
Abstract
Lantibiotics are a unique group within the antimicrobial peptides characterized by the presence of thioether amino acids (lanthionine and methyllanthionine). These peptides are produced by and primarily act on Gram-positive bacteria exerting multiple activities at the cytoplasmic membrane of susceptible strains. Previously, the cell wall precursor lipid II was identified as the molecular target for the prototype lantibiotic nisin. Binding and sequestration of lipid II blocks the incorporation of the central cell wall precursor into the growing peptidoglycan network, thereby inhibiting the formation of a functional cell wall. Additionally, nisin combines this activity with a unique target-mediated pore formation, using lipid II as a docking molecule. The interaction with the pyrophosphate moiety of lipid II is crucial for nisin binding. We show that, besides binding to lipid II, nisin interacts with the lipid intermediates lipid III (undecaprenol-pyrophosphate-N-acetyl-glucosamine) and lipid IV (undecaprenol-pyrophosphate-N-acetyl-glucosamine-N-acetyl-mannosamine) of the wall teichoic acid (WTA) biosynthesis pathway. Binding of nisin to the precursors was observed at a stoichiometry of 2:1. The specific interaction with WTA precursors further promoted target-mediated pore formation in artificial lipid bilayers. Specific interactions with lipid III and lipid IV could also be demonstrated for related type A lantibiotics, for example, gallidermin, containing the conserved lipid-II-binding motif.
Introduction
The first discovered and most extensively studied lantibiotic is nisin. 24 It is produced by some Lactococcus lactis strains and widely used as a food preservative. Due to its low stability at physiological pH, nisin has never been developed for systemic application. However, the increasing problems with antibiotic resistance and the urge to find potent antibiotics have renewed the interest in potential clinical applications of lantibiotics. This seems even more promising, as the understanding of the organization of the lantibiotic gene clusters and the biosynthetic pathways has made protein engineering feasible. Several derivatives of nisin and other lantibiotics with improved stability or antimicrobial activity have been synthezised.23,25
Early experiments demonstrated that, due to their strong amphiphilic nature, nisin and related lantibiotics impair membranes causing dissipation of the membrane potential, rapid efflux of small metabolites, and cessation of cellular biosynthetic processes.27,29,30 Subsequently, nisin was shown to destabilize model membranes at concentrations in the micromolar range15,19 and to bind and sequester the peptidoglycan precursor lipid II, which enables the formation of specific pores in the nanomolar range.8,10,38
Lipid II represents the central cell wall building block of peptidoglycan biosynthesis that is structurally conserved among eubacteria. The precursor consists of the bactoprenol carrier lipid (C55-P), which is linked to the disaccharide unit N-acetyl-muramyl-pentapeptide-N-acetyl-glucosamine (MurNAc-pp-GlcNAc) via a pyrophosphate bridge. Lipid II synthesis 6 occurs at the inner face of the cytoplasmic membrane, where the translocase MraY and the transferase MurG link the soluble, UDP-activated sugars UDP-N-acetyl-muramic acid-pentapeptide and UDP-N-acetyl-glucosamine yielding lipid I and lipid II, respectively. Lipid II is then translocated across the cytoplasmic membrane and incorporated into the growing peptidoglycan network through the activity of the penicillin-binding proteins, releasing C55-PP, which after dephosphorylation enters a new synthesis cycle. Binding of nisin to lipid II locks the cell wall precursor in a stable complex, thereby blocking the entire peptidoglycan synthesis cycle.
This dual mode of action, in which inhibition of peptidoglycan synthesis and pore formation are most efficiently combined, makes nisin a very potent antimicrobial agent and impedes the emergence of resistant strains. The binding motif of nisin is conserved among a number of lantibiotics targeting lipid II, for example, subtilin, epidermin, gallidermin, plantaricin C, and mutacin 1140. 40 The interaction primarily involves the formation of five hydrogen bonds between the pyrophosphate moiety of lipid II and the amide backbone of the N-terminal ring pair of nisin, hence referred to as pyrophosphate cage. 20
Besides lipid II, nisin and related lantibiotics were also shown to interact with lipid I in vitro 10 ; additionally, complex formation with bactoprenolpyrophosphate (C55-PP) has been described for nisin. 4 However, this interaction does not seem to promote the formation of pores in vitro. These findings suggest that nisin and related lantibiotics containing the lipid-II-binding motif may interact with further cell envelope precursors, attached to undecaprenylphosphate (C55-P) as a carrier molecule, such as those for capsule or wall teichoic acid (WTA) biosynthesis.
In Staphylococcus aureus, WTA biosynthesis involves at least nine enzymatic steps.11,41 The first step is catalyzed by the membrane-bound enzyme TagO that transfers the N-acetyl-glucosamine-1 phosphate moiety of UDP-N-acetyl-glucosamine to the lipid carrier C55-P, generating lipid III. The epimerase MnaA catalyzes the conversion of UDP-GlcNAc to UDP-N-acetyl-mannosamine, which is subsequently attached to lipid III by TagA, yielding lipid IV. Two units of glycerol-phosphate are further added by TagB and TarF, leading to the formation of lipid V and lipid VI. The polymerase TarL sequentially adds up to 40 ribitol-phosphate-repeating units before the completed precursor is translocated across the cytoplasmic membrane by the activity of the ABC transporter TagGH and attached to the peptidoglycan by a so far unknown mechanism. The carrier lipid is released and enters a new synthesis cycle.
In this study we report the in vitro reconstitution of the TagO-catalyzed reaction, which allows for the first time the fully enzymatic synthesis of WTA intermediates, using the natural bactoprenol lipid carrier. Such full-length lipid chain WTA precursors were examined for complex formation with type A lantibiotics and their ability to serve as docking molecules for nisin-induced pore formation.
Materials and Methods
Chemicals and reagents
All chemicals were of analytical grade or better. 1,2-Dioleoyl-sn-glycero-3-phosphocholine (DOPC) was purchased from Avanti Polar Lipids, Inc. Isoprenoids were obtained from Larodan. Radioactive substances were purchased from Hartmann Analytic. Carboxyfluorescein (CF) was purchased from Sigma Aldrich Chemie GmbH. Gallidermin was obtained from Genmedics.
