
Abstract
Introduction
In the last decade, technologies based on silver nanomaterials have been introduced into many science areas, including medicine, but the medicinal properties of silver were known much earlier. Silver was used in wound healing 27 because of its ability to promote epidermis synthesis by regulating zinc and iron metabolisms. 71 It was noted that nanocrystalline silver as used in wound dressings was an efficacious antibacterial agent and significantly promoted wound healing without compromising essential events in the wound-healing cascade. 77 Additionally, since silver nanoparticles (AgNPs) inhibit proliferation, migration, and the development of capillary-like structures, they may present antiangiogenic properties though activation of the PI3K/Akt-dependent pathway. 20
Silver has very high electrical and thermal conductivity, low variability, and is stable at high temperatures. 35 To be effective against microorganisms, it must have a small size to make direct contact with their cells. Silver ions show such an activity, but are also more toxic.15,17 The effectiveness of silver against bacteria increases when nanoparticles are used; then, their active surface, as well as their ability to act on microbes, increases millions of times. The ratio of the surface of nanoparticles to their mass is high, as is their ability to adsorb and transfer other compounds. Thermodynamic stabilization of nanoparticles is achieved by adding capping agents, which bind to the nanoparticles, prevent nanoparticle aggregation, and increase the solubility of the nanosystem. 65 The activity of AgNPs depends also on their shape: triangular particles are more reactive than, for example, spherical or cylindrical ones. 76
The high activity of nanoparticles against many microorganisms seems to be a promising alternative for antibiotics and disinfectants used in healthcare units. Interestingly, the activity of some antibiotics may be enhanced by the addition of AgNPs (ceftazidim, ampicillin, kanamycin, and erythromycin), but for the others (chloramfenicol), the results are not so clear.18,25 AgNPs demonstrate high antimicrobial and antifungal activity. It seems that most microorganisms cannot develop resistance, because silver binds to several targets in the cell, preventing the bacteria from replicating and expressing ribosomal proteins,15,48 as well as inactivating other enzymes. It is proposed that AgNPs act also on the bacterial cell membrane, producing pores and leading to cell death. 33 Both forms of silver also cause production of oxygen radicals and destroy cells.19,32,44 However, resistance to silver has been noted. 37
Some tests have shown that almost all viable cells (99.9%) of gram-positive bacteria, Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa were killed after 18 hours of exposure to the AgNPs. Almost total growth inhibition of Staphylococcus aureus (up to 99.27%) and E. coli (up to 98.44%) was recorded when the strains were cultured in the presence of nanofibers with silver-loaded zirconium phosphate. 12 Additionally, human dermal fibroblasts (attached to the nanofibers) continued to proliferate, suggesting that the fibers may be used as wound dressings. However, other studies present contrary results; neither silver ion release from nanoparticles nor nanoparticles themselves exhibited microcidal effects at dosing levels studied in vitro in tests against P. aeruginosa strains in the context of developing extended-wear or continuous-wear contact-lens formulations. 58
Such a rapid development of nanotechnology in recent years has increased the level of human exposure to nanomaterials. Therefore, an important issue is toxicity of AgNPs. 53 Occupational health studies have demonstrated that the greatest risk of silver absorption is related to chronic exposure to silver compounds, which may cause argyria and argyrosis.17,74 AgNPs have been shown to be cytotoxic and genotoxic to mammalian cell lines in vitro,4–6,16,23,24,32,50 to rats and Zebrafish in vivo,7,8,34 and to Vicia faba. 51 Clinical studies conducted with AgNP-impregnated wound dressings have shown prevention of infections in burn patients, 22 but clinical use of AgNPs (wound dressings) have also been associated with the development of hepatotoxicity and argyria-like symptoms. 73 The inhalation toxicity of AgNPs and oral toxicity tests indicated dose-dependent increases in mixed inflammatory cell infiltrate, chronic alveolar inflammation, and small granulomatous lesions related to AgNP exposure.34,69 It was shown that AgNPs induce cell necrosis or apoptosis in macrophages, 78 and decrease the viability of liver and neuron cells.25,26 However, on the contrary, Jena et al. report that chitosancoated (CS)-AgNPs exhibit potent antibacterial activity against some human pathogens (Mycobacterium smegmatis), but no damage to host cell DNA after treatment with a bactericidal dose of nanoparticles. 26 Additionally, toxicological tests of an AgNP gel in primary cultures and in a rat model, carried out in India, showed its safety for topical application. 25
