
Abstract
Urogenital Neisseria gonorrhoeae isolates (266) collected in San Francisco, CA, in 2009 were analyzed for antimicrobial susceptibility and were subsequently genotyped by N. gonorrhoeae multiantigen sequence typing (NG-MAST). Isolates of identical or closely related sequence types were found to possess highly similar phenotypes with regard to drug susceptibility. Isolates containing decreased susceptibility to oral cephalosporins were detected in 2009 and were found to contain the mosaic penA allele (XXXIV) found previously to be associated with decreased susceptibility to cephalosporins. A better understanding of the relationships between phenotypic and genotypic markers for antimicrobial resistance may be helpful to the development of effective surveillance systems for drug-resistant N. gonorrhoeae.
Introduction
We sought to characterize the relatedness of all GISP isolates collected at the San Francisco, CA, municipal STD clinic during 2009. Isolates were genotyped by the N. gonorrhoeae genotyping method, N. gonorrhoeae multiantigen sequence typing (NG-MAST). 12 Isolates considered identical or closely related according to NG-MAST were compared to one another with regard to their antimicrobial susceptibility phenotypes. Such an analysis allowed a determination of whether phenotypic susceptibility patterns of isolates segregated according to the NG-MAST sequence type. Moreover, during the course of this analysis, isolates from 2009 with reduced susceptibility to oral cephalosporins were detected and their genotypes subsequently analyzed.
Materials and Methods
Bacterial collection
Two hundred sixty-six isolates of urogenital N. gonorrhoeae were collected from symptomatic men at San Francisco City Clinic, the municipal STD clinic, during 2009. All cultures were initially isolated on Modified Thayer Martin Selective Agar (Becton Dickinson) and immediately incubated for 24 hours in 10% CO2 at 35°C. Plates were transferred to the San Francisco Public Health Laboratory and identified via Gram stain and oxidase tests. Cultures were purified on chocolate agar and incubated for 24 hours in 10% CO2 at 35°C. Pure cultures were frozen as paired samples in a Tryptic Soy Broth with 1% Glycerol. One set of samples was shipped to a CDC-sponsored GISP laboratory at the University of Washington, Seattle. The second set was stored at −30°C.
Gonococcal Isolate Surveillance Project
Isolates sent to the University of Washington GISP Laboratory were subjected to antibiotic susceptibility testing using the agar dilution method as outlined by the CDC GISP protocol. 9 For standardization and to facilitate execution, this protocol includes the collection of urogenital specimens from men. Isolates were examined for minimum inhibitory concentration (MIC) against the following eight drugs: penicillin, tetracycline, spectinomycin, cefixime, ceftriaxone, ciprofloxacin, cefpodoxime, and azithromycin (only MIC for ciprofloxacin, cefpodoxime, and azithromycin are shown). The CDC assigns the following alert MIC values to the following compounds: cefixime, ≥0.250 μg/ml; cefpodoxime, ≥0.250 μg/ml; azithromycin, ≥2.0 μg/ml; ceftriaxone: 0.125 μg/ml.
N. gonorrhoeae multiantigen sequence typing
N. gonorrhoeae DNA for genetic analysis was prepared by automated DNA extraction using the Total NA protocol on the MagNA pure instrument (Roche Diagnostics). NG-MAST sequence typing was performed by amplifying regions of the porB and tbpB genes using a published polymerase chain reaction (PCR) protocol. 12 porB and tbpB products were visualized on a 1% agarose gel; porB products were visible around 700 bp and tbpB products at the around 500 bp after electrophoresis for 60 minutes. The PCR products were purified using an automated PCR-Cleanup protocol (Qiagen). All strains that occurred with a frequency greater than five isolates detected in 2009 were subject to analysis and are shown in this work. This correlates to a prevalence of 2%. Primers were obtained from Integrated DNA Technologies, San Diego, CA.