Bacterial strains and growth conditions
Staphylococcus simulans 22 and Micrococcus luteus DSM 1790 were maintained on tryptic soy agar and grown at 37°C and 30°C, respectively. Escherichia coli strains were grown in Luria-Bertani broth at 37°C unless otherwise stated.
Cloning of WTA biosynthesis enzymes
Standard procedures for molecular cloning and transformation were used.14,31 Genes encoding enzymes involved in WTA biosynthesis were amplified from S. aureus N315 genomic DNA with specific oligonucleotides listed in Table 1. Polymerase chain reaction products, pET21b and a modified pET20b vector (pVLL002, kindly provided by B. Berger-Bächi, Zürich, Switzerland), were digested for ligation by use of appropriate restriction enzymes (Table 1). Cloning was performed in E. coli DH5α. Ampicillin (50 μg/ml) was used as selection marker.
Restriction sites are shown by underline.
Expression and purification of cytoplasmic proteins
E. coli strain BL21 harboring recombinant plasmid pET21b-mnaA or pET21b-tagA was grown at 37°C in LB medium supplemented with 50 μg/ml ampicillin. Proteins were expressed in mid-log phase cultures (OD600 ∼ 0.6) after induction with 0.5 mM IPTG for 3–4 hours at 30°C or 37°C. Cells were harvested by centrifugation, washed in 50 mM Tris/Hcl, pH 7.5, 300 mM NaCl, and resuspended in the same buffer (MnaA-His6) or the same buffer supplemented with 1% Triton X-100 (TarA-His6). 11 After treatment with lysozyme (200 μg/ml), DNAse (100 μg/ml), and RNAse (10 μg/ml), and subsequent sonification and centrifugation, the cleared lysate was subjected to Ni2+-affinity chromatography. After washing with lysis buffer, weakly bound material was removed with 50 mM Tris-HCl (pH 7.5), 300 mM NaCl, and 20 mM imidazole. His-tagged proteins were eluted with buffer containing 50 mM Tris-HCl (pH 7.5), 300 mM NaCl, and 200 mM imidazole. Elution fractions were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Protein-containing fractions were stored at −20°C in 50% glycerol.
Expression and purification of the membrane-bound protein TagO
Purification of recombinant TagO-His6 (Llm) enzyme was performed as described previously with minor modifications. 26 In short, E. coli strain C43 harboring the recombinant plasmid pVLL002-tagO was grown at 30°C in 2YT medium supplemented with 50 μg/ml ampicillin until an OD600 of ∼0.7. Expression of TarO-His6 was induced by addition of IPTG in a final concentration of 1 mM and incubation was continued for 16 hours at 25°C. Cells were harvested by centrifugation and washed in 25 mM Tris-HCl (pH 7.5). Cells were resuspended in buffer A (25 mM Tris-HCl [pH 7.5], 2 mM β-mercaptoethanol, 150 mM NaCl, 30% glycerol, and 1 mM MgCl2) and sonicated, and the resulting suspension was centrifuged (20,000 g, 30 minutes, 4°C). Pelleted bacterial membranes were washed three times to remove remaining cytoplasmic content. Membranes and associated proteins were solubilized in two successive steps with buffer A containing 17.8 and 21.5 mM m-Dodecyl-β-D-maltoside. Solubilized membrane proteins were separated from cell debris by centrifugation (20,000 g, 30 minutes, 4°C). The supernatant containing recombinant TagO-His6 was mixed with Ni2+-NTA-agarose and purification was performed as described previously. 26
Nisin purification
Nisin was obtained and purified from culture supernatants of L. lactis NIZO22186 by chloroform extraction as described previously.3,22
Antibiotic susceptibility testing
Minimal inhibitory concentrations (MICs) were determined by standard broth microdilution, according to the Clinical and Laboratory Standards Institute guidelines, in polypropylene microtiter plates (Nunc brand) using cation-adjusted Mueller-Hinton broth (Oxoid).
Antagonization assays
Antagonization of the antibiotic activity of nisin and gallidermin by potential target molecules was performed by an MIC-based setup in microtiter plates. Nisin (0.2 μg/ml) and gallidermin (0.04 μg/ml) (each corresponding to 8×the MIC) were mixed with potential HPLC-purified antagonists (C55-P, C55-PP, lipid I, lipid II, lipid III, and lipid IV) in a 10-fold molar excess with respect to the antibiotic. M. luteus DSM 1790 (5×105 cfu/ml) was added and samples were examined for visible bacterial growth after overnight incubation.
In vitro WTA synthesis reactions using purified proteins and substrates
The enzymatic activity of the TagO-catalyzed lipid III (undecaprenol-pyrophosphate-N-acetyl-glucosamine) synthesis was determined using purified recombinant TagO protein incubated in the presence of 5 nmol C55-P, 67.5 nmol of UDP-GlcNAc, and 0.75 nmol [ 14 C]-UDP-GlcNAc in 83 mM Tris-HCl (pH 8.0), 6.7 mM MgCl2, 8.3% (v/v) dimethyl sulfoxide, and 10 mM N-lauroylsarcosine. The reaction was initiated by the addition of 6.15 μg of the enzyme TagO-His6 and incubated for 90 minutes at 30°C. TagA- and MnaA-catalyzed synthesis of lipid IV (undecaprenyl-pyrophosphate-N-acetyl-glucosamine-N-acetyl-mannosamine) was performed using HPLC-purified lipid III. About 1 nmol lipid III was incubated with 15 nmol UDP-GlcNAc and 0.2 nmol [ 14 C]-UDP-GlcNAc in the presence of 0.2% Triton X-100, 100 mM Tris-HCl (pH 7.5), and 250 mM NaCl in a final volume of 50 μl. About 1.1 μg of TagA-His6 and MnaA-His6 was added to start the reaction. Synthesized lipid intermediates were extracted from the reaction mixtures with n-butanol/pyridine acetate (pH 4.2) (2:1; vol/vol), analyzed by thin layer chromatography (TLC) as described earlier, 3 and quantified by phosphoimaging. Purification of WTA lipid intermediates was performed as described for lipid II with slight modifications. 34
Nisin and gallidermin were added in a 2:1 molar ratio with respect to the amount of purified C55-P, lipid III, and lipid IV, respectively, in all in vitro assays.
