
Abstract
Commensal Escherichia coli from healthy adult humans were screened for antibiotic resistance genes. Two unrelated strains contained the sul2 sulphonamide resistance gene and strAB streptomyicn resistance genes with the dfrA14 trimethoprim resistance gene cassette in the strA gene and conferred resistance to trimethoprim and sulphamethoxazole. A 6.8 kb plasmid, pCERC1, that contains these resistance genes was recovered and sequenced. Deletions were constructed, and the pCERC1 replication region was confined to a 1 kb segment carrying genes for RNAs that are closely related to the ColE1 replication initiation RNAs. Polymerase chain reaction assays, developed to detect the sul2-strA-strB gene cluster in this context, identified a streptomycin and sulphonamide resistance plasmid, pCERC2, identical to pCERC1 without the dfrA14 cassette in two further E. coli isolates. Bioinformatic analysis revealed plasmids similar to pCERC1 and two more members of this family. One, the probable progenitor, carries only the sul2 gene adjacent to the small mobile element CR2. The other has a variant resistance gene cluster that has evolved from pCERC2 via acquisition of the tet(A) tetracycline resistance determinant. pCERC1 and pCERC2 have been detected in many countries, indicating a global distribution and appear to have been circulating in Gram-negative bacteria for more than 25 years.
Introduction
Serovar Typhimurium DT104.
It has long been clear that antibiotic resistance genes spread rapidly around the world. However, in the last 10 years, it has become apparent that specific bacterial clones that have spread globally significantly contribute to the resistance burden. 26 There is also increasing evidence for the global spread of plasmids; for example, the small IncQ plasmid RSF1010 that carries the sul2-strA-strB-ΔCR2 configuration. 28 However, plasmids can be more difficult to track, as they are not confined to a single clone or species.
The large bacterial pools that constitute the commensal flora of humans and animals serve as reservoirs that can harbor antibiotic-resistant bacteria and the plasmids, genomic islands, transposons, and resistance genes that they carry. 1 Here, we have recovered plasmids carrying the dfrA14 cassette in strA from commensal Escherichia coli isolated from the feces of healthy Australian adults with no recent exposure to antibiotics, and examined their structure and their relationship to known plasmids and to plasmids in other commensal E. coli isolates.
Materials and Methods
E. coli strains
The commensal E. coli strains used in this study were recovered from subjects who had participated in a study that was approved by the University of Sydney Human Research Ethics Committee (04-2008/10778). Informed consent was obtained from all volunteers as per the requirements of the approval. Subjects were healthy adult members of the community (18–60 years), who had not taken antibiotics or traveled abroad within the previous 6 months. The strains were either derived from published studies1,2 or consisted of the antibiotic-resistant isolates from additional fecal samples. The characteristics of the isolates not reported 1 are shown in Table 2. Methods used to isolate, differentiate, and characterize the additional E. coli strains are described elsewhere. 1 The strains were assessed for resistance to 10 antibiotics, and the resistance genes conferring the observed resistance phenotypes were identified, as previously described. 1
Number of antibiotic resistant isolates per 100 E. coli isolates examined per fecal sample.
Numbers 1 and 2 indicate sul1 and sul2 or intI1 and intI2.
The context of the blaTEM gene has been previously described 2 except 3.6-R4, where the blaTEM gene is in Tn2. The context of the blaTEM gene in 3.6-R5 and 11.3-R3 could not be determined.
Letters indicate the type of the tetA gene.
strA interrupted by dfrA14. Plasmid pCERC1 present.
merA gene of Tn21 present.
Resistance variants of the same strain.
pCERC2 detected.
Rare isolates. Incidence was calculated from the total number of isolates recovered from Meuller–Hinton agar plates supplemented with sulphamethoxazole, as a percentage of the total abundance of the E. coli population in that fecal sample.
Resistance variants of the same strain.
No product with L1 and R1 primers.
