
Abstract
Forty-three vancomycin-resistant enterococci (VRE) from different patients were recovered in a Spanish Hospital (2003–2010), representing 0.4% of the total of enterococci recovered. Mechanisms detected were vanA (five Enterococcus faecium, two E. faecalis), vanB2 (seven E. faecium, five E. faecalis), vanB1 (one E. faecalis), and vanC1/2 (22 E. gallinarum, 1 E. casseliflavus). Four different Tn1546 structures were found among the seven vanA strains, three of them with insertions (ISEf1 or IS1542) or deletions. Most of the VRE presented a multiresistance phenotype and harbored different resistance genes [erm(B), tet(M), tet(L), ant(6)-Ia, aac(6′)-aph(2′′), aph(3′)-IIIa, and catA]. Sixteen unrelated pulsotypes were detected among the 20 vanA/vanB E. faecalis and E. faecium isolates by pulsed-field-gel-electrophoresis and 11 unrelated pulsotypes among the 22 E. gallinarum isolates. Six different sequence types (ST) were demonstrated among the 12 vancomycin-resistant E. faecium strains (one of them new), and 5 were included into the clonal-complex (CC) CC17. Five different ST were detected among the eight E. faecalis strains. The esp gene was detected in 58% and 25% of E. faecium and E. faecalis strains, respectively, and the hyl gene in 78% and 89%, respectively. A high diversity of clones and genotypes of VRE were detected in this hospital.
Introduction
At present, 10 different genotypes of glycopeptide resistance have been described in enterococci either of acquired (vanA, vanB, vanD, vanE, vanG, vanL, vanM, and vanN), or intrinsic type (vanC1 and vanC2/vanC3, associated with E. gallinarum and E. casseliflavus/flavescens species, respectively). 8 To date, vanA and vanB2 resistance mechanisms have important clinical relevance since they are predominant by far in humans. 14 The vanA and vanB2 mechanisms are mediated by the vanA and vanB2 cluster of genes, respectively, commonly included into conjugative transposons (transposons Tn1546 and Tn5382 or Tn1547, respectively); these transposons can be located on plasmids, frequently in the case of Tn1546 and sporadically in the case of Tn5382, being able to spread widely.6,8,12,31
In the last decade, the analysis of vancomycin-resistant E. faecium and E. faecalis strains by multilocus sequence typing (MLST) allowed their inclusion into predominant clonal complexes (CC) (http://efaecium.mlst.net/, http://efaecalis.mlst.net/).18,21,32 Most vancomycin-resistant E. faecium isolates recovered from nosocomial infections worldwide, belong to the high risk CC (HRCC) CC17.23,35 This CC of E. faecium, which is well adapted to the hospital environment, shows generally ampicillin and quinolone resistance, and frequently contains the esp virulence gene. Some HRCC (CC2 and CC9) have also been detected among vancomycin-resistant E. faecalis strains.16,24
Very few studies have been reported on the prevalence and the population structure of VRE in Spanish hospitals.10,30 Moreover, most studies on VRE are based on the description of outbreaks or on infections produced by VRE in particular types of patients,24,27,37,38 and are not focused on the analysis of isolates over a wide period of time in the last years. The aim of this study was to determine the prevalence of VRE in a Spanish hospital over a period of 8 years (2003–2010), to analyze the resistance and virulence patterns of these isolates and to perform their molecular characterization, to track the evolution of the mechanisms of resistance.
Materials and Methods
Hospital setting and bacterial strains
The Hospital General Universitario Gregorio Marañón of Madrid (Spain) is a tertiary hospital. Over the period of study, the number of beds in this hospital ranged from 1,550 to 1,750 and the population attended ranged from 600,000 to 815,000 inhabitants.
Over the period of study (2003–2010), all enterococcal isolates recovered in the microbiology laboratory were identified and subjected to antimicrobial susceptibility testing using the automated microdilution system MicroScan (Siemens).