Contrary to silver ions, AgNPs were seen to inhibit the growth of Candida spp. below concentrations that are toxic to human fibroblasts (30 mg/ml).6,49 Mei et al. indicate that 5-nm AgNPs are mutagenic in mouse lymphoma cells, and that mutation results from AgNP-induced oxidative stress. 44 Moreover, a significant decrease in mitochondrial function has been shown in germline stem cells (at a concentration of 10 μg/ml). 9 Some studies show no toxic effect on immortalized rat liver cells or human hepatocellular cell line (HepG2) up to 10 μg/ml 23 and 13 mg/ml 66 of AgNPs, respectively. In zebra fish embryos, AgNPs induced significant phenotypic changes starting from 25 μg/ml. 7
Mammalian cells seem to be able to phagocytose nanoparticles, and can subsequently degrade these particles by lysosomal fusion, 2 reducing toxicity and free-radical damage. This may account for the selectivity of AgNPs in promoting tissue-forming cell functions, while also inhibiting the bacterial functions that lead to infection. However, it was also found that exposure of cells in culture to AgNPs or Ag ions led to significant decreases in the new incorporation of selenium into selenoproteins, which may have significant implications in the defense against oxidative stress during long-term exposure. 68
The aim of our work was to evaluate the activity of AgNPs obtained by the physical method against standard, clinical, and environmental Enterococcus spp. strains, which showed diverse profiles of resistance against commonly used antibiotics and the presence of at least several virulence genes.
Materials and Methods
Enterococcus spp. strains (n=30) were isolated from the operative wards of one of the Teaching Hospitals of Lodz, Poland. Isolates were obtained from the urology ward (n=8), surgical ward (n=13), internal medicine ward (n=2), laryngology (n=6), and intensive care unit (n=1). Strains were isolated from patient samples (wound swabs n=7; urine n=4; throat swabs n=1; central venous catheters n=1), swabs from healthcare staff (hands, white coats, nose, throat n=4 strains), swabs from medical equipment and unanimated surfaces (n=13 strains). E. faecalis ATCC 29212 and E. faecium ATCC 35667 strains were used as controls. Standard and clinical strains were stored at −70°C for further investigations.
Enterococci were cultured according to the standard microbiological methods. Bacteria were identified by API 20 Strep tests (bioMerieux), and then identification was confirmed by PCR for
AgNPs (Axonnite, 50 ppm, 4 N; NanoTech Poland), obtained by microexplosion, were characterized by high purity (99.9999%). The AgNPs were suspended in demineralized water according to the Polish Pharmacopoea VI, 52 and Index Colour for Axonnite was CI 77820, CAS-7440-22-4. The other properties of Axonnite were pH—6.5, conductivity—5 to 50 μS, the concentration of AgNP—50 ppm±10%, density—0.990–1.010; turbidity max. 8 NTU, viscosity—1,000×10−6 Pascal seconds (Pa·s); color—max. 5 Pt/l, average particle size—2–5 nm (70%–75%) and 5–100 nm (25%–30%).
Susceptibility to AgNPs was tested by microdilution in 96-well microplates in Mueller Hinton Broth (bioMerieux). Thirty clinical and environmental strains and standard strain E. faecalis ATCC 29212 were investigated in three independent tests. Each isolate was tested in two repeats each time. To each dilution of AgNPs (from 25 to 0.048 μg/ml), 10 μl of 0.5 McFarland of bacteria was added. Positive and negative growth controls were used. Microplates were incubated for 24 hours in 37°C.
Susceptibility testing was carried out by the disc-diffusion method on Mueller-Hinton II Agar (bioMerieux) for the following antibiotics (Becton Dickinson): ampicillin (10 μg), ciprofloxacin (5 μg), erythromycin (15 μg), gentamicin (120 μg), penicillin (10 U), chloramfenicol (30 μg), streptomycin (300 μg), linezolid (30 μg), tetracyclin (30 μg), teicoplanin (30 μg), vancomycin (30 μg), and imipenem (10 μg). The results were interpreted according to the Clinical and Laboratory Standards Institute guidelines. 11 The Chi-squared test was used first, to show any correlation between ace and epa, agg, cylL/S, esp, gelE, and sprE, and secondly, to identify any significant relationship between resistance to antibiotics and the presence of virulence genes. The resistance to each antibiotic was categorized: the first group comprising sensitive strains, and the second group comprising both moderately sensitive and resistant strains. A p-value of 0.05 was used for statistical significance in these studies.