In vitro lipid I/II synthesis and purification
Large scale synthesis and purification of the peptidoglycan precursors lipid I and II were performed as described previously. 34
Mass spectrometry
Mass spectrometry analysis of cell wall precursors was performed using a matrix assisted laser desorption/ionization-time of flight (MALDI-TOF) mass spectrometer (Bruker Biflex; Bruker Daltonics) working in a linear negative mode. Samples were mixed with 6-aza-2-thiothymine (in 50% [v/v] ethanol and 20 mM ammonium citrate) as matrix material and spotted onto the MALDI target. Mass spectra were measured in the range of 1,000–1,200 and 1,300–1,350 Da, respectively, and analyzed by Flexanalysis 2.0 (Bruker Daltonics).
Complex formation of nisin with cell wall precursors
Binding of nisin to C55-P, C55-PP, lipid II, and lipid III was analyzed by incubating 5 nmol of each purified cell wall precursor with 0, 10, or 20 nmol nisin in 50 mM Tris-HCl (pH 7.5). Complex formation was analyzed by incubating the lipid precursors with the respective amount of nisin. Free precursors were extracted with n-butanol/pyridine acetate (pH 4.2) (2:1; vol/vol) and analyzed by TLC. Lipid-containing precursors were detected by phosphomolybdic acid staining.
Quartz crystal microbalance
Quartz crystals were prepared and cleaned as described previously. 12 Supported bilayers were completed by transferring a monolayer consisting of DOPC±0.1 mol% lipid intermediate onto a covalently fixed C16H33SH monolayer using the Langmuir-Blodgett technique. Fitting the quartz crystals into the flow chamber was performed under water to guarantee the integrity of the bilayer. Nisin was investigated at 300 μM. A LiquiLab21 quartz crystal microbalance (ifake.V.) was employed, which enables the detection of frequency in real time. Measurements and calculation of the kinetic binding constants were performed as described.13,36
CF efflux experiments
Large unilamellar vesicles for CF efflux experiments were prepared by the extrusion technique as described previously. 39 Vesicles were made of DOPC doped with 0.1 mol% of the respective cell wall precursors (C55-P, C55-PP, lipid I, lipid II, lipid III, or lipid IV; referring to the total amount of phospholipid). CF-loaded vesicles were prepared with 50 mM CF and then diluted in 2 ml of TBS buffer (10 mM Tris-HCl [pH 7.2]; 0.85% NaCl [w/v]) at a final concentration of 25 μM phospholipid on a phosphorous base. Nisin was added in a final concentration of 50 nM, with or without pre-incubation of liposomes with 50 nM of gallidermin for 60 seconds, respectively. The increase in fluorescence intensity was measured at 520 nm (excitation at 492 nm) on an RF-5301 spectrophotometer (Shimadzu) at room temperature. The peptide-induced CF leakage was expressed relative to the total amount of CF released after lysis of the vesicles by addition of 20 μl of 20% Triton X-100.
Results
In vitro synthesis of WTA precursors lipid III and lipid IV
Undecaprenylphosphate represents the central-membrane-bound lipid carrier involved in the synthesis of diverse bacterial cell envelope polymers and polysaccharides, including peptidoglycan and WTA biosynthesis.
We used the well-characterized in vitro lipid II biosynthesis system to calibrate a TLC detection method for the WTA lipid intermediates.32,33
Purified, recombinant MraY and MurG synthesize the peptidoglycan precursors lipid I and lipid II in vitro, 5 in the presence of C55-P and the respective soluble UDP-activated sugars UDP-MurNAc-pentapeptide and UDP-GlcNAc. Compared to C55-P, which migrates to a defined position in the TLC system used (Fig. 1, lane 3; Rf=0.95), migration of lipid I (lane 4; Rf=0.63) and lipid II (lane 5; Rf=0.57) is retarded, as a result of increased polarity.
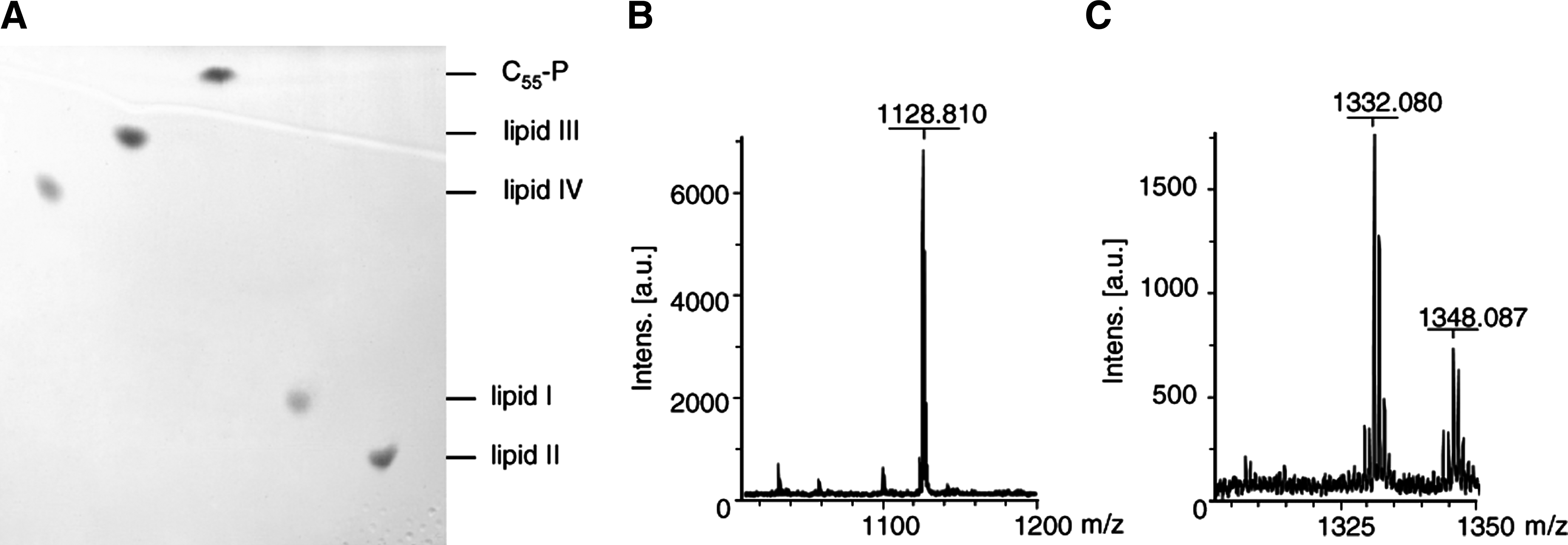
Thin-layer chromatography of purified wall teichoic acid (WTA) (lipid III and lipid IV) and peptidoglycan (lipid I and lipid II) precursors
Altered migration was also observed for the TagO reaction product lipid III (undecaprenyl-PP-GlcNAc) (lane 2). The TagO-catalyzed addition of N-acetylglucosamine to C55-P increased the polarity of the molecule and lowered the mobility in the TLC system (Rf=0.89). The same was true for the second WTA intermediate lipid IV, which was synthesized using the recombinant purified enzymes TagO, TagA, and MnaA. The addition of a second sugar further increased polarity, correlating with the altered migration behavior (Rf=0.84). The identity of lipid III and lipid IV was confirmed by MALDI-TOF mass spectrometry (Fig. 1BC).