All VRE with minimal inhibitory concentration (MIC) for vancomycin ≥8 mg/L, isolated from patients during the 8-year period were included in the study. In addition, all VRE isolates were identified by species-specific PCR. 22 Patients were considered to be infected by VRE if they presented clinical signs and symptoms of infection and when enterococci were isolated from blood (primary or secondary bacteremia, and catheter-related bacteremia), cerebrospinal fluid (CSF), intraabdominal fluids, deep wounds and abscesses, and urine (>104 cfu/ml in the urine culture). VRE were considered to be colonizers in the absence of clinical signs and symptoms of infection and when they were isolated from superficial wounds, catheters (less than 15 cfu/plate) or transport media used to send the samples to the microbiology laboratory (cardiac valves in patients without bacteremia and/or endocarditis).
Pulsed field gel electrophoresis analysis
The clonal relationship of all VRE strains was studied by a macrorestriction analysis by pulsed field gel electrophoresis (PFGE). Briefly, a small slice of agarose plugs prepared with whole-cell DNA was digested with the SmaI enzyme (BioLabs) for 4 hours at 30°C, and loaded into a 1.2% agarose gel (Bio-Rad Laboratories). This technique was performed using clamped homogeneous electric fields (CHEF DRII; Bio-Rad), and the running parameters were as follows: 6 V/cm2 with initial and final pulse times of 5 and 35 seconds, for E. faecium and E. faecalis, and of 5 and 12 seconds, for E. gallinarum and E. casseliflavus, and 24 hours at 14°C for both. Isolates were classified as indistinguishable, closely related, possibly related, or unrelated by following previously defined criteria for bacterial strain typing. 34
Antimicrobial susceptibility testing
As described above, MICs of vancomycin, teicoplanin, and ampicillin were determined using the automated MicroScan system. All isolates with vancomycin MICs of ≥8 mg/L were included in this study. In addition, the disk diffusion method was used to determine the susceptibility to streptomycin (300 μg), gentamicin (120 μg), kanamycin (120 μg), chloramphenicol (30 μg), tetracycline (30 μg), erythromycin (15 μg), and ciprofloxacin (5 μg) following the CLSI guidelines. 9 E. faecalis ATCC 29212 and Staphylococcus aureus ATCC 29213 were used as control strains.
Characterization of vancomycin resistance mechanisms
Vancomycin resistance genes were analyzed using specific primers for amplification of the vanA, vanB, vanC-1, and vanC-2/3 genes, 22 and positive vanA and vanB amplicons were sequenced to confirm the mechanism and also to determine the vanB allele type (vanB1, vanB2, or vanB3).
The possible inclusion of the vanB2 operon into a Tn5382-like element was studied in vanB2-positive isolates by specific PCR assays. The possible presence of insertion sequences in this operon was also investigated amplifying the intergenic vanSB/vanYB region as well as the ISEnfa110 and ISEnfa200 insertion sequences, with specific primers.12,22 vanB2-positive enterococci were also checked for pbp5-Tn5382 linkage by PCR amplification of a 1,079 bp region between pbp5 and Tn5382. 12 All positive PCR amplicons were sequenced to verify results.
The vanA cluster of genes was further characterized in all vanA-positive isolates. The whole structure of Tn1546 was analyzed by overlapping-PCR, using primers previously reported,4,23,29and by sequencing.
PCR detection of other antibiotic resistance genes and virulence genes
The presence of genes encoding resistance to erythromycin [erm(A) and erm(B)], tetracycline [tet(M) and tet(L)], aminoglycosides [ant(6)-Ia, aac(6′)-Ie-aph(2′′)-Ia, and aph(3′)-IIIa], and chloramphenicol [catA] was analyzed by PCR using primers and conditions previously reported. 22 The nucleotide sequence of the pbp5 gene from one selected AMP-resistant E. faecium isolate was studied to detect amino acid changes in the C-terminal region of the penicillin-binding protein 5 (PBP5), associated with ampicillin resistance. 19
The potential presence of genes related with putative virulence factors (esp, hyl, agg, ace, and gelE) was also investigated by PCR using previously described primers.11,13,23 Positive and negative controls from the collection of strains of the University of La Rioja were included in all PCR assays.