Ap, ampicillin; Cm, chloramphenicol; Gm, gentamicin; Nx, nalidixic acid; Sm, streptomycin; Su, sulphamethoxazole; Tc, tetracycline; Tm, tobramycin; Tp, trimethoprim.
Plasmid recovery and analysis
Total plasmid DNA was extracted from 1.5 ml Luria broth cultures of the E. coli grown overnight at 37°C using the alkaline lysis miniprep method, 22 followed by extraction with phenol:chloroform:isoamyl alcohol (25:24:1) and chloroform:isoamyl alcohol (24:1), and precipitation with ethanol. DNA was re-suspended in 10 mM Tris HCl 0.1 mM EDTA (pH 8.0). Plasmid DNA was digested with restriction enzymes according to the manufacturer's instructions (New England BioLabs). Fragments were resolved by gel electrophoresis on 1% (w/v) agarose gel, stained with ethidium bromide (5 μg/ml), and visualized using a GelDoc1000 image analysis station (BioRad). The predicted fragment sizes were calculated from available sequences, and fragment sizes were estimated using 100 bp or 1 kb ladders (New England BioLabs).
Plasmids were recovered by transformation into chemically competent E. coli DH5α, prepared using calcium chloride according to standard protocols 22 with selection on Mueller Hinton agar containing sulphamethoxazole (100 μg/ml) or trimethoprim (25 μg/ml). Colonies were screened for their resistance to streptomycin (25 μg/ml), sulphamethoxazole, and trimethoprim as well as for their resistance to any other antibiotic indicated by the phenotype of the donor strain.
Construction of deletions
Deletions were constructed by digesting pCERC1 with restriction enzymes and re-ligating the fragments using T4 DNA Ligase according to the manufacturer's instructions (New England Biolabs). If necessary, the ends were filled using T4 polymerase according to the manufacturer's instructions (New England BioLabs). The re-ligated fragments were transformed into DH5α, selecting them for resistance to sulphamethoxazole (100 μg/ml). The constructs were verified by digestion with restriction enzymes and sequencing across the boundaries of the re-ligated fragments.
Plasmid stability
Plasmid stability was determined by measuring the loss of antibiotic resistance from cells grown in the absence of antibiotic selection. Overnight cultures were diluted 1,000-fold into fresh medium each day for 25 days. The fraction of antibiotic susceptible cells was measured by plating on medium with and without sulphamethoxazole, and by picking and patching 100 colonies from the antibiotic-free plates to determine their resistance phenotype.
Polymerase chain reaction, DNA sequencing, and sequence analysis
Polymerase chain reaction (PCR) amplification was carried out as previously described 1 using primers previously published 1 or primers described in Table 3. Fragments were resolved as just described.
Amplifies the resistance region flanked by the sequences present in pCERC1 and p9123.
The PCR products used as sequencing templates were purified by ethanol precipitation or were recovered from gels using the QIAquick Gel Extraction Kit (Qiagen). Automated sequencing was performed at the Sydney Node of the Australian Genome Research Facility on an AB3730xl DNA Analyzer, and sequences were assembled using Sequencher version 4.10 (Gene Codes Corporation). Sequenced regions were identified using Basic Local Alignment Search Tool (BLAST) or the BLAST paired alignment facility (http://blast.ncbi.nlm.nih.gov). Gene Construction Kit version 2.5 (Textco) was used to create figures to scale.
GenBank accession number
The 6.8 kb sequence of pCERC1 was submitted to GenBank under accession number JN012467.
