
Abstract
We examined the prevalence and diversity of carbapenem-resistant bacteria (CRB) in untreated drinking water. Prevalence was estimated in plate count agar (PCA) and R2A media with or without antibiotics. Clonal relatedness of isolates was established by repetitive extragenic palindroitic (REP)-PCR. Phylogeny was based on the 16S rRNA gene. Antimicrobial susceptibility was assessed by disc diffusion methods. Genes encoding beta-lactamases and integrases were inspected by PCR. CRB ranged from 0.02% to 15.9% of cultivable bacteria, while ampicillin-resistant bacteria ranged from 1.5% to 31.4%. Carbapenem-resistant isolates affiliated with genera Stenotrophomonas, Pseudomonas, Janthinobacterium, Chryseobacterium, Sphingobacterium, Acidovorax, Caulobacter, Cupriavidus, and Sphingomonas. CRB were highly resistant to beta-lactams, but mostly susceptible to other classes. Transmissible beta-lactamase genes and integrase genes were not detected. The genus-specific blaL1 was detected in 61% of the Stenotrophomonas isolates. Contrarily to what has been reported for extensively used antibiotics, low levels of carbapenem resistance were detected in untreated drinking water, often represented by intrinsically resistant genera. Production of chromosomal-encoded carbapenemases was the prevalent carbapenem resistance mechanism. Results suggest that the dissemination of anthropogenic-derived carbapenem resistance is at an early stage. This presents an opportunity to rationally develop monitoring strategies to identify dissemination routes and assess the impact of human actions in the environmental resistome.
Introduction
In Portugal, potable but untreated water (e.g., from public fountains) is commonly used for human needs, particularly in rural and small urban communities.17,29 Water from these fountains is considered pristine because, according to routine implemented surveillance, anthropogenic-derived contaminants are not present in significant amounts. However, as in other countries, the regular microbial quality control following the legislation guidelines does not include any parameter related to AR. 6 Hence, untreated drinking water may be a relevant, but mostly overlooked, vehicle for dissemination of AR; deciphering resistance prevalence as well as phylogenetic diversity of resident-resistant bacteria is an essential step to assess potential risks for human health.
Carbapenems are among the most powerful beta-lactams and one of the last therapeutic choices to fight multidrug-resistant gram-negative rods. 19 However, the emergence of carbapenem resistance in clinical settings may compromise the efficacy of these antibiotics.3,19 Resistance to carbapenems is frequently mediated by production of transmissible or chromosomal carbapenemases (for review see refs.3,28). The most common transmissible carbapenemases belong to molecular class B and specifically to IMP and VIM families. 3
Studies focusing on AR in environmental bacteria are frequently restricted to the characterization of resistance toward the more intensively used antibiotics. In contrast, little attention has been paid to resistance to last-resort antibiotics, such as carbapenems. As the use of these antibiotics has been limited, we hypothesize that the spread of resistance may be at an earlier stage especially in pristine environments such as potable untreated drinking water. To confirm this hypothesis, we examined the prevalence and diversity of carbapenem-resistant bacteria (CRB) and resistance genes in untreated drinking water.
Materials and Methods
Samples
Water samples were collected from 12 fountains (designated A–L; geographic coordinates are shown in Table 1) between October 2009 and February 2010. Fountains were distributed in the North and Central regions of Portugal within an area of ∼6,000 km2. Fountains J, K, and I were located in rural regions, while fountains A–H and L were located in urban areas. Samples were kept on ice and processed within 2–4 hr after collection. Selected fountains are regularly evaluated by local authorities to confirm drinking water safety according to the Portuguese legislation. 6
ND, not determined. PCA, plate count agar; CFU, colony-forming unit.
Prevalence of culturable CRB and ampicillin-resistant bacteria
To enumerate the total number of colony forming units (CFUs), samples were filtered through 0.45-μm cellulose ester membranes (Pall Life Sciences). Filters were placed on either R2A agar or plate count agar (PCA) without antibiotics. Plates containing 50 μg/ml of ampicillin or 5 μg/ml of imipenem were used to determine the prevalence of bacteria resistant to each antibiotic. Triplicates were counted after incubating at 37°C for 72 hr (R2A) or 25°C for 48 hr (PCA).