Assay of gelatinase activity
Gelatinase production was detected by inoculating enterococci onto freshly prepared Tryptic Soy Agar (Becton, Dickinson and Company) plates containing 1.5% skin milk. A clear halo around colonies after overnight incubation at 37°C was considered as positive gelatinase production. 3
Multilocus sequence typing
One isolate per patient and per PFGE pattern, belonging to the E. faecium or E. faecalis species, was selected for further characterization by MLST. For this purpose, internal fragments of seven housekeeping genes (atpA, ddl, gdh, purK, gyd, pstS, and adk for E. faecium; gdh, gyd, pstS, gki, aroE, xpt, and yiqL for E. faecalis) were amplified and sequenced. The sequences obtained were analyzed and compared with those included in the website databases (http://efaecium.mlst.net/and http://efaecalis.mlst.net/), and a specific sequence type (ST) and CC was assigned.18,32
Results
Epidemiology
Fifty-six VRE were recovered, belonging to 43 different patients, among a total of 13,362 enterococci (10,578 E. faecalis, 2,540 E. faecium, and 244 of other species) isolated in the microbiology laboratory (0.42%) over the study period. All isolates belonging to the same patient showed the same PFGE profile, then, only one isolate per patient was further characterized in this study (43 VRE). The origin of the isolates is shown in Table 1. Thirteen patients (30%) with VRE showed infection, and 30 (70%) were colonized (Table 1). Species identification of the 43 VRE isolates showed 12 E. faecium (28%), 8 E. faecalis (19%), 22 E. gallinarum (51%), and 1 E. casseliflavus (2%).
Efcalis, PFGE pattern of E. faecalis; Efcium, PFGE pattern of E. faecium; Egal, PFGE pattern of E. gallinarum.
Infection/colonization.
High-level resistance to kanamycin, gentamicin, and streptomycin.
VRE was recovered from other specimens in this patient, with similar genetic characteristics to the one recorded in this table.
The symbol - indicates that no virulence genes or gelatinase activity was detected.
PFGE, pulsed field gel electrophoresis; ST, sequence types; CC, clonal complexes; ND, not determined.
Clonal relationship
Table 1 shows the results of clonal relationships by the PFGE analysis among the studied strains. Eight different pulsotypes were found among the eight E. faecalis strains (named as Efcalis-A to Efcalis-H). In addition, eight different patterns were also identified among the 12 E. faecium strains (named as Efcium-A to Efcium-H), with one closely-related pattern (Efcium-F1).
The 22 E. gallinarum isolates were resolved into 11 different pulsotypes (named as Egal-A, to Egal-K) (Table 1). One pulsotype, Egal-A, was detected in 11 strains, however, the isolates corresponded to different patients admitted in different wards of the hospital in different years without any epidemiological relationship among them (Table 1). Six isolates recovered from cardiac valves corresponded to a possible contamination of the transportation fluid used to send the samples to the laboratory. 28
Resistance phenotype and genotype
All E. faecium and E. faecalis isolates, as well as 11 E. gallinarum and 1 E. casseliflavus isolates showed an MIC for vancomycin ≥32 mg/L, and the remaining 11 E. gallinarum isolates exhibited an MIC for vancomycin 16 mg/L. Concerning to teicoplanin, only seven strains (two E. faecalis and five E. faecium) showed resistance to this antibiotic (MIC ≥32 mg/L).
Thirteen of the 43 VRE strains carried a vanB gene (30%), presenting 12 of them the vanB2 genotype and 1 E. faecalis the vanB1 genotype. All of them showed a vancomycin MIC ≥32 mg/L and a teicoplanin MIC ≤1 mg/L. The vanB2 genotype was detected in seven E. faecium and five E. faecalis isolates (Table 1).
On the other hand, the vanA genotype was found in seven isolates (16% of VRE): two E. faecalis and five E. faecium (Table 1); all of them showed vancomycin and teicoplanin resistance (MIC ≥32 mg/L). Intrinsic vancomycin resistance was also detected in 23 isolates (22 E. gallinarum/vanC1 and 1 E. casseliflavus/vanC2) and they showed a vancomycin MIC ≥16 mg/L and teicoplanin MIC ≤1 mg/L (Table 1).