Results
Incidence and linkage of the strAB and sul2 genes
A set of 25 E. coli isolates that were resistant to at least one of the antibiotics screened for were recovered from fecal samples from healthy Australian adults who had not recently taken antibiotics (Collection 2; Table 2). They were screened for the presence of antibiotic resistance genes as previously described. 1 Eleven strains carried the strA and strB genes adjacent to one another and also contained the sul2 gene. In 8 cases, the genes were linked in the order sul2-strA-strB. Seven further isolates from a collection of antibiotic-resistant E. coli (Collection 1) previously described 1 carried the strA, strB, and sul2 genes, and in six of these, the sul2-strA-strB cluster was present, bringing the total of this type to 14. Three isolates in collection 1 and one in collection 2 carried only the sul2 gene, but no isolates with only strA and strB were detected in either set.
dfrA14 in strA
In two of the E. coli from Collection 2 (1.2-R3 and 6.2-R1), the amplicon obtained for the strA gene was 0.6 kb larger than predicted. These isolates were recovered from different subjects, and had different characteristics (phylogenetic group, API20E profile, RAPD profiles; data not shown), and, hence, were not related. Digestion of the larger amplicons with EcoRI demonstrated that they were the same, and the sequence revealed the presence of the 568 bp dfrA14 gene cassette in a secondary site in the strA gene. The sequence was identical to the ones previously reported for plasmids pUK1329 30 and pSTOJO1, 18 which were recovered from clinical E. coli isolates in Scotland and Nigeria, respectively, as well as from clinical isolates in Spain and commensal E. coli from Kenyan animals (Table 1). Linkage PCRs demonstrated that the interrupted strA was flanked on one side by strB and on the other by sul2, and the sequence of this region was identical to that reported for plasmid pSTOJO1. 18 Where sequence was available, the dfrA14 gene cassette is in precisely the same position, indicating that they result from a single cassette integration event into a secondary site. 20
pCERC1
The E. coli isolates 1.2-R3 (SuRTpRTcR) and 6.2-R1 (SuRTpR) were resistant to sulphamethoxazole and trimethoprim, but did not exhibit resistance to streptomycin. Plasmids conferring resistance to trimethoprim and sulphamethoxazole were recovered from both E. coli strains. These small plasmids exhibited the same pattern of fragments after digestion with restriction enzymes EcoRV and PstI. Since no plasmid carrying the dfrA14 gene in a secondary site had been completely sequenced (see Table 1), one of these plasmids, designated pCERC1 (
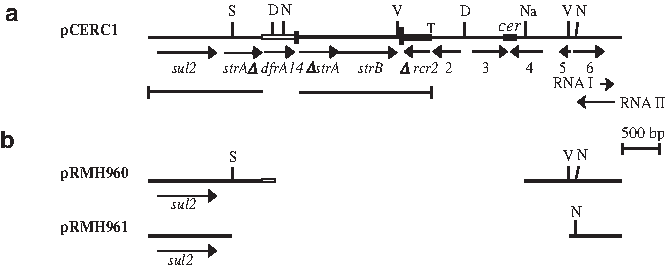
Map of
Plasmids related to pCERC1
When the pCERC1 sequence was compared with other sequences available in the GenBank non redundant DNA database (last search 19th October, 2011), the closest match was for a streptomycin and sulphonamide resistant plasmid p9123 recovered before 2003 from a clinical E. coli isolate in the United Kingdom. 6 Plasmid p9123 lacks the dfrA14 but is 99.99% identical (3 differences in 6,222 bp) to pCERC1 with all bases surrounding the cassette present in it (Fig. 2), confirming that the cassette was integrated into a plasmid identical to p9123. Plasmid p9123 was the first 6 of several plasmids identical or nearly identical to it to be found in E. coli, Shigella, and Salmonella recovered in many different countries (Table 4). A variant form of p9123, named both pE2348-2 and p6148, that fails to confer resistance to sulphonamides due to a premature stop codon in the sul2 gene was recovered from a 1969 enteropathogenic E. coli isolate.10,13
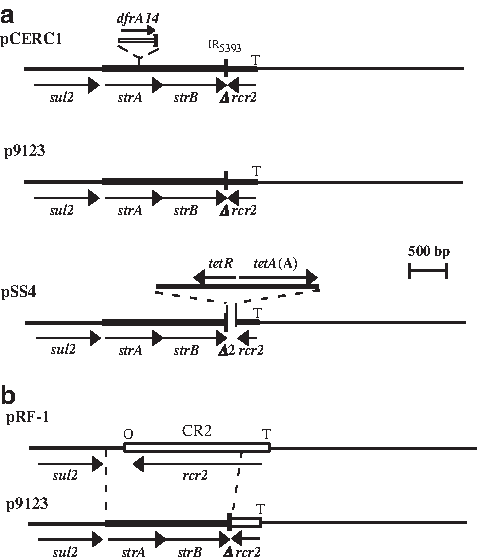
The p9123 plasmid family.