Maximum likelihood tree generation
porB alleles and tbpB alleles were combined into a single sequence for all NG-MAST sequence types in this study, por and tbpB sequences were combined into a single sequence and were assembled into standard FASTA files. These sequences were aligned using Bioedit standard CLUSTAL-W alignment. 8 The aligned sequences were assembled into a relatedness tree using DNAml DNA maximum likelihood method. 7
DNA sequencing
Purified samples were subjected to DNA sequencing for penA, tbpB, or porB. Sequencing reactions for the por gene were carried out by using Bigdye 3.1 on the ABI Genetic Analyzer and separated by matrix (NoanoPOP 7) on 3730xl (Applied Biosystems Inc.). Each sample was subjected to 30 cycles of 98°C for 10 seconds, 50°C for 5 seconds, and 60°C for 2.30 minutes; reactions for the tbpB gene were run with the same conditions, but with an annealing temperature of 58°C. The resulting DNA sequences were entered into the NG-MAST online database (www.ng-mast.net) and assigned allele/sequence type identities. Primers were obtained from Integrated DNA Technologies, San Diego, CA.
Mosaic penA identification
The mosaic penA allele was screened using a real-time PCR assay previously described by Ochiai et al. 13 penA sequencing was performed as previously described. 16
Results
In total, 266 N. gonorrhoeae isolates were collected evenly from January through December 2009 and were analyzed with regard to antimicrobial susceptibility by the agar dilution method within 3 months of collection. These 266 isolates constituted 40% of all positive gonorrhea cases in men over the course of 2009 at the clinical collection site. Antimicrobial agents tested included penicillin, tetracycline, spectinomycin, ceftriaxone, cefixime, cefpodoxime, ciprofloxacin, and azithromycin. All isolates were subsequently subjected to genotyping by way of the NG-MAST method. This included amplification and sequencing of the porB and tbpB alleles before submission of the allelic sequences to the NG-MAST database for sequence-type assignment.
Application of NG-MAST to all 266 isolates revealed 117 different sequence types. The majority of the isolates (150/266, 56.4%) belonged to 15 sequence types, whereas the remaining isolates (116, 43.6%) belonged to 102 distinct sequence types. One sequence type, ST2992, accounted for 20% of all isolates. Of the 117 different sequence types detected, 82 were novel to the NG-MAST database and as such were first-time contributions to the database.
To determine the relatedness of the eight most common sequence types in our dataset, we generated a relatedness tree (Fig. 1) using the DNA maximum likelihood methodology on the porB and tbpB sequences for each sequence type. Two features were apparent in this relatedness tree. First was that two porB and tbpB alleles were common among our set of isolates. Both ST3935 and ST1407 contain the porB 908 allele, while sequence types 3935, 730, and 2992 all contain tbpB 29. Overall, 119 of 266 isolates (45%) possessed the tbpB 29 allele. These shared allele types suggest possible relatedness among the strains surveyed in our collection. Additionally, it was found that ST4254 and ST589 are identical in their tbpB sequence and nearly identical in their porB sequence (only two nucleotides different). By comparison, ST589 and ST2992 (the next closest sequence type in this study, by relatedness) have 74 nucleotide differences and a number of insertions and deletions when aligned with each other. The near identity between ST4254 and ST589 suggests that they are likely descendant from a common ancestor strain in the very near past. Both sequence types were found also to possess identical sensitivity patterns to the antimicrobials tested.

Unrooted relatedness tree generated using DNA maximum likelihood on the por and tbp sequences for the seven most common strain types in 2009. Each terminal branch is labeled with the Neisseria gonorrhoeae multiantigen sequence-typing (NG-MAST) strain type represented on that branch. The blue-shaded region indicates all strain types with the tbp allele 29. The red-shaded area indicates all strain types with the por allele 908.
These results led us to investigate whether the other major sequence types in our study were closely related to any of the minority sequence types. We found that each sequence type had at least one additional isolate that was a near identical (seven or fewer nucleotide differences, which constitutes a 1% overall variability in sequence divergence for the sequences of tbpB and porB combined) sequence match for NG-MAST. These sequence types are listed in Table 1. An example of the close relatedness of these additional sequence types is portrayed in Fig. 2. For this figure, we analyzed the relatedness of the eight most common sequence types and the five sequence types closely related to ST2992. Expanding the grouping of sequence types to include these other, very closely related sequence types (using the same criteria of seven or fewer nucleotide differences) creates an ST2992-like group with a membership of 60 isolates (60/266, 22.5%). When we compare the antimicrobial sensitivities for these additional strains that are nearly identical to one of our eight most common sequence types, we find that in all but one case, the sensitivity profiles are identical (data not shown). The single exception is seen for ST28 (nine total isolates) and ST1440 (four total isolates). These sequence types have identical tbpB sequences and only one nucleotide difference in their porB sequence. However, all nine isolates of ST28 show sensitivity to the lowest concentrations of ciprofloxacin (MIC ≤0.030). In contrast, all four isolates of ST1440 possessed alert levels for ciprofloxacin MIC (MIC levels of 0.25, 4.0, 4.0, and 16.0 for these four isolates). While these two sequence types are clearly related to a common ancestor in the recent past, divergence has occurred. Two possibilities for this include either that ST1440 has acquired a decreased susceptibility to ciprofloxacin or that ST28 has lost decreased susceptibility to ciprofloxacin.