Almost all VRE analyzed in this study presented a phenotype of multiresistance (Table 1) and harbored different associated resistance genes (number of isolates): erm(B) (30), tet(M) (30), tet(L) (20), ant(6)-Ia (23), aac(6′)-Ie-aph(2′′)-Ia (16), aph(3′)-IIIa (30), and catA (2). Regarding the different enterococcal species, differences in resistance phenotypes were detected; all E. faecium isolates showed resistance to ampicillin, but none of the E. faecalis isolates were ampicillin resistant. Tetracycline resistance was more frequent among E. faecalis (62%) than among E. faecium isolates (16%), and fluoroquinolone resistance was frequent in E. faecalis and E. faecium (88% and 100%, respectively), and almost absent in E. gallinarum/E. casseliflavus isolates (2%). In the case of high-level resistance to aminoglycosides, E. faecalis was the predominant resistant species. The combination of the different genes as well as the correlation with the resistance phenotype is shown in Table 1.
Gelatinase activity and virulence genes
Fourteen VRE isolates of the 43 tested showed a positive gelatinase activity (33%); most of them carried the vanA or vanB genes (11 strains), and only three contained the vanC1 gene. Regarding species, E. faecalis was the most common species showing the gelatinase activity (67%) versus 36% and 13% for E. faecium and E. gallinarum/E. casseliflavus species, respectively. All but two of the strains with the gelatinase activity harbored the gelE gene, associated with the production of this enzyme.
The presence of other virulence genes was also studied. Esp and hyl were the most frequently detected virulence traits (48% and 71%, respectively), although they were detected only in E. faecium (58% and 25%) and E. faecalis species (78% and 89%). These genes were detected in different associations [esp/hyl (no. strains)]: +/+ (9); +/− (5); −/+ (2); and −/− (3) (Table 1). Ace and agg genes were found in a lower percentage and mostly among E. faecalis strains (43% and 24%, respectively).
Vancomycin resistance mechanism
Characterization of the Tn1546 structure of all seven vanA-containing enterococci was carried out to detect different polymorphisms. Four of these strains presented the Tn1546-type I, identical to the prototype (GenBank accession no. M97297), which contains the complete structure, with no insertions or deletions. The remaining three strains presented the structures: one E. faecium strain had the ISEf1 in the vanX-vanY intergenic region; two E. faecium strains harbored the IS1542 between orf2 and vanR, and one of them also presented a deletion in orf1 (Fig. 1).
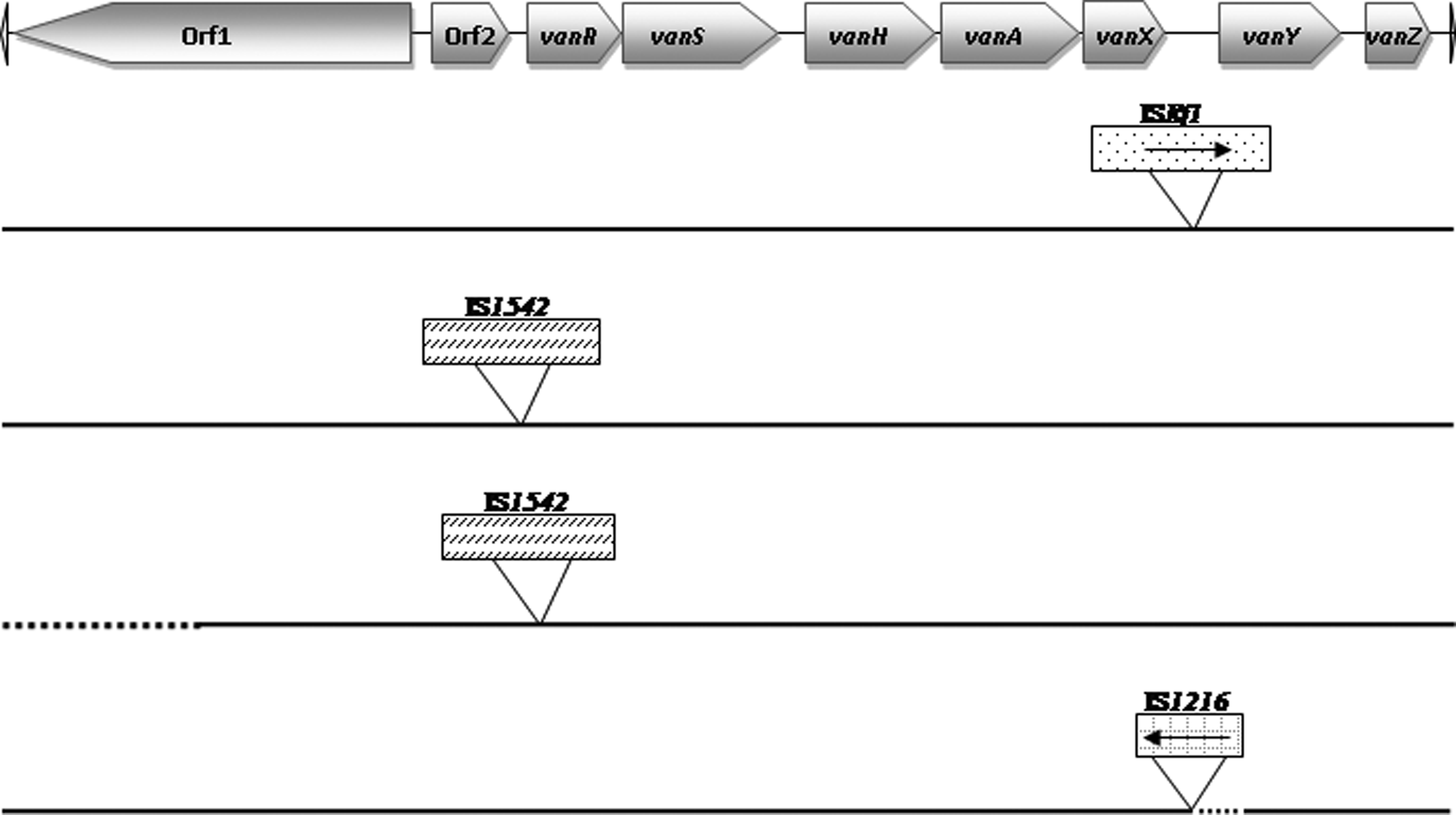