Interaction of nisin and gallidermin with WTA precursors
Binding of nisin to lipid II mainly involves the pyrophosphate and the MurNAc moiety as revealed by structural analysis of the nisin:lipid II complex, 20 while the second sugar (GlcNAc) does not appear to significantly contribute to binding.
As lipid III, the first lipid intermediate of the WTA biosynthesis pathway, has a similar pyrophosphate sugar linkage moiety, we investigated the impact of nisin on the lipid III synthesis reaction using purified TagO, C55-P, and [ 14 C]-UDP-GlcNAc. In the positive control, a radiolabeled lipid band was detected, corresponding to lipid III (Fig. 2, lane 2; Rf=0.89), whereas no synthesis product was observed when TagO was omitted (lane 1).
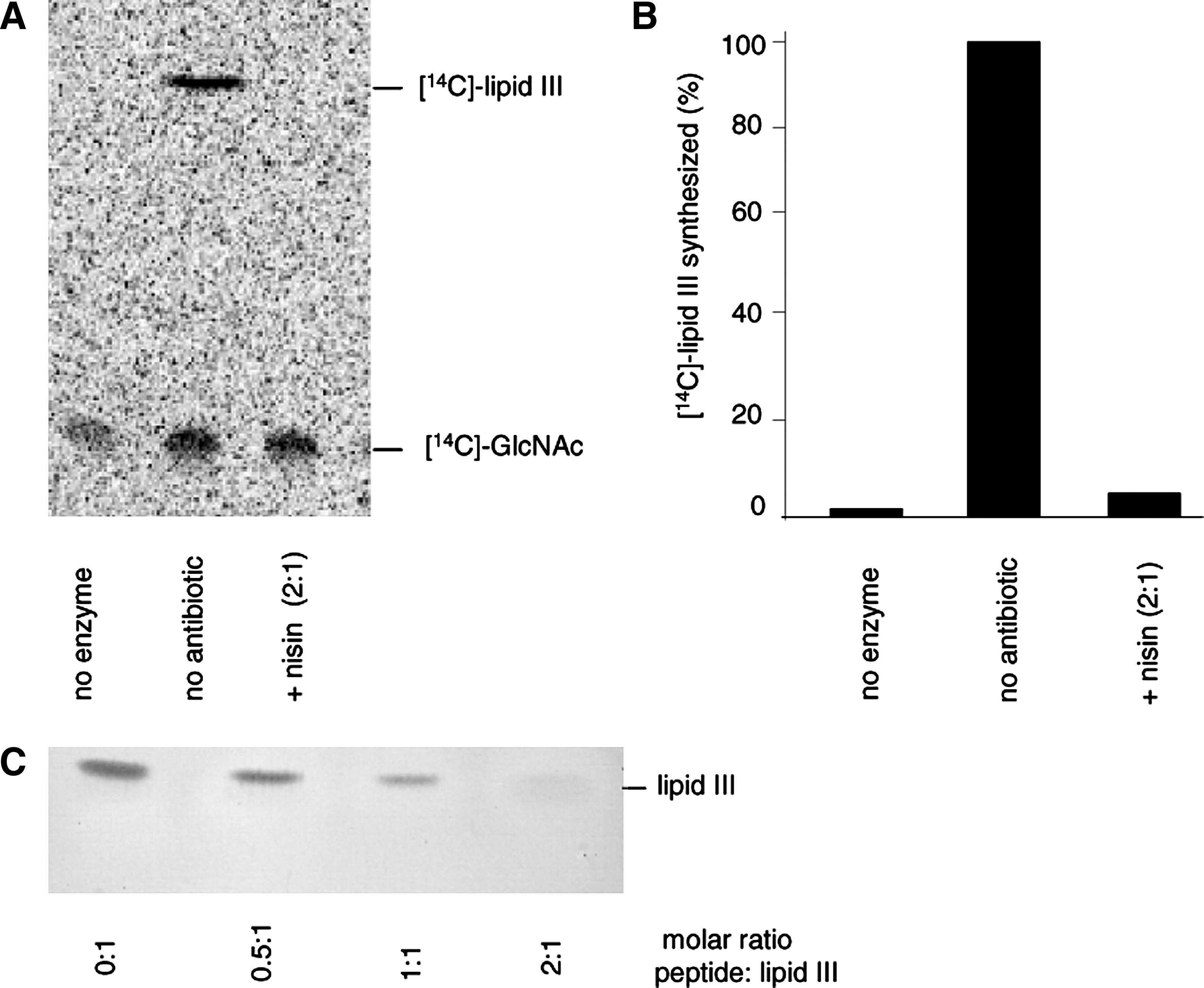
Nisin forms a stoichiometric complex with the WTA precursor lipid III.