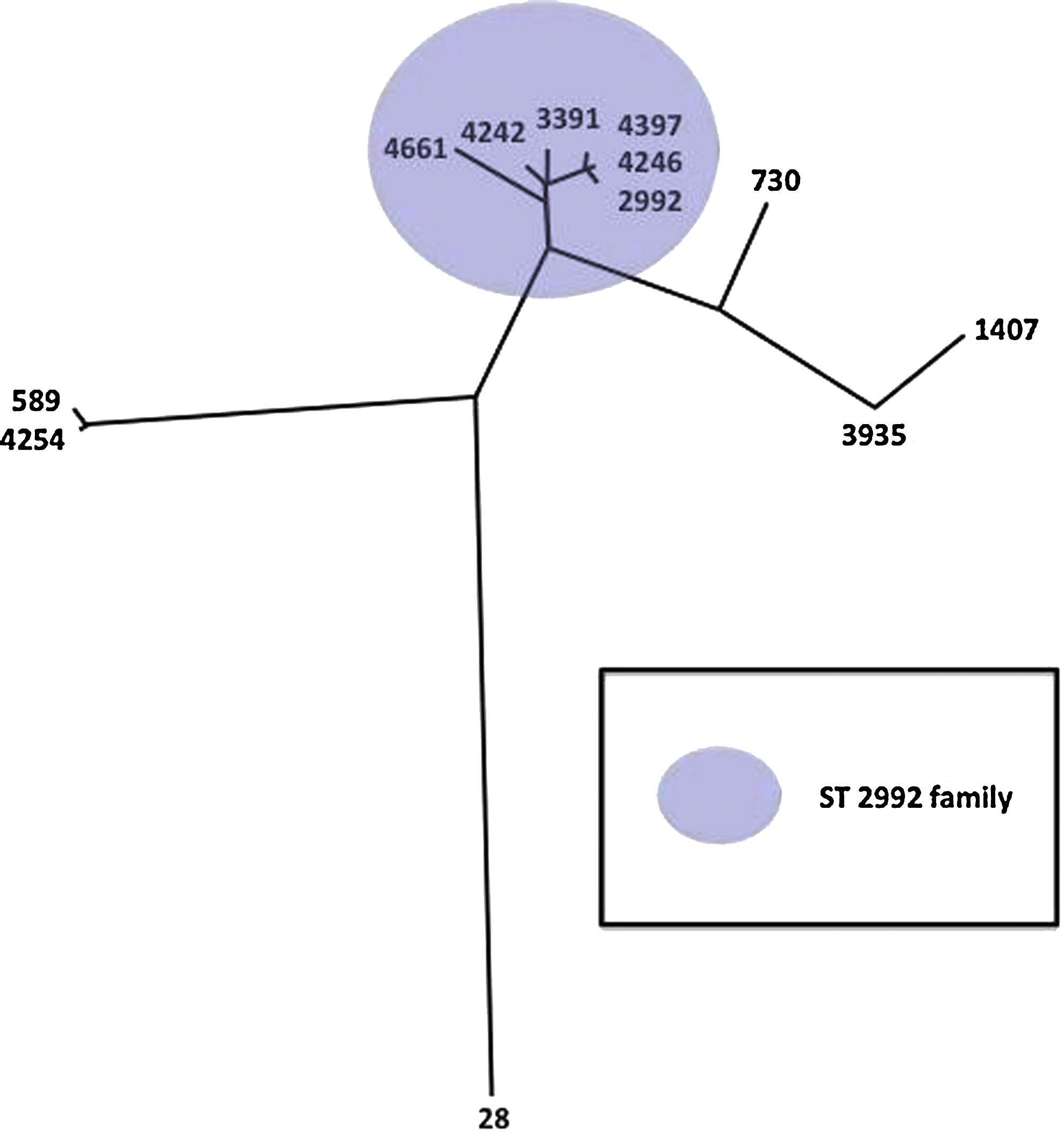
Unrooted relatedness tree generated using DNA maximum likelihood on the por and tbp sequences for the seven most common strain types in 2009 and the six strain types closely related to the strain type 2992. Each terminal branch is labeled with the NG-MAST strain type represented on that branch. The purple-shaded region indicates all strain types in the 2992 strain type family.