Human isolates are clinical except for pCERC1.
Sequence contains 3 SNPs relative to other plasmids in the group.
Date of isolation was not provided; < denotes before the GenBank entry date.
The earliest mention of the strain was in 1980.
The earliest mention of the strain was 1999.
Cervus elaphus (wild living).
Isolated from E. coli ETEC strain E2348/69. A +1 frameshift mutation has inactivated the sul2 gene.
serovar Choleraesuis.
Two further variants were also found (Table 4 and Fig. 2). In one, the strA and strB genes are missing, and the CR2 element is complete (Fig. 2b). This form of the resistance region is the predicted precursor of the more complex sul2-strA-strB configuration. 28 The final variant type (Fig. 2a) includes a tet(A) tetracycline resistance determinant adjacent to the IR of the portion derived from Tn5393, and a part of the CR2 remnant has been lost. A close inspection of the boundaries in this variant type (Fig. 3) revealed that the IR of Tn5393 abuts one end of the tet(A) fragment, which is identical to other sequences for this tetracycline resistance determinant. At the other end, the junction has been formed by recombination between a very short region of identity (8/9 bases or 11/14 bases) near the end of the tetA(A) gene and within CR2 (Fig. 3). The latter event has led to the replacement of the last six codons of the tetA(A) gene, and the six amino acids are replaced by six alternate amino acids encoded by the CR2-derived segment.

tetA(A)-CR2 boundary in pSS4. The nucleotides in the crossover region are bold and boxed. The stop codon of the tetA(A) gene is underlined. The sequence of pSS4 (GenBank accession number) has been aligned with Tn1721 and CR2 (GenBank accession numbers ×61367 and JN247441).
Identification of pCERC1 replication and stability functions
To determine which portion of the 3.5 kb lying outside the resistance region was required for plasmid replication, deletions were constructed. Two sulphonamide resistance plasmids containing large deletions were recovered (Fig. 1b), confining the replication functions to a region of 1.1 kb that included only orf6. This finding provides support for the previous identification of genes for RNAs equivalent to RNAI and RNAII of ColE1, which regulate replication initiation in this region. 11
A cer site, which is believed to increase plasmid stability by allowing multimer resolution by the E. coli XerCXerD site-specific recombination system, 24 was also identified between the orf3 and orf4 (Fig. 1a). 11 However, when cells containing pCERC1 or the deletion derivatives, pRMH964 and pRMH965, both of which lack the cer site, were grown without antibiotic selection for 250 generations, no loss of plasmid was detected. This suggests that these plasmids are very stably maintained in the absence of the cer site, perhaps due to a very high copy number. 11 Hence, dimer resolution at the cer site does not appear to significantly contribute to their stability.
pCERC2
In order to determine whether any of the variant plasmids identified by bioinformatic analysis are present in the commensal E. coli collections, isolates containing sul2 with either strA and strB or with sul2 alone were screened for the presence of a resistance segment flanked by a sequence found in the pCERC1 backbone using primers RH963 and RH964 (Table 3). Two strains, 3.6-R4 (ApRSmRSuR) from collection 2 and 10.1-R1 (ApRSmRSuRTcRTpR) from collection 1, 2 yielded amplicons of 3.6 kb, and 6.2 kb plasmids conferring resistance to sulfamethoxazole and streptomycin were recovered from both of them. These plasmids exhibited EcoRI and PstI digestion patterns identical to those predicted for p9123. Amplicons of the predicted size were also obtained using PCRs spanning the whole plasmid (see Table 3). These plasmids, which were designated pCERC2, are likely to be identical or nearly identical to p9123.