No synthesis product could be detected when nisin was added at a twofold molar excess with respect to C55-P (lane 3), suggesting that nisin forms a stable complex with lipid III, which escapes extraction of the lipid intermediate from the synthesis mixture (lane 3), as described for the nisin:lipid II complex. 39
To further verify the formation of a nisin:lipid III complex, purified lipid III was incubated with defined amounts of nisin, followed by extraction and subsequent TLC analysis.
In accordance with the results obtained in the TagO synthesis assay, increasing nisin concentrations gradually diminished the amount of lipid III visible on the TLC plate and almost complete sequestration of lipid III was observed at a twofold molar excess of nisin (Fig. 2C). Similar results were obtained with gallidermin (data not shown).
Quantitative analysis of the TagA-catalyzed lipid IV synthesis reaction using radiolabeled UDP-GlcNAc revealed dose-dependent inhibition by nisin and gallidermin (Fig. 3). Added in equimolar concentrations, nisin and gallidermin inhibited the formation of lipid IV, compared with a positive control—where no antibiotic was added—to about 70% and 50%, respectively. At a molar ratio of 2:1 (nisin:lipid), nisin inhibited the reaction to about 20% and gallidermin prevented the synthesis of radiolabeled lipid IV almost completely. Again, with this experimental setup, it was not possible to differentiate whether the lantibiotics inhibit the synthesis of lipid IV by binding to the precursor lipid III, or whether they block the extraction of the reaction product by complexation. To obtain further information on the nature of the inhibition, we performed antagonization assays with nisin and gallidermin. We used conventional microbroth dilution for MIC determination and added the lipid intermediates in a 10-fold molar excess. Lipid III and lipid IV, as much as lipid I and lipid II, completely antagonized the activity of both lantibiotics (Table 2).

Impact of nisin (gray) and gallidermin (white) on the TagA-catalyzed lipid IV synthesis. TagA was incubated with purified lipid III and [14C]-UDP-GlcNAc. The reaction was initiated by the addition of 1.1 μg MnaA and TagA and incubated for 4 hours at 30°C. Lantibiotics were added at molar ratios of 1 and 2 with respect to the substrate lipid III. Reaction products were extracted with BuOH/PyrAc (2:1) and separated by TLC. Detection and quantification was carried out using a Storm Phosphoimager.
MICs, minimal inhibitory concentrations.
M. luteus DSM 1790 was incubated with 0.2 mg/L nisin and 0.04 mg/L gallidermin, corresponding to 10-fold the MIC; antagonists were added in 10-fold molar excess with respect to the antibiotics. (+) Antibiotic activity antagonized, (−) antibiotic activity unaffected.
C55-P, in contrast, had no impact, even when added at higher concentrations (25-fold excess; data not shown). This molecule lacks the pyrophosphate moiety, which is essential for the interaction with the nisin-binding motif. Nisin further trapped C55-PP in a stable complex that resided in the aqueous phase after extraction (Fig. 4). An almost complete sequestration of the lipid carrier C55-PP was observed when nisin was added in a twofold molar excess, pointing to a complex stoichiometry of 2:1, as seen with the other WTA and peptidoglycan intermediates.
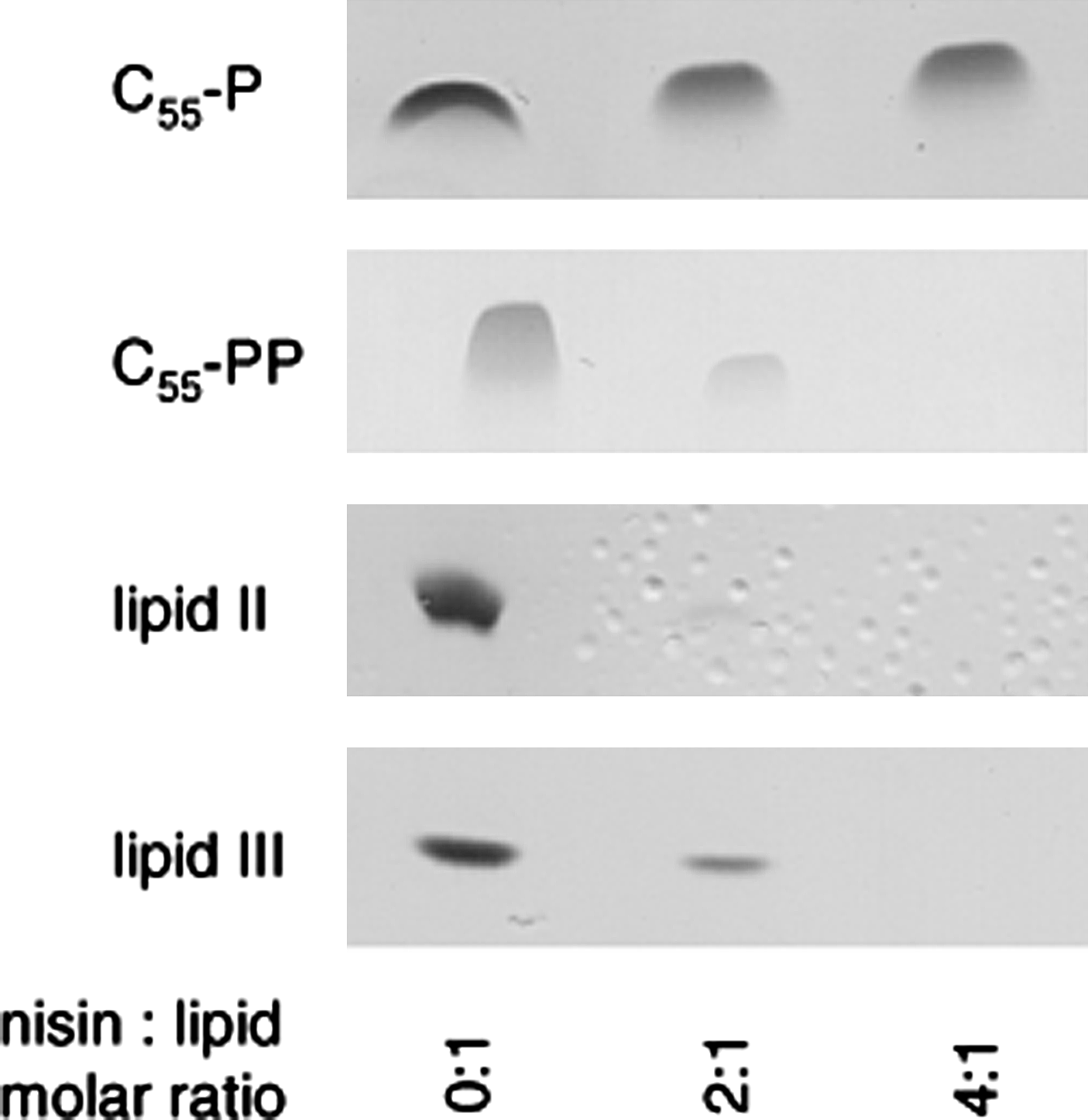
Complex formation of nisin with various cell wall precursors. Cell wall precursors C55-P, C55-PP, lipid II, and lipid III were incubated with increasing concentrations of nisin in molar ratios of 0, 2, and 4 with respect to the respective precursor. Reaction mixtures were extracted and unbound lipids were analyzed by TLC.
