resistant mutant Cj126m as described previously. As previously observed and as shown in Figure 4, titers of Cj126 were higher than those of Cj126m (p<0.01) during the first days, but Cj126m2 and Cj126 demonstrated similar levels of colonization (p>0.05). The rrlB gene was sequenced for 10 clones collected 23 days post-inoculation from animals inoculated with the mutant Cj126m. Again, results revealed the presence of the mutation C2551G with the substitution A2074C. Finally, during in vivo experiments, strains Cj126 and Cj126m2 exhibited the same typical colony morphology on the plates obtained from fecal samples, whereas the mutant Cj126m formed smaller colonies compared with its respective parental isogenic strain Cj126.
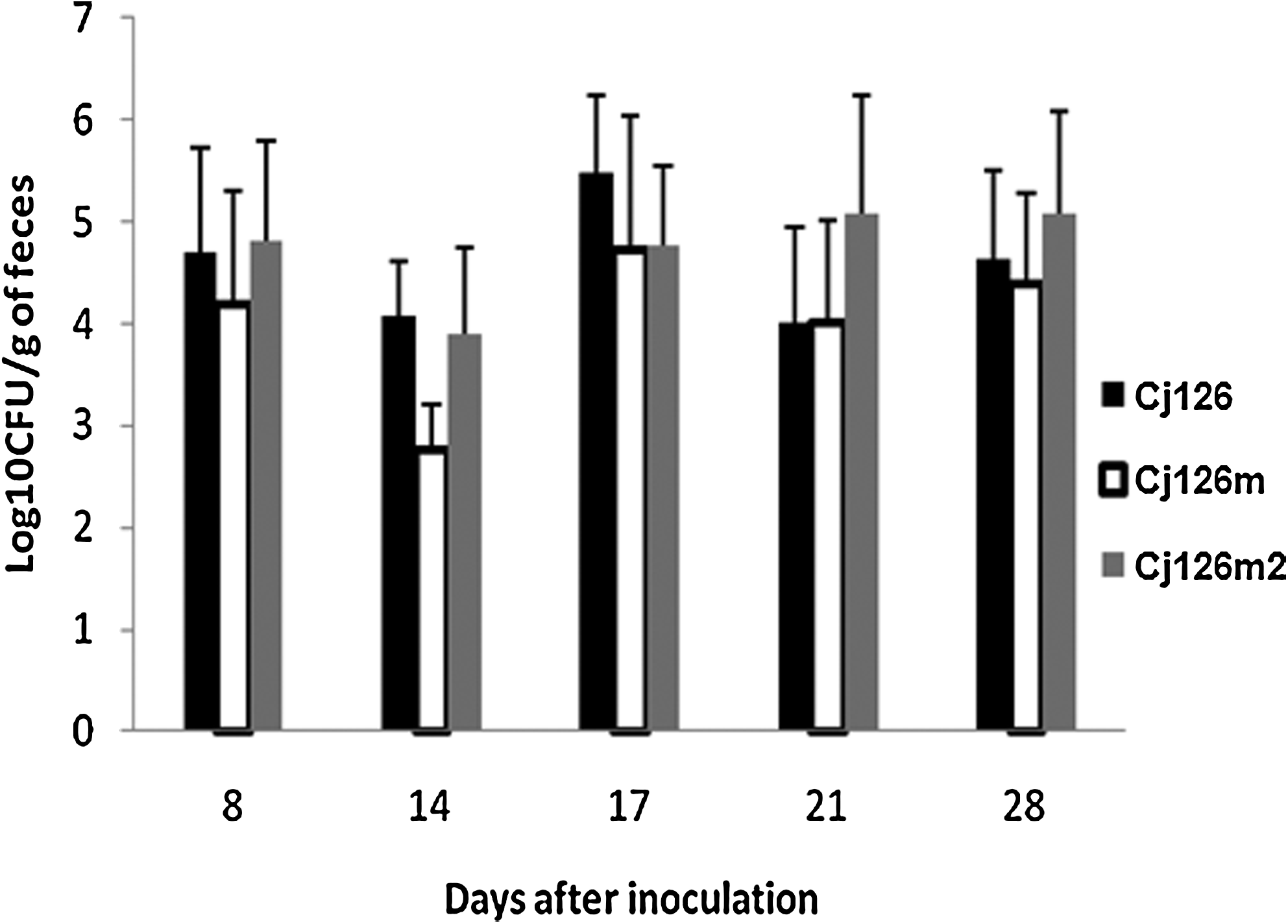
Colonization of macrolide susceptible, resistant, and evolved mutant C. jejuni in the chicken host. Animals were inoculated with either Cj126 (black bars), Cj126m (white bars), or Cj126m2 (gray bars). Each bar represents the arithmetic mean shown with its standard deviation of log10 CFU/g of feces.