The number in parentheses indicates the number of isolates of that strain type in our study. No number indicates that only a single isolate of that strain type was found.
We next determined that whether isolates of the same NG-MAST sequence type possessed comparable phenotypic profiles with regard to drug susceptibility. Inspection of antibiograms for isolates of the same NG-MAST type revealed highly comparable MIC values for most antimicrobial agents. For each sequence type that occurred with a frequency >2% (i.e., sequence types for which more than 5 isolates were available for comparison), we constructed histograms to illustrate the distributions of MIC that occurred for cefpodoxime, ciprofloxacin, and azithromycin. (Fig. 3) Histograms describing ceftriaxone were not constructed because of the complete homogeneity of MIC values to this agent across all isolates (only 2/266 instances of MIC >0.03). Cefixime is likewise not shown since another oral cephalosporin, cefpodoxime, is shown.
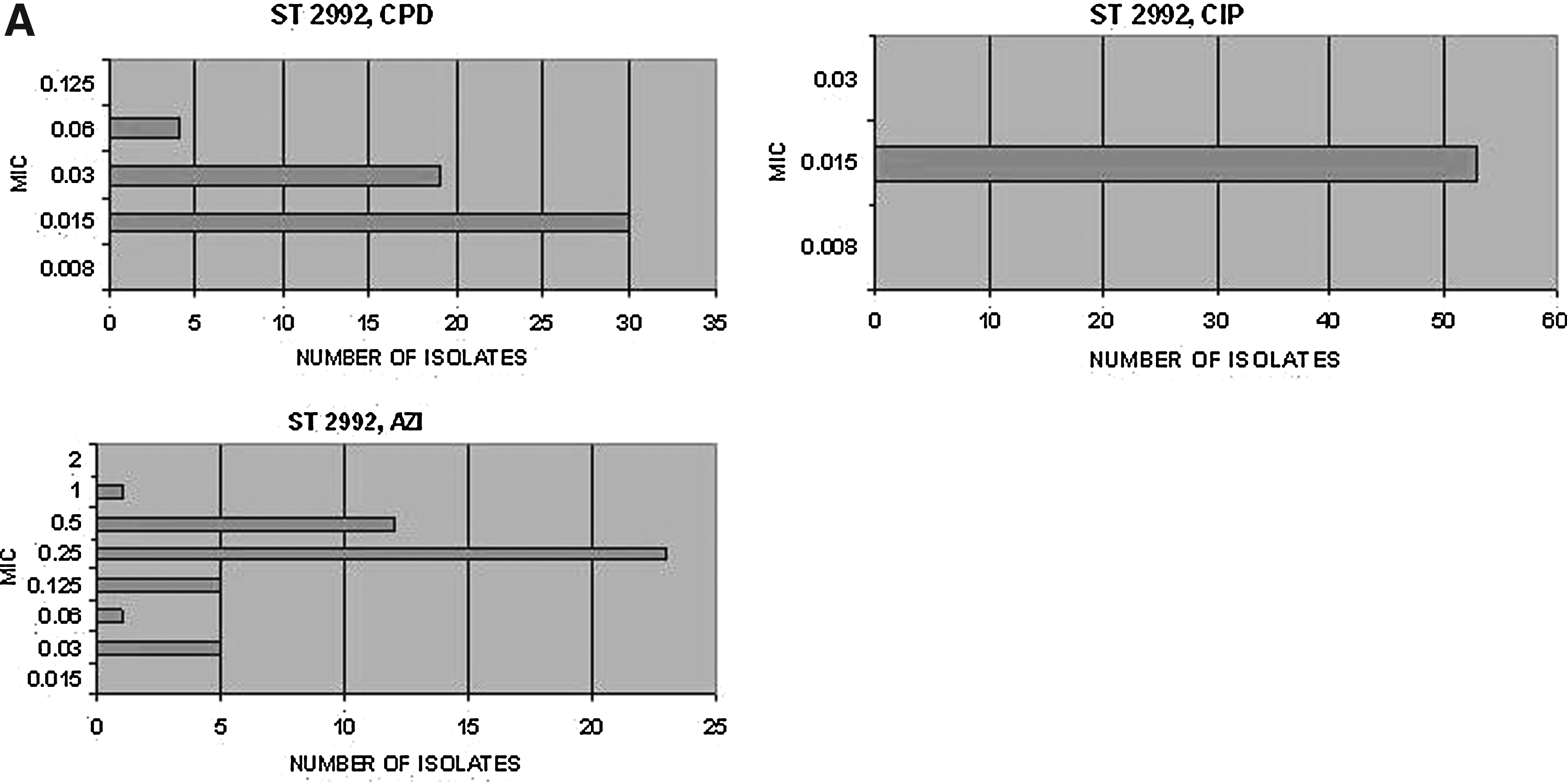
Within sequence types, MIC values for most antimicrobial agents were highly similar. A summary of the variability in drug susceptibility seen for each of the major genotypes is provided in Table 2. Across the evaluated sequence types and weighted for the number of isolates, the NG-MAST sequence type was found to be predictive of MIC within a twofold range from the modal MIC: for cefpodoxime, this occurred 91% of the time, for ciprofloxacin 98% of the time, and for azithromycin 81% of the time. A twofold differential from a given MIC value is considered as an acceptable level of variation for proficiency testing in the performance of the GISP. 9 A notable variation was seen in the cefpodoxime MIC for ST1407. This sequence type has been shown, previously, both to possess a decreased susceptibility for and to be associated with treatment failures to oral cephalosporins.1,14,16
ST730 was multimodal for CPD.
An MIC of 8.0 occurred, n=1. All other isolates (n=8) occurred with MIC of 0.015.