Determination of binding parameters
To determine the binding parameters of nisin for phospholipid bilayers doped with various lipid intermediates, we used the quartz crystal microbalance (QCM) biosensor technique. 13 Kinetic binding constants, such as kass, kdiss, and kD, were calculated from the QCM frequency curves (Table 3). Compared with bilayers composed of DOPC, nisin-binding affinity was significantly increased in the presence of 0.1 mol% lipid III or lipid IV. Association constants were increased up to sevenfold, in the presence of both WTA precursors, compared with pure DOPC membranes (Kass=751 M−1 seconds−1).
Association rate (kass), dissociation rate (kdiss), and binding affinity (kD) of nisin as determined by quartz crystal microbalance measurements.
In line with previous observations and the results obtained from the antagonization assays, binding to C55-P was not detectable. On the contrary, C55-PP-binding affinity is comparable to that of bactoprenol-bound lipid intermediates. However, nisin binding to C55-PP was distinguished by a less-pronounced association and a considerably slower dissociation compared to lipid II, lipid III, and lipid IV.
Nisin-induced CF efflux from lipid-III- and lipid-IV-doped liposomes
To investigate whether the interaction of nisin with lipid intermediates of WTA biosynthesis, that is, lipid III and lipid IV, facilitates the formation of target-specific pores, we prepared CF-loaded DOPC liposomes doped with 0.1 mol% of lipid III and lipid IV and monitored the efflux of the fluorescent dye after addition of nisin. As observed with lipid-I- and lipid-II-doped control vesicles, nisin induced an immediate and rapid CF release from liposomes doped with either lipid III or lipid IV (Fig. 5), reaching comparable values, that is, 50% CF release at 50 nM of nisin. As described for nisin and lipid II, 38 pore formation in lipid-III-doped vesicles was dependent on the concentration of both, peptide and lipid intermediate (data not shown).
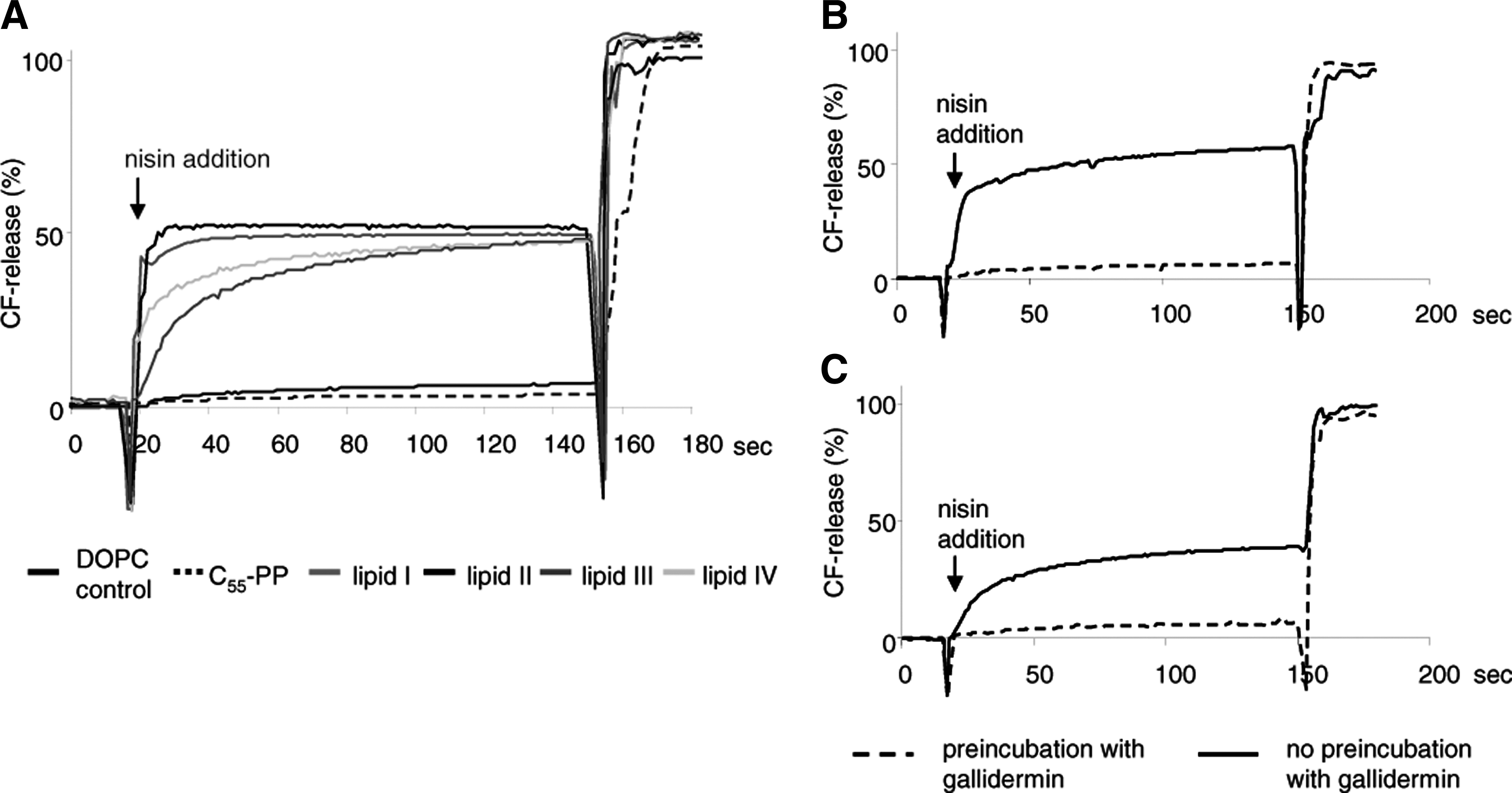
Nisin-induced pore formation. Nisin-induced carboxyfluorescein (CF) release from liposomes made of 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC; black), and DOPC supplemented with 0.1 mol% lipid I (blue), lipid II (red), lipid III (orange), lipid IV (green), and C55-PP (black, dotted). Nisin was added at a concentration of 300 μM. Marker release from the liposomes (25 mM phospholipid on a phosphorous basis) was determined for 3 minutes
Only minor marker release was observed with pure DOPC vesicles or vesicles doped with C55-PP. In the experimental system used here, gallidermin is unable to form pores, but was found to prevent nisin-induced pore formation by shielding lipid II. We therefore preincubated lipid-III- and lipid-IV-doped liposomes with gallidermin before nisin addition and found that its pore formation capacity was completely blocked.
The results provide evidence that WTA precursors can function as docking molecules for targeted pore formation in vitro.
Discussion
Lantibiotics are active against Gram-positive bacteria and most of them target components of the bacterial cytoplasmic membrane, with the majority of the known compounds exerting their antimicrobial activity by binding to lipid II. 2
Type B lantibiotics, that is, mersacidin, selectively inhibit peptidoglycan biosynthesis by targeting lipid II. 10 In contrast to this, type A lantibiotics, such as nisin, combine a multiplicity of activities in one molecule. The interaction with the cell wall precursor lipid II most efficiently blocks the assembly of a functional cell wall and may promote, dependent on the overall size of the individual lantibiotic, efficient target-mediated pore formation.8,10,38 Moreover, sequestration of lipid II profoundly impacts on the entire cell wall synthesis machinery, resulting in delocalization of the precursor from the septum, aberrant septum formation, and most likely the disorganization of multimeric protein complexes. 17
The various mechanisms combined in one lantibiotic molecule may potentiate the antibiotic activity and explain MIC values in the nanomolar range, as observed with some very sensitive bacteria, such as lactococci. 18 Moreover, development of resistance is gravely hampered, as different essential bacterial components are simultaneously targeted. 1