Discussion
The dfrA14 gene in plasmid pCERC1 accounts for the trimethoprim resistance of two of the three strains in our collections that conferred resistance to trimethoprim, but did not include a class 1 or a class 2 integron carrying a dfrA gene cassette (see 1 and Table 2). The small plasmids pCERC1 and pCERC2 account for the sul2-strA-strB-ΔCR2 configuration in four of the 14 strains from collections 1 and 2 that carry it. In a further strain (1-R1), we have previously shown that it is a part of the transposon Tn6029Δ. 2 The location of these genes in the remaining ten isolates carrying sul2-strA-strB-ΔCR2 remains to be established. Moreover, how the sul2-strA-strB-ΔCR2 configuration has moved to the several different locations where it has already been found remains to be established.
We have previously proposed a scheme for creation of the sul2-strA-strB-ΔCR2 from the transposon Tn5393, which carries strA and strB adjacent to one another, and the sul2 gene located adjacent to CR2. 28 This scheme is consistent with the configurations found in the set of related plasmids described here, one of which carries sul2 adjacent to CR2. Plasmid pSS4, the additional member of the set of small plasmids described here, carries an additional segment consisting of the tet(A) tetracycline resistance determinant and adjacent sequence, as found in the IncP1α plasmid RP1 and its close relatives. 19 Analysis of the boundaries between the new portion of this plasmid and the surrounding p9123-derived region revealed that it arose via steps that are very similar to those by which the sul2-strA-strB-ΔCR2 arose. 28
We suggest that this set of small plasmids with the same ColE1-like replication initiation functions but with various resistance gene clusters be named the p9123 family after the first member to be completely sequenced. 6 Members of this family appear to be widely distributed in Gram-negative bacteria, and they have previously been found in human clinical isolates as well as in bacteria recovered from animals (Table 4). Here, two types from this plasmid family were found in a relatively small collection of commensal E. coli from Australian adults in the community, indicating that they are quite abundant. This is consistent with the findings of an earlier study that found a streptomycin and sulphonamide resistance plasmid, pBP1, was abundant in E. coli isolates from Germany, Egypt, England, the USSR, Mexico, and Japan that conferred resistance to these antibiotics. 15 Though a sequence for pBP1 is not available, the restriction map is essentially the same as that predicted from the p9123 sequence. These small plasmids do not encode any protein identified as a mobilization protein, and their ability to disseminate widely in Gram-negative bacteria may be due to natural transformation.
Plasmids in the p9123 family also appear to be stably maintained in the Gram-negative bacterial population without antibiotic selection. Our subjects had not taken antibiotics for at least 6 months before sampling, and our experiments were unable to detect significant loss of the plasmids in the absence of antibiotic selection. Though a fitness advantage has been reported to be associated with the presence of p9123, 6 this was not observed in a later study, 10 and further studies will be needed to clarify this point.
Footnotes
Acknowledgment
The authors thank the study participants who donated the fecal samples. They thank Jannine Bailey and Jeremy Pinyon for assistance with the characterization of some isolates shown in ![]() . This study was supported by the NHMRC Project grant 512434. R.M.H. was supported by the NHMRC Fellowship grant 358713. S.A. was supported by an Australian Postgraduate Award.
. This study was supported by the NHMRC Project grant 512434. R.M.H. was supported by the NHMRC Fellowship grant 358713. S.A. was supported by an Australian Postgraduate Award.
Disclosure Statement
No competing financial interests exist.