NG-MAST, Neisseria gonorrhoeae multiantigen sequence typing; MIC, minimum inhibitory concentration; CPD, cefpodoxime; CIP, ciprofloxacin; AZI, azithromycin.
Bold values indicate a singular MIC for all isolates tested.
We noted that some of the isolates from 2009 were found to possess alert values (MIC ≥0.250 mg/L) for cefpodoxime (5 of 266 isolates, 1.9%). Previous studies in the same geographic area had revealed the presence of N. gonorrhoeae isolates that possessed a reduced susceptibility to oral cephalosporins. 16 Such isolates had been shown to possess highly altered mosaic penA alleles. For that reason, we analyzed all 266 of the 2009 isolates described herein for the presence of mosaic penA alleles using a real-time PCR screening assay. This PCR assay detects mosaic penA alleles, including those previously associated with reduced susceptibility to oral cephalosporins in vitro, or with actual treatment failures.10,13,16 Screening by this method revealed that 9 of 266 (3.8%) isolates possessed PCR-reactive penA alleles. All nine isolates were subsequently subjected to DNA sequencing of the entire penA gene to confirm the PCR results and to determine specifically what the penA allele possessed. As shown in Table 3, all nine PCR-reactive isolates possessed a mosaic penA allele that was previously described and detected in San Francisco. Four of the isolates possessed the penA allele XXXIV (previously referred to as SF-A 16 ), which contains a high degree of similarity with that of the mosaic allele X previously described and associated with oral cephalosporin treatment failures in Europe and Asia. 14 Five of the isolates contained the penA allele XXXVIII (previously referred to as SF-B 16 ), which possesses a markedly different sequence, and lacks certain of the mutations shown previously to be required for conference of oral cephalosporin resistance.14,17
Mosaic penA allele nomenclature as per Ref. 11
1 base-pair difference with the allele 908.
PEN, penicillin; TET, tetracycline; CFX, cefixime; CRO, ceftriaxone.