To determine the relationship between antibiotic resistance and minimal inhibitory concentrations (MICs) to AgNPs, the antibiotic resistance values were categorized. Two groups were tested: 1—strains sensitive to all drugs or resistant to one or two of them; 2—strains resistant to three or more tested drugs. MIC values were compared in both analyzed groups. Before further analysis, the results were tested for normality using the Shapiro–Wilk test. Because the results were significantly non-normal, the Mann–Whitney test was applied to the data. A p-value of 0.05 was used for statistical significance in all studies.
Specimens were then prepared for the scanning electron microscope (SEM). Standard strain E. faecalis ATCC 29212 was used to observe the activity of AgNPs on enterococcal cells. After overnight growth in dilutions of AgNPs (75% MIC), bacteria were centrifuged, and the resulting pellet was then fixed with 5% glutaraldehyde (in phosphate buffer, pH 7.4; Merck) for 2 hours. The pellet was washed four times in phosphate buffer (POCH) and taken through a graded alcohol dehydration series (50%, 60%, 70%, 80%, 90%, and 96%, in each for 10 minutes; POCH). Once dehydrated, the specimens were dried, mounted, coated with gold in a thermal-sputtering system (JEE-4 ×, JEOC) and viewed on the SEM (Hitachi S-3000 N), at a magnification of 6,000 ×. The images were analyzed with an Olympus CX 41 microscope and Cell B software (Olympus). Cells treated with AgNPs (Axonnite) were compared with control cells to show differences in the shape of enterococcal cells. The width and length of the enterococcal cells were measured. Before further analysis, the results were tested for normality using the Shapiro–Wilk test and for equality of variances (Levene test). Because the results were significantly normal, a statistical analysis was performed with the t-test. A p-value of 0.05 was used for statistical significance in all studies.
Results
Susceptibility to AgNPs and comparison to antibiotic susceptibility
The values of the MIC for Enterococcus spp. were between 0.39 and 3.125 μg/ml. The MIC was 1.56 μg/ml for the standard strain of E. faecalis ATCC 29212. The detailed results for each clinical and environmental strain are shown in Table 1. There were 12 strains with an MIC=3.125 μg/ml, 13 strains with an MIC=1.56 μg/ml, 3 strains with an MIC=0.78 μg/ml, and 2 strains with an MIC=0.39 μg/ml. The tested clinical strains of the Enterococcus spp. were sensitive to AgNPs at low concentrations.
MIC, minimal inhibitory concentration; AgNP, silver nanoparticle; AMP, ampicillin; C, chloramphenicol; CIP, ciprofloxacin; E, erythromycin; GM, gentamicin; IMP, imipenem; LNZ, linezolid; P, penicillin; S, streptomycin; TE, tetracycline; TEC, teicoplanin; VA, vancomycin.
Generally, 12 E. faecalis strains with an MIC=3.125 μg/ml seem more resistant to antibiotics than E. faecalis strains for which MIC=1.56 μg/ml. However, the statistical analysis revealed no relationship between the MIC of silver and resistance to antibiotics (p=0.26). Among 12 E. faecalis strains, 2 were resistant to ampicillin (16.66%), 6 to ciprofloxacin (50%), 8 to erythromycin (66.66%), 8 to gentamicin (66.66%), 5 to streptomycin (41.66%), and 9 to tetracycline (75%). For isolates for which colloid silver MIC=1.56 μg/ml, only one was resistant to ampicillin (10%), eight to ciprofloxacin (80%), two to erythromycin (20%), six to gentamicin (60%), two to streptomycin (20%), and seven to tetracycline (70%). All the isolates remained sensitive to both glycopeptides (teicoplanin and vancomycin) as well as linezolid. Only three strains (E. faecalis, E. faecium, and E. durans) were characterized by the MIC=0.78 μg/ml, and for one E. avium isolate, the MIC was 0.39 μg/ml. Additionally, the correlation between the presence of virulence genes and resistance to antibiotics was compared. No association was found besides the presence of the sprE gene and resistance to ciprofloxacin (p=0.047).