Isolation and identification of CRB
Individual colonies grown on media supplemented with imipenem were selected, purified, and stored in 15% glycerol at −70°C. DNA was obtained from pure cultures by using the Genomic DNA purification kit (MBI Fermentas). Bacterial isolates were typed by REP-PCR to exclude clones. 27 One isolate representing each REP profile was selected and identified by sequencing the 16S rRNA gene. 15
Phylogenetic analysis
Phylogenetic relationships were determined by comparing the 16S rRNA gene sequences obtained with sequences from the GenBank. Sequences from the closest relatives as well as the closest type strains, based on a BLAST search, 1 were selected as reference sequences. Nucleotide sequences were aligned by CLUSTAL X. 25 The analysis was performed by the neighbor-joining method (Kimura two-parameter distance optimized criteria) using MEGA version 5.0. 23 The robustness of the tree was confirmed by bootstrap analysis based on 1,000 resamplings. 8
Antimicrobial susceptibility test
AR profiles were determined by disk diffusion in accordance with the Clinical and Laboratory Standards Institute (CLSI). 5 Discs (Oxoid) contained amoxicillin (10 μg); amoxicillin/clavulanic acid (20 μg/10 μg); piperacillin (100 μg); piperacillin/tazobactam (100 μg/10 μg); cephalothin (30 μg); cefotaxime (30 μg); ceftazidime (30 μg); aztreonam (30 μg); imipenem (10 μg); nalidixic acid (30 μg); ciprofloxacin (5 μg); chloramphenicol (30 μg); gentamicin (10 μg); kanamycin (30 μg); streptomycin (10 μg); sulfamethoxazole/trimethoprim (25 μg); and tetracycline (30 μg). The strain Escherichia coli ATCC 25922 was used as quality control. The isolates were classified as susceptible or resistant according to CLSI guidelines. 5 Isolates displaying intermediate susceptibility to an antibiotic were classified as susceptible. When taxa-specific recommendations were unavailable, the resistance breakpoints defined by CLSI for Enterobacteriaceae were applied.
Detection of resistance genes and integrases
All isolates were screened for the presence of commonly acquired beta-lactamase genes (blaTEM, blaSHV, and blaOXA) and of the most frequent transmissible carbapenemase genes (blaIMP and blaVIM) as previously described. 12 The presence of integrons, frequently associated with blaIMP and blaVIM, was assessed by detecting integrase-coding genes intI1 and intI2. 12 Stenotrophomonas isolates were also screened for the genus-specific carbapenemase gene L12 using primers L1_F (5′-CACCTGGCAGATCGGCAC-3′) and L1_R (5′-CTGGCGCCCGGATGCGG-3′) and an annealing temperature of 56°C. Amplification was carried out in a MyCycler Thermal Cycler (Bio-Rad Laboratories), using reagents purchased from MBI Fermentas. PCR experiments contained a positive control (strains carrying resistance genes or integrons) and a negative control (sterile water instead of DNA). Amplified products were separated by 1% agarose gel electrophoresis and visualized by ethidium bromide staining.
Results
Colony-counting results are summarized in Table 1. Total viable colony counts ranged from 2×102 to 7×104 CFU per milliliter (in PCA) and from 2.3×102 to 1.2×104 CFU per milliliter (in R2A). Imipenem-resistant bacteria accounted for 0.02–15.9% (in PCA) and 0.12–7.5% (in R2A) of the total bacteria. Ampicillin-resistant bacteria accounted for 8.4–31.2% (in PCA) and 1.5–31.4% (in R2A).
Imipenem-resistant isolates were recovered from all the sampled fountains. A total of 68 isolates were analyzed by REP-PCR and one representative of each profile was identified by partial 16S rRNA gene sequence analysis (Fig. 1 and Table 2). The 36 identified isolates were most closely related to nine genera in five classes of bacteria. Twenty-two isolates grouped with species of Gammaproteobacteria and were most closely related to Stenotrophomonas (19 isolates; 52.7%) and Pseudomonas (three isolates; 8.3%). Eight isolates fell into class Betaproteobacteria and were closely related to genera Janthinobacterium (three isolates; 8.3%), Ralstonia (three isolates; 8.3%), Acidovorax (one isolate; 2.8%), and Cupriavidus (one isolate; 2.8%). Two isolates affiliated with Alphaproteobacteria within genera Caulobacter (one; 2.8%) and Sphingomonas (one; 2.8%). Classes Flavobacteriia and Sphingobacteriia were represented by two isolates each, belonging to genera Chryseobacterium (two; 5.6%) and Sphingobacterium (two; 5.6%), respectively.