Discussion
To our knowledge, this work represents the first report documenting the fitness cost of macrolide resistant Campylobacter in food matrix as well as in vivo, in the animal reservoir. However, the fitness cost of fluoroquinolone resistant mutants of C. coli (strain 103) and C. jejuni (strain 126) had already been studied under such conditions. Co-inoculation of fluoroquinolone resistant and susceptible strains on food matrices or in chickens showed that fluoroquinolone resistance induced a biological cost for these isolates. 21
The two mutants obtained in our study were stable in vitro up to the final eighth passage. Mutations harbored by selected mutants have already been described.5,6,12 Hao et al. 15 and Caldwell et al. 6 showed that mutants of low-level ERY resistance were not stable in vitro or in vivo in the absence of macrolide selection pressure. While the most highly resistant mutants with 23S rRNA gene mutations have been shown to be stable in vitro,12,15,16 an A2074G resistant mutant reverted to ERY susceptibility after 55 subcultures on ERY-free medium. 12
In our study, as showed by Han et al. 14 and Hao et al., 15 no fitness cost was observed when WT strains and their mutants were inoculated in separated broth cultures. It remains to be elucidated whether the fitness cost in our separated broth cultures is truly nonexistent, or whether it is so low that it could not be measured with current laboratory methods. However, in agreement with recent studies performed on C. jejuni14,15 and after conducting sequential passages on mixed cultures, we revealed that macrolide resistance imposed a significant burden on bacterial growth competition either in C. coli or in C. jejuni. Likewise, Almofti et al., 2 Hao et al., 15 and Han et al. 14 showed an in vitro fitness cost of macrolide resistance in C. jejuni in relation to the A2074C and A2075G mutations, respectively, although the fitness cost associated with alterations in only one mutated copy of the 23S rRNA gene and one mutation in rplD gene was not known.
Given that Campylobacter is more prevalent in chicken products, skin fragments of SPF chickens were chosen as a model. In our conditions, macrolide resistance had no impact for C. coli but resulted in a rapid disappearance of C. jejuni resistant mutant. When inoculated in food matrix, WT and resistant mutants can be exposed to various oxidative, thermal, or starvation stresses. Because ribosomal activity is altered in the resistant mutant, the expression of many genes implicated in stress regulation may affect the viability of the resistant strain in food matrix. 19 The difference in results obtained between the two species C. coli and C. jejuni can be attributed to the diverse mutations harbored by their respective mutants.
For C. coli, no difference in colonization or diffusion between the WT and the resistant mutant was observed in vivo. For C. jejuni, the different inoculation schemes showed that the resistant mutant was clearly a weaker colonizer compared with the parental strain. However, during the course of the experiment, a new isolate Cj126m2 arose and appeared more competitive than Cj126m, but still resistant to macrolides. The molecular analysis showed that they had an additional mutation in the 23S rRNA gene. The same additional mutation also appeared during the second in vivo experiment using C. jejuni 126m. To our knowledge, this C2551G mutation has never been reported for Campylobacter. The localization of the C2551G mutation does not clearly explain the apparent advantage or the slightly decreased macrolide MIC. However, it is possible that the Cj126m2 clones bear, in other parts of their genomes, other ignored mutations that may explain why the evolved Cj126m2 and the parental Cj126 strains demonstrate similar levels of colonization. On the other hand, the effect of the C2551G mutation on the vicinity of the L16 ribosomal protein may be an advantage for the ribosomal activity. These types of compensatory mutations, which appear in vivo, have already been observed for other bacteria or other antimicrobials 4 such as clarithromycin resistant Helicobacter pylori in mice. These observations underline the fact that resistant mutants can benefit from compensatory mutations acquired in vivo and can thus be stabilized in the population in the absence of antibiotic selection pressure.
Footnotes
Acknowledgments
The authors are grateful to members of the ANSES-SEEPA and to Julien Le Bloas and Nicolas Rose for their precious help. This work was supported by ANSES and the “Conseil Général des Côtes d'Armor.” Salman Zeitouni was financed by the French Ministry of Research.
Disclosure Statement
The authors disclose any commercial associations that might create a conflict of interest in connection with the submitted manuscript.