Susceptibility to AgNPs and presence of virulence genes
The presence of virulence genes in enterococcal strains with an MIC=3.125 μg/ml and an MIC=1.56 μg/ml was comparable. Among 12 E. faecalis isolates with an MIC=3.125 μg/ml, 8 strains had the agg gene, 9 cyl gene, 6 esp gene, 11 gelE sprE, and ace genes, and all of them possessed the epa and elrA genes. Eight of ten E. faecalis isolates with an MIC=1.56 μg/ml had the agg gene, three cyl gene, seven esp gene, and the other genes were found in ten isolates (Table 2). Isolates for which MIC=0.78 μg/ml had generally not so many virulence genes, and none possessed the esp or gelE gene. The E. avium strain had none of the investigated genes. Only one E. faecalis strain did not have the ace gene (0.33%; isolated from an environmental sample). Both epa genes were found in 28 enterococcal strains (93.33%). Strains without the genes (epa 5 and epa 6) were obtained from the environmental sample (n=1) and the wound (n=1). All enterococcal isolates possessed the elrA gene. Detailed results are presented in Table 2. The statistical analysis revealed significant correlation between the presence of the agg and gelE genes (p=0.018), the esp gene and the gelE (p=0.031), and the esp and sprE genes (p=0.017).
Analysis by scanning electron microscope
An analysis of the obtained pictures revealed some changes in the size of the enterococcal cells. When compared to an untreated control, cells grown with AgNPs were atypical. The analysis of cells treated with AgNPs in Axonnite performed with the t-test revealed that they were significantly wider than control cells (p=0.000087), but not significantly longer than control cells (p=0.609844). Figures 1 and 2 show an untreated control and enterococcal cells after using AgNPs, respectively.

Enterococcus faecalis ATCC 29212 cells. Control cells grown without AgNPs (Axonnite).