Phylogenetic tree showing relatedness between imipenem-resistant isolates and selected strains and environmental clones. The first letter of the isolates name corresponds to the sampled fountain (A–L) and the second corresponds to the culture media (R-R2A or P-PCA). The number of isolates with similar REP-profiles is shown in parenthesis. The branch numbers refer to the percent confidence (>50%) as estimated by a bootstrap analysis with 1,000 replications. Scale bar indicates the number of nucleotide substitutions per site. PCA, plate count agar.
AML, amoxicillin; AMC, amoxicillin/clavulanic acid; ATM, aztreonam; PRL, piperacillin; TZP, piperacillin/tazobactam; KF, cephalothin; CAZ, ceftazidime; CTX, cefotaxime; IPM, imipenem; CN, gentamycin; K, kanamycin; S, streptomycin; SXT, sulfamethoxazole/trimethoprim; NA, nalidixic acid.
Strains were screened for resistance against 17 antibiotics. The frequencies of AR are presented in Fig. 2. Besides imipenem, high levels of resistance against other beta-lactams were registered, especially against amoxicillin (94.4% resistant strains), cephalothin (91.7%), and aztreonam (86.1%). In fact, 72% of the isolates were resistant to at least six beta-lactams. Besides beta-lactams, the most effective antibiotics were nalidixic acid (33% resistant bacteria) and streptomycin (28%). Of the 36 strains, 38.9% were only resistant to beta-lactams and 52.8% were resistant to antibiotics belonging to two classes, namely, beta-lactams and quinolones (nine isolates) or beta-lactams and aminoglycosides (10 isolates). Multiresistance (resistance to three or more classes of antibiotics) was found in only three isolates (Table 2). Among these isolates, two affiliated with Chryseobacterium (strains AR1 and CP4) and were resistant, besides beta-lactams, to nalidixic acid and to the combination sulfamethoxazole/trimethoprim. The other isolate (LR3) was identified as Stenotrophomonas and was resistant to beta-lactams, kanamycin, and nalidixic acid. The most common resistance profile was shared by seven Stenotrophomonas strains and included all tested beta-lactams plus streptomycin and kanamycin. The antibiotics with the highest activity (100% sensitivity) were ciprofloxacin, chloramphenicol, and tetracycline.