In this study we report the in vitro reconstitution of TagO and demonstrate that nisin and related type A lantibiotics interact with lipid-bound precursors of the WTA biosynthesis pathway. Moreover, specific binding enables subsequent target-mediated formation of pores in model membranes, similar to those promoted by lipid II binding.
Lipid II and the WTA intermediates differ in the identity of the first sugar being MurNAc or GlcNAc, respectively. Moreover, lipid II is characterized by a pentapeptide side chain attached to the first monosaccharide. Antimicrobial activity of nisin against VRSA (VanA type), already suggested that the terminal D-Ala-D-Ala terminus, is not involved in the interaction with nisin. 8 This hypothesis was further substantiated by nuclear magnetic resonance studies in which the solution structure of the nisin:lipid II complex was described. 20 Hsu et al. reported marginal chemical shift changes in the D-Ala-D-Ala moiety upon nisin binding. However, oligomerization of the pore complex in a lipid bilayer and underlying structural changes have not yet been analyzed in molecular detail. Residues different from those involved in primary interaction could be important for further complex assembly. Here, we provide experimental evidence that the entire pentapeptide side chain of lipid II is not involved in high affinity binding of nisin. Lipid III (C55-PP-GlcNAc) (kD=3.7*E-7) is efficiently bound by nisin, exhibiting binding constants in the same order of magnitude as observed for lipid II (kD=5.58*E-7). Moreover, the finding that lipid III enables targeted pore formation in vitro clearly demonstrates that the pentapeptide side chain is not involved in the dynamic process of nisin complex formation. Nisin binding to lipid II primarily involves the pyrophosphate moiety 20 ; however, a second binding site was deduced from the observation that methylated nisin is able to bind to lipid II, but not to C55-PP. 4 Our results restrict such a second binding site, relevant for efficient target-mediated pore formation, to the sugar backbone. Apparently, at least association with both binding sites, that is, the pyrophosphate and the first sugar as present in lipid I and lipid III, is essential for the formation of a stable pore. The absence of this second binding site in C55-PP might explain that this interaction does not facilitate the formation of pores. Nevertheless, C55-PP when added to a conventional MIC determination antagonized the antimicrobial activity of nisin and gallidermin. Incubation with increasing concentrations of nisin prevented the extraction of C55-PP, indicating the formation of a stable complex with a 2:1 stoichiometry in solution. Moreover, as revealed by QCM, nisin binding constants determined for lipid II and WTA precursors were in the same order of magnitude as for lipid bilayers doped with C55-PP. This allows for the conclusion that the primary site of interaction, the pyrophosphate cage, 20 is the one contributing most to affinity. As expected, QCM data obtained with C55-P indicated a severe drop in affinity compared with C55-PP. According to this, the proposed secondary site of interaction would influence pore formation by other mechanisms than just increasing binding affinity. Specific interaction with the sugar headgroup could be necessary for the proper orientation of the nisin molecule toward the lipid bilayer. Conformational changes upon interaction with the sugar moiety could also influence the oligomerization of the full-pore complex. However, the final experimental proof for existence of a specific secondary site of interaction is still lacking and other explanations for the importance of the first sugar for pore formation are conceivable. The C55-PP pyrophosphate moiety has an additional negative charge as it is not engaged in an ester binding with a sugar group. The 2:1 nisin:C55-PP complex should therefore have a different surface charge distribution than nisin complexes formed with WTA or peptidoglycan intermediates. This different charge distribution could impact on the assembly of the full-pore complex or reduce its stability. The lack of bulky sugar headgroups could also lead to formation of nisin:C55-PP pore that has a smaller diameter (nisin:lipid II pore diameter=2–2.5 nm 16 ), allowing no efflux of the relatively large CF molecules (MW: 367.32 g/mol).