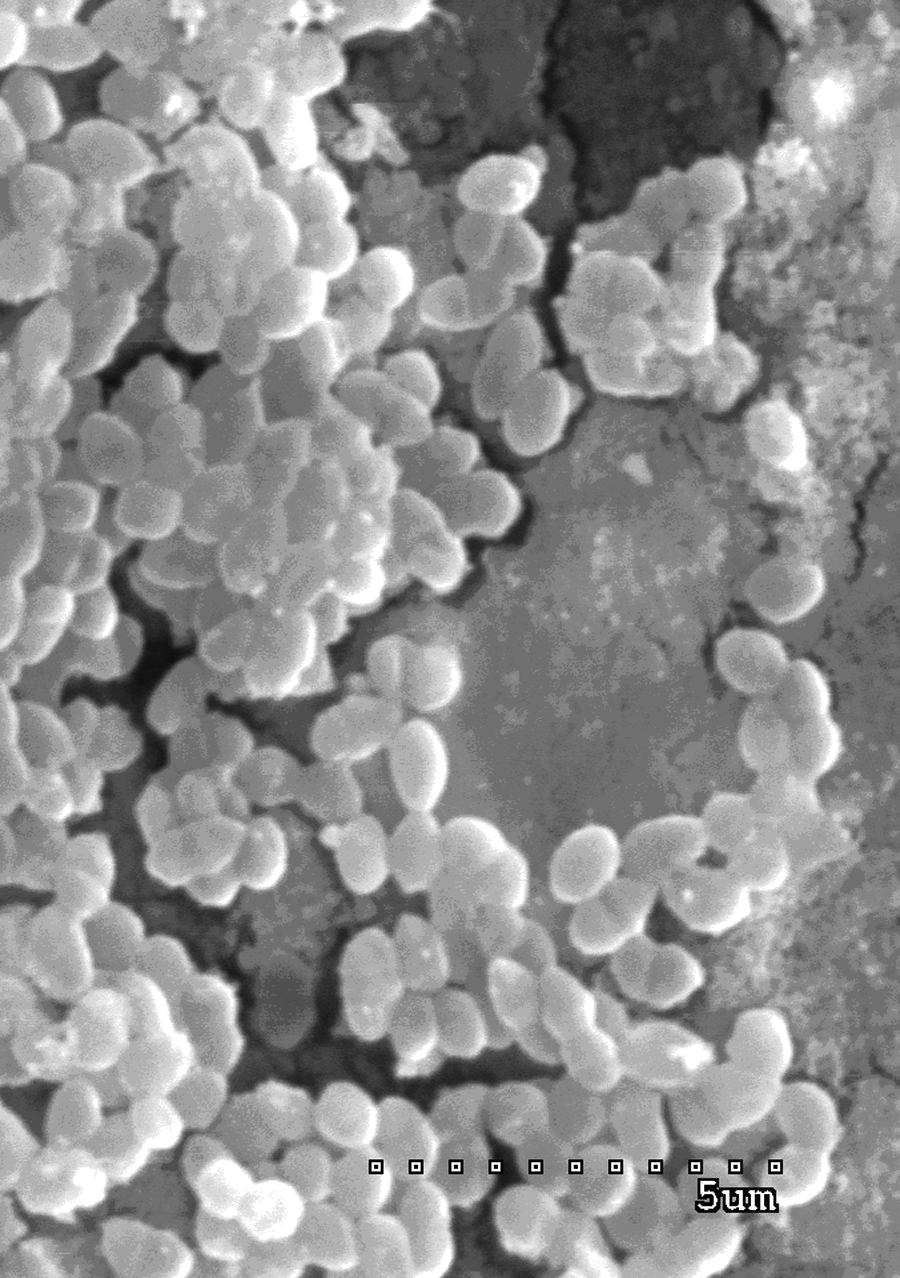
E. faecalis ATCC 29212 cells treated with 75% minimal inhibitory concentration of silver nanoparticles (Axonnite).
Discussion
The use of AgNPs for preventing infections has been under investigation for many years, but with recent advances in diverse techniques, silver is being more widely adopted in the medical community. Thanks to the multiple mechanisms by which silver acts on bacterial cells, AgNPs may be very effective against many antibiotic-resistant microorganisms, like enterococci. The features of clinical enterococcal strains are the presence of many virulence genes and resistance to several antibiotics.1,13,60 It has been shown that clinical strains of E. faecalis are more likely to present virulence genes than E. faecium strains.1,13,40,60 Contrary, E. faecium strains are usually more resistant to antibiotics.13,38 Thanks to these traits, it is important to look for new possibilities for the treatment and prevention of enterococcal infections.
One of the new approaches to preventing such infections may be the use of AgNPs. Contrary to ionic forms, nanoparticles may be obtained by physical methods, such as Axonnite, and are therefore not as chemically contaminated as ion forms. The other way of obtaining AgNPs is its production by some species of lactic acid bacteria (e.g., Lactobacillus spp. and Pediococcus sp.). Lactobacillus fermentum produce small particles (a mean size of 11.2 nm, a purity of 70%). 67 It was shown that those AgNPs inhibited the growth of E. coli, S. typhimurium, E. faecalis, and C. albicans. 56 However, the stability and purity of those nanoparticles seem to be lower than those found in Axonnite (purity 99.9999%).
It has been reported that gram-positive bacteria were less susceptible to silver than gram-negative ones.29,33 In our study, all of the investigated enterococcal strains were inhibited at MIC ranging from 0.39 to 3.125 μg/ml, including all the multidrug-resistant strains possessing multiple virulence genes. Sadhasivam et al. found that the biosynthesized AgNPs displayed a pronounced antimicrobial activity against different pathogenic microorganisms. The mean of the diameter of the zone of inhibition for E. faecalis and E. coli was 14.2±0.46 and 17.2±0.26, respectively. 56 Staphylococcus spp. MIC was 4 μg/ml. 59 The others tested AgNPs against several types of bacteria. It was shown that some (7–20 nm) were effective at MIC 0.78–6.25 μg/ml against S. mutans strains. 21 Inhibition of bacterial growth was obtained after 10.5 hours (P. aeruginosa) or after 1.3 hours (Staphylococcus spp.). 25 In all tests, AgNPs were more effective than other compounds investigated, and possessed strong biocidal effects on multiresistant bacteria such as methicillin-resistant S. aureus or multidrug-resistant E. coli.55,61 Also, AgNPs in Axonnite were very active when compared to antibiotics tested. Only glycopeptides and linezolid, last-line drugs against enterococci, were active against all strains investigated. However, according to other data, strains resistant to vancomycin, teicoplanin, and linezolid have appeared worldwide.46,47,54,57 Nosocomial infections caused by such resistant strains have been especially dangerous in older patients with multiple comorbidities: their occurrence has been associated with a significantly longer ICU stay and a trend toward higher a mortality rate. 45 Therefore, preparations such as Axonnite may well be a valuable weapon against those multidrug-resistant bacteria, but further studies on its toxicity are required.
There was no statistically significant relationship between resistance to antibiotics and MIC of AgNPs (Axonnite). The lack of association may be the result of small sample size; hence, further studies should be conducted. It is difficult to make a correlation between antibiotic resistance and the MIC of AgNPs, as other studies have focused on investigating single strains from one species.15,21,49,55,56 In our study, six clinical E. faecalis strains, which were susceptible to AgNPs at 1.56 μg/ml, were comparably resistant to six E. faecalis strains with AgNPs MIC 3.125 μg/ml.