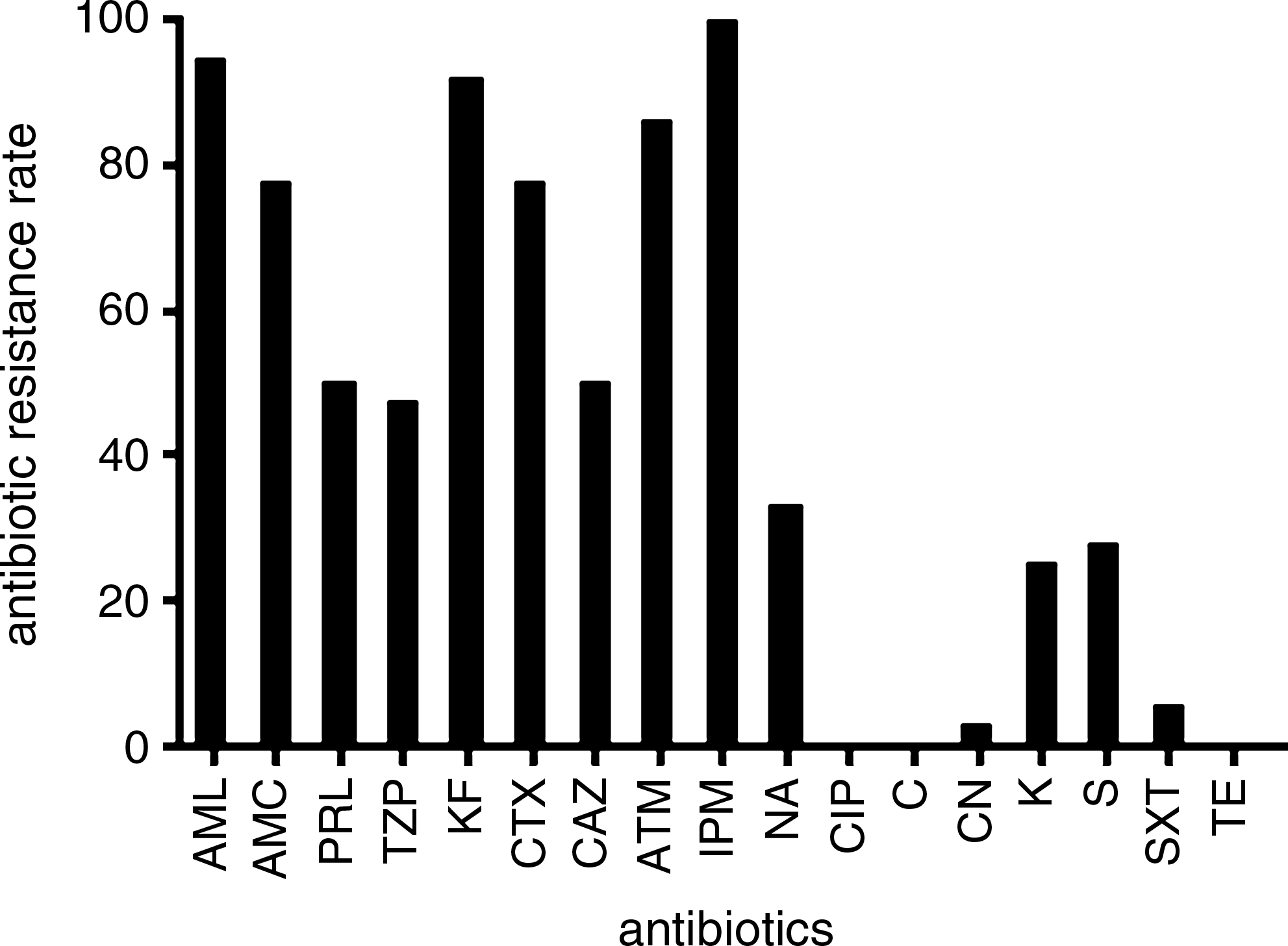
Frequencies of antibiotic resistance of the imipenem-resistant isolates in public fountains. AML, amoxicillin; AMC, amoxicillin/clavulanic acid; PRL, piperacillin; TZP, piperacillin/tazobactam; KF, cephalothin; CTX, cefotaxime; CAZ, ceftazidime; ATM, aztreonam; IPM, imipenem; NA, nalidixic acid; CIP, ciprofloxacin; C, chloramphenicol; CN, gentamycin; K, kanamycin; S, streptomycin; SXT, sulfamethoxazole/trimethoprim; TE, tetracycline.
Common acquired beta-lactamase genes (blaTEM, blaSHV, blaIMP, blaVIM, and blaOXA) were not detected in any of the isolates. Also, the integrase genes intI1 and intI2 were not found. The genus-specific carbapenemase gene blaL1 was detected in 61% of the Stenotrophomonas isolates (Table 2).
Discussion
The dissemination of resistance to last-resort antibiotics in the environment may have been delayed due to restrictions to their use. Our purpose was to examine the prevalence and diversity of CRB in untreated drinking water. We selected this environment as an example of pristine ones and also because the presence of antibiotic-resistant bacteria in drinking water can pose serious threats to human health. These bacteria may be transferred to humans via consumption, contributing to the spread of AR in pathogenic bacterial populations. Several studies have reported that antibiotic-resistant bacteria and resistance genes occur in drinking water.10,17,30 We added to those studies by specifically assessing resistance to last-resort antibiotics, namely, carbapenems. Water from the fountains here evaluated meets the quality standards according to national surveillance programs. However, surveillance programs ignore the presence and diversity of antibiotic-resistant bacteria.