Although C55-PP probably does not serve as docking molecule for the formation of a stable pore, it might contribute to nisin activity in vivo. The pyrophosphorylated lipid carrier is exposed at the cell surface after completion of the synthetic cycle and therefore readily accessible for the lantibiotic. Nisin binding would block the recycling of this central carrier molecule, thereby leading to a fatal depletion of the cellular pool (Fig. 6). Bacitracin, used as topical agent for eye and skin infections, is known to act by such a mechanism. 37 In contrast to nisin, which has been shown to bind its molecular targets via five hydrogen bonds, 20 Bacitracin sequesters C55-PP by electrostatic interactions in the presence of divalent Zn2+ ions. 37
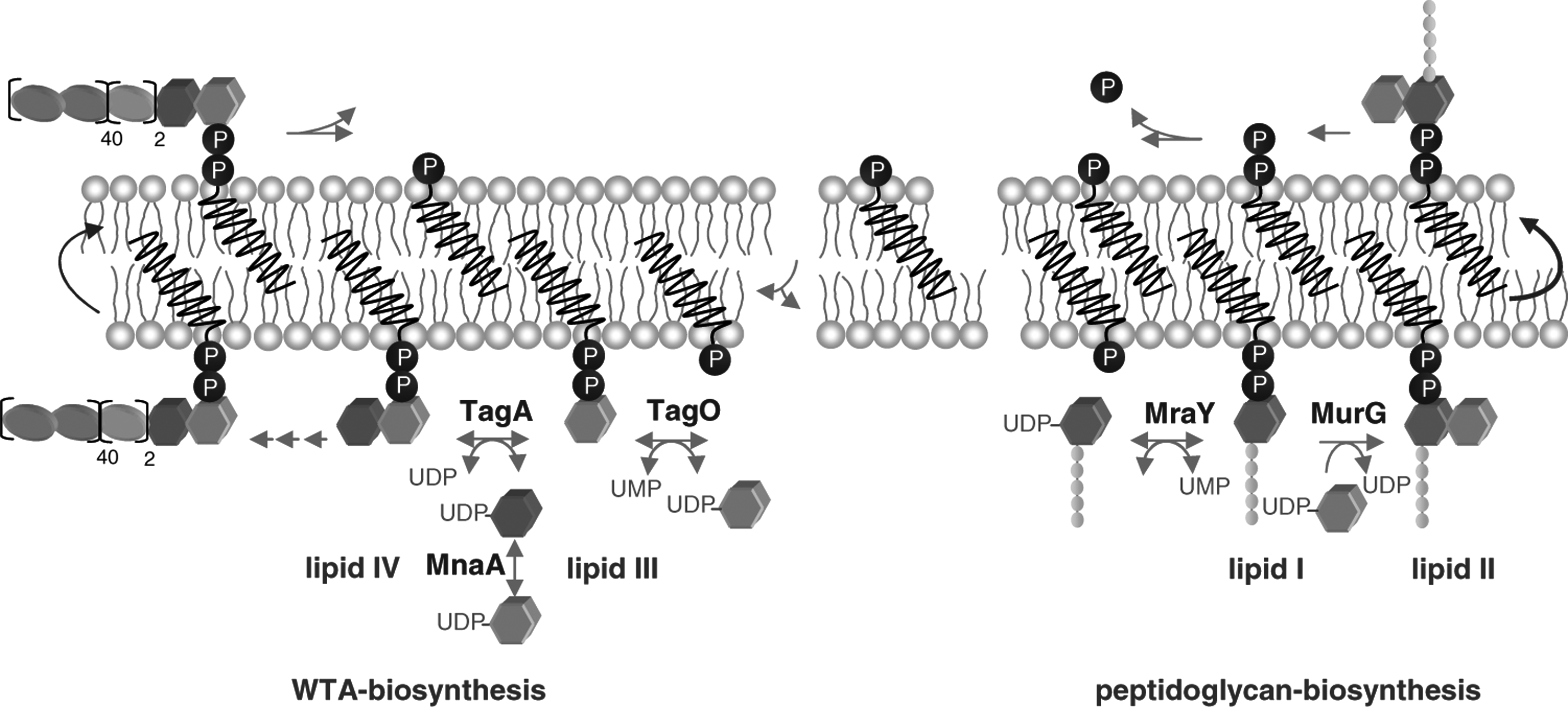
Schematic representation of peptidoglycan and WTA biosynthesis.
The accessibility of WTA precursors must be taken into account to evaluate their importance as targets in vivo. Nisin translocation across lipid bilayers has been observed in several studies.7,9,34 As lipid III and lipid IV are further processed in the cytoplasm, they could only function as targets after internalization of the lantibiotic. It is questionable how much this would contribute to killing in vivo as the lipid-II-facilitated mechanisms would induce much more rapid effects. The fully assembled WTA polymer seems to be the most promising candidate for an in vivo target, as it is the only WTA building block exposed at the cellular surface (Fig. 6).
Footnotes
Acknowledgment
The authors thank Michaele Josten for mass spectrometry analysis and nisin purification. This work was supported by the German Research Foundation (DFG; SA292/15-1, SCHN 1284/1-2, WI 1912/2-1) and the BONFOR Program of the Medical Faculty, University of Bonn.
Disclosure Statement
None of the authors has a financial relationship with a commercial entity that has an interest in the subject of this article.