Genetic maps of the different Tn1546 structures found among the 7 vanA strains. Tn1546 type I corresponds to the original sequence previously described 4 without insertions or deletions. Insertion sequence (IS) elements are represented by rectangles and the insertion position is indicated. Deletions are represented by dots.
In relation to isolates containing the vanB2 gene, the inclusion of the vanB2 operon in Tn5382 was analyzed in one strain of each different PFGE pattern. Five out of the eight analyzed strains (C2270, C2278, C2292, C2307, and C2309) showed positive PCR results for Tn5382, and the sequences obtained showed 100% identity with a partial sequence of the previously reported Tn5382 and Tn1549 transposons (Genbank accession numbers AF175739 and AF192329, respectively). Positive amplicons for the vanXB/ORFC region were obtained for these five isolates, and their sequences corresponded (100% of homology) with the AF203412 sequence included in the Genbank. 12 According to these results, the vanB2 gene cluster is included in a Tn5382-like element in these seven strains, but not in the other three. None of them harbored the insertion sequences ISEnfa110, ISEnfa200, or IS150.
In two of the strains with vanB2 included in Tn5382 (C2292 and C2307), it was possible to detect the presence of pbp5-Tn5382 amplicons of the expected size, confirming that sequences of Tn5382 were located immediately downstream from a pbp5 gene as previously reported. 12 These two strains were AMP-resistant E. faecium strains. The analysis of the pbp5 gene was performed in one of these strains (number C2292), and nine amino acid changes were found (Q461K, H470Q, M485A, N496K, A499T, E525D, V586L, E629V, and P667S).
Molecular characterization by MLST
All 12 vancomycin-resistant E. faecium strains were characterized by MLST (one isolate per patient and per PFGE pattern). Six different STs were identified, one of them not previously described, with a new allele combination (9, 1, 1, 1, 5, 1, and 1), and registered in the database as ST612; the other five were ascribed to the high-risk clonal complex CC17 (ST17, ST18, ST132, ST203, and ST267). Interestingly, all E. faecium-vanB2 belonged to ST18, except one strain presenting the ST267, a single locus variant of ST18. However, this strain showed an indistinguishable PFGE pattern with other E. faecium-vanB2 strain (Table 1).
Five different STs were identified among the eight vancomycin-resistant E. faecalis strains (one isolate per patient and per PFGE pattern) and they were included in various ST and CCs: ST6 (CC2), ST9 and ST17 (CC9), ST16 (CC58), and the singleton ST49 (Table 1).