We found low levels of carbapenem resistance in our samples. Moreover, the occurrence rates of CRB were lower than those of ampicillin-resistant bacteria. These results support the hypothesis that resistance to last-resort antibiotics is infrequent in ecosystems that have rarely been exposed to anthropogenic-derived contaminants. On the contrary, as shown here and by previous studies,16,30 resistance to widely used antibiotics is widespread even in pristine environments.
Even so, phylogenetically diverse CRB were isolated from our samples. Isolates belonged to genera that are common residents of freshwater environments 18 and naturally resistant to carbapenems. Stenotrophomonas was by far the most prevalent genus. Members of this genus and especially the species Stenotrophomonas maltophilia are intrinsically resistant to several antibiotics, including most beta-lactams. 21 Resistance to carbapenems in these bacteria results from the expression of the class B carbapenemase L1. 2 This gene was detected in 61% of our isolates. Negative results for the remaining Stenotrophomonas isolates may be attributed to the high genetic diversity of the L1 gene. 2 This heterogeneity may prevent annealing of the primers, even though they were designed based on most of the L1 sequences currently deposited in the GenBank.
Likewise, intrinsic carbapenem resistance mediated by chromosomal carbapenemases has been described in members of the genera Chryseobacterium, Janthinobacterium, Caulobacter, and Ralstonia.4,7,11 Isolates affiliated within these genera were also retrieved from our samples. Thus, our results suggest that the carbapenem resistance mechanism prevalent in untreated drinking water is the production of genus-specific chromosomal carbapenemases. Furthermore, the burden of carbapenemase-producers in drinking water may have been underestimated by the use of imipenem-supplemented media, since production of these enzymes may not always confer an in vitro resistance phenotype.
Transmissible genes are frequently co-selected with other resistance genes located in the same mobile elements resulting in multiresistance phenotypes. 22 Such phenotypes were uncommon in the isolated strains. Furthermore, we also have not detected genes encoding the most common transmissible beta-lactamases. These results, together with the dominance of intrinsically carbapenem-resistant genera, indicate that acquired resistance to these antibiotics is still scarce in bacteria inhabiting untreated drinking water. On the contrary, acquired resistance determinants to widely used antibiotics have previously been detected in similar environments. 17
A possible limitation of this study is the low number of analyzed isolates. However, this is a consequence of the low prevalence of CRB. Twelve fountains were sampled and all the colonies growing on imipenem-supplemented agar were analyzed. Also, two culture media were used to extend the diversity of targeted bacteria: the standard PCA medium and also the R2A, used for slow-growing bacteria that may be suppressed in nutritionally richer media. 26 This group of isolates is, therefore, representative of the culturable carbapenem-resistant community present in untreated drinking water.
The low levels of carbapenem resistance found in this study may be related to the infrequent use of these drugs in Portugal. In fact, use of imipenem is restricted to the clinical setting and only for the treatment of serious infections in hospitalized patients. 14
Conclusions
To our knowledge, this is the first attempt to specifically define the status of dissemination of resistance to last-resort antibiotics in drinking water. We found low levels of carbapenem resistance, mainly represented by genera previously described as intrinsically resistant to these antibiotics. Despite the possible methodological or regional drawbacks that may hamper definitive conclusions, our results suggest that the dissemination of transmissible carbapenem resistance in pristine environments, and specifically in drinking water, is at an earlier stage, simulating a pre-antibiotic era situation. This presents an opportunity to implement monitoring strategies to follow and clarify dissemination routes and distinguish between naturally occurring and human-derived resistance. Surveillance data, along with other preventive measures, will contribute to control the anthropogenic-driven dissemination of antibiotic-resistant bacteria in drinking water and to anticipate public-health problems.
Footnotes
Acknowledgments
Authors are grateful to Ana Luísa Ramos for assistance on sampling. This work was supported by Fundação para a Ciência e a Tecnologia in the form of grants to I. Henriques (SFRH/BPD/63487/2009), J. Azevedo (SFRH/BD/64057/2009), and A. Pereira (SFRH/BPD/26685/2006).
Disclosure Statement
No competing financial interests exist.