All seven of the ST 1407 isolates detected during this study possessed mosaic penA alleles. Those ST1407 isolates that contained the XXXIV penA allele possessed MIC of 0.5 and 2.0 for cefpodoxime. One isolate (ST 4533) possessed a mosaic penA allele (XXXIV) and an MIC of 1.0 for cefpodoxime. Another isolate, ST 3431, also contained the XXXIV allele and possessed an MIC of 1.0 for cefpodoxime. This sequence type had been previously described, as an isolate of this sequence type was associated with a treatment failure to cefixime in England. 10 All isolates that contained the XXXVIII penA allele possessed MIC of 0.125 or below for cefpodoxime. All isolates containing either XXXIV or XXXVIII penA alleles also possessed allele 110 for tbpB.
Discussion
A previous study has shown that isolates of the same NG-MAST sequence type were found to possess highly similar phenotypic patterns of drug susceptibility. 15 This is perhaps not surprising, given that isolates of the same NG-MAST type are either identical or closely related. Larger NG-MAST families in our study unsurprisingly tended to possess a greater variability of MIC. For most NG-MAST types investigated, variability was particularly high among the MIC for azithromycin. A previous study investigating isolates from San Diego County with unusually a high MIC for azithromycin (MIC of 8.0–16.0) showed that those isolates were found to have the NG-MAST type ST2992. 20 In this study, ST2992 isolates were found to have azithromycin MIC ranging from 0.03 to 1.0 (nonalert value). Possibly, the loci that control the phenotype of azithromycin susceptibility are polymorphic and change on a time scale that may be much faster than that of either the por or tbpB gene, which dictates the NG-MAST-typing method. This indicates that while NG-MAST typing may provide valuable information about a particular isolate's antimicrobial susceptibilities, it would by no means be definitive. This caution is further supported by an example such as that seen for ST730, for which one of nine isolates possessed an MIC for ciprofloxacin 533 times higher than that of the other eight ST730 isolates.
This study also revealed the presence of isolates with reduced susceptibility to oral cephalosporins in San Francisco in 2009. Seven of 266 isolates (2.6%) possessed the ST1407 sequence type, which has previously been associated with treatment failures to oral cephalosporins. All ST1407 isolates were found to possess mosaic penA alleles, previously described in a study of local Gonococcal isolates (XXXIV and XXXVIII). Previously, it was shown that the XXXIV allele possesses the I312M, V316T, and G545S mutations in penA, which are each vital in contributing to the phenotype of reduced susceptibility. 17 The XXXVIII allele was shown to possess I312M and V316T with the wild-type residue (glycine) at position 545. 16
Isolates of sequence types ST1407 and ST3431 that were found to possess the XXXIV mosaic allele also possessed markedly elevated MIC to oral cephalosporin, whereas those possessing the XXXVIII mosaic allele did not. Of note was the fact that all mosaic penA-containing isolates also possessed the 110 tbpB allele. The explanation for this may be rooted in the fact that penA and tbpB are very close to one another on the N. gonorrhoeae chromosome. 6 The exchange of genetic material responsible for acquisition of a penA allele may also involve incorporation of a portion of the 110 tbpB locus.
The relatively low frequency (10.2%) of isolates with alert value MIC for ciprofloxacin found in 2009 in this study was of interest. In previous years, in San Francisco, particularly 2005–2007, frequencies of isolates with alert value ciprofloxacin MIC ranged from 30% to 45% annually. It is unknown whether the drop in frequency seen for alert value MIC for ciprofloxacin is due to a shift in sequence types located in San Francisco, or whether sequence types have persisted over time, but have lost the mutations required for resistance due to a lack of selective pressure. Experiments are in progress to answer this question.
The data herein support that phenotypic markers of antimicrobial resistance are associated with specific N. gonorrhoeae sequence types. These methods hold promise in improving our ability to better monitor potential increases in local transmission and the importation of resistant strains of N. gonorrhoeae. Further characterization of the phenotypic, genotypic, and epidemiologic characteristics of NG from diverse geographic areas may be needed.
Footnotes
Acknowledgments
We thank the San Francisco Department of Public Health STD Control branch and City Clinic for the acquisition of GISP isolates. Antimicrobial phenotype testing was provided by the CDC through the University of Washington Regional GISP Laboratory in Seattle, WA. This research was supported in part by the Emerging Infectious Diseases (EID) Fellowship Program administered by the Association of Public Health Laboratories (APHL) and funded by the CDC.
Disclosure Statement
No competing financial interests exist.