Discussion
The low prevalence of VRE among enterococcal isolates recovered over the 8-year period in our hospital is similar to data presented in another study previously performed in Spain. 30 The most frequently isolated VRE species was E. gallinarum, which is known to present intrinsic resistance to glycopeptides. 8 Concerning isolates with acquired resistance to vancomycin, E. faecium was more frequently recovered than E. faecalis (28% vs. 19%), although E. faecalis was the most common species isolated among the total enterococci (79% vs. 19%). This result is in agreement with other studies, which reported that E. faecium was the most prevalent VRE-isolated species. 7
The most frequently acquired vancomycin resistance mechanism in our study was vanB (30%), being vanA detected with lower frequency (16%). However, vanA seems to be the most widespread genotype in hospital settings all over Europe,5,7,15 as well as in other institutions worldwide. 2 It is of relevance the detection in this study of one E. faecalis isolate with the vanB1 genotype, mechanism first described in 1993, 17 and afterward, very infrequently detected.
The study of the clonal relationship among the isolates demonstrated a wide diversity among them. All the E. faecalis isolates showed unrelated-PFGE patterns and eight unrelated patterns were identified among the 12 vancomycin-resistant E. faecium isolates, of which only five vanB2-E. faecium isolates exhibited indistinguishable or closely related patterns. Lower diversity of clones have been detected in other previous studies.2,5
The genetic element carrying the vanA or vanB cluster of genes was characterized. The Tn1546 type I, identical to the prototype, was the most frequent structure among our vanA strains (57%). Three different variants were detected. All of them have been previously described, also in clinical isolates, 23 and resistance genes were not interrupted, although one of them presented a deletion in the orf1, which codifies a transposase.
The vanB2 mechanism was also characterized, and the vanB2 cluster of genes was detected in five out of the eight strains included into Tn5382, as previously found by other authors.12,36 The pbp5 gene was located immediately upstream of Tn5382, supporting their genetic linkage described by other authors. 12 The vanB2 gene could be included into other transposons in the remaining three strains, as is the case of Tn1547. 31
The MLST genetic characterization revealed that, as expected, all E. faecium strains were ascribed to the CC17, and belonged to different STs (ST17, ST18, ST132, ST203, and ST267). A strong association was found between this HRCC and the hospital environment.35,39 Furthermore, this CC is highly associated with ampicillin and fluoroquinolone resistance as well as with the presence of esp virulence gene, as it is shown in our strains. The five different STs found among E. faecalis strains had been previously detected in isolates from the clinical setting, 32 and three of them were included into the HRCC CC2 and CC9, related with the hospital environment. The sequence type ST49 found in one E. faecalis strain had been previously detected in glycopeptide-resistant clinical isolates. 33 Both HRCCs detected in E. faecalis and E. faecium strains in this study had been previously reported both in vancomycin-resistant and vancomycin-susceptible strains, suggesting that these special lineages represent a good substrate for the later acquisition of resistance.33,40
The detection of the gelatinase activity in 33% of the VRE strains is of relevance because this activity has been related to pathogenesis through its implication in biofilm formation. 26 It is interesting to note that esp and hyl genes were only detected among E. faecalis and E. faecium isolates (specially in E. faecalis), and not among the other enterococcal species. Other studies found similar prevalence of these genes in vancomycin-resistant E. faecalis and E. faecium species, 5 although others identified the esp gene more frequently than the putative virulence hyl gene.10,25
In summary, although a low proportion of the enterococci recovered in the studied hospital showed an acquired mechanism of vancomycin resistance, it is of interest the diversity of the detected genotypes (vanA, vanB2, and vanB1) and the high clonal diversity of these VRE strains, most of them included in HRCCs, as well as the diversity of the Tn1546-vanA detected structures. Most of these VRE strains frequently carried antimicrobial resistance and virulence genes. These results show the relevance of monitoring the evolution of VRE along time in this and in other hospitals to detect future possible changes in prevalence and genotypes of this important multidrug-resistant pathogen, which holds special significance in human health.
Footnotes
Acknowledgments
M. López has a fellowship of the Gobierno de La Rioja of Spain. This study was financially supported by Project SAF2009-08570 from the Ministerio de Ciencia e Innovación of Spain.
Disclosure Statement
No competing financial interests exist.