Environmental strains of E. faecalis with MIC=1.56 μg/ml seem to be more sensitive to antibiotics than those with MIC=3.125 μg/ml, that is, resistant to one to four antibiotics tested versus three to eight antibiotics in the group with MIC=3.125 μg/ml. E. avium and E. durans were inhibited at low concentrations of AgNPs (0.39 and 0.78 μg/ml, respectively). Surprisingly, E. faecium isolates, usually more resistant to antibiotics,13,38 were inhibited at lower AgNP concentrations (0.78 and 1.56 μg/ml).
According to the literature, clinical enterococcal strains that possess multiple virulence traits are more dangerous. It is proposed that some virulence factors, like an aggregation substance and cytolisin, may occur altogether and be transferred between Enterococcus spp. strains.60,75 In our study, the agg genes were found in 18 of 30 Enterococcus spp. strains (60.0%). The agg gene was present in eight strains with MIC=3.125 μg/ml and in eight strains with MIC=1.56 μg/ml. Differences were shown for cytolisin genes. Cytolisin genes were found in eight strains for which MIC=3.125 μg/ml and only three strains with MIC=1.56 μg/ml. The other genes, for Esp protein, are typical to hospital clonal strains and were found with almost 100% frequency. 72 In the present study, the gelatinase gene, gelE, was found in 21 enterococcal strains (70.0%). The number of strains that had the gelE gene and were characterized by MIC=3.125 μg/ml or MIC=1.56 μg/ml were comparable: 10 and 11, respectively. Arciola et al. discovered a high frequency of the gel gene among both clinical and epidemic enterococcal strains. 3 The presence of that gene among investigated strains indicates their virulence, but on the other hand, the MIC of AgNP in Axonnite supports its high activity. The next investigated virulence gene was the serine protease gene, 63 also situated in the fsr locus. 30 In the present study, the majority of strains had the sprE gene, both with MIC=3.125 μg/ml and MIC=1.56 μg/ml. The results of the statistical analysis revealed that there was significant association between the agg and gel genes (p=0.018), the esp gene and gelE genes (p=0.031), or the esp and sprE (p=0.017) genes. It may be due to their common transfer among enterococcal strains, but further studies should be conducted to confirm this result.
The presence of some genes seems to be necessary in the pathogenesis of enterococcal infections.10,36,70 For example, it was shown that strains of E. faecalis with the ace gene were often isolated from urinary tract infections.1,41 In the present study, we found the ace gene in the majority of tested strains of E. faecalis (n=29; 96.66%). Another important group of virulence factors are Epa polysaccharides, 70 which play an important role in the initial attachment and biofilm accumulation of E. faecalis OG1RF. 64 In our study, we checked the frequency of two genes from the epa cluster (epa5 and epa6) among 30 strains of E. faecalis. Our results confirm the common presence of the epa genes among enterococci. Malathum et al. showed that the orfde4 to orfde10 epa genes were in all of the 12 genotypically and geographically diverse strains of E. faecalis investigated in their study. 42 In our work, the epa genes (at least one) were found in 28 enterococcal strains (93.33%). Our results show that almost all strains had two of the investigated genes. Those results seem to support the theory that the epa genes are required during infection. The final tested gene, elrA, is widespread among E. faecalis strains, and ElrA may facilitate the evasion of the early bactericidal activities of macrophages. 10 In our studies, the elrA gene was found in all strains investigated, which implies that the gene is necessary during enterococcal pathogenesis. After a comparison of the presence of virulence traits in tested enterococci with the MICs for AgNPs, we may conclude that there are no significant differences between strains susceptible to higher and lower concentrations of AgNPs. Such results can be due to a small sample size. All tested strains remained highly sensitive to nanoparticles, irrespective of the presence of virulence genes.
To our knowledge, there are no studies demonstrating the susceptibility to AgNPs of clinical and environmental strains of Enterococcus spp. According to Egger et al., standard strains of gram-positive bacteria were more tolerant to silver than gram-negative cells, except for E. faecalis ATCC 19433, for which the MICs and minimal bactericidal concentrations (MBCs) were similar to those for gram-negative bacteria. 14 The MICs of silver particles for other species, such as E. coli, ranged from 1.9 to 75 μg/ml.14,59 Our results for all strains tested show that enterococci are highly susceptible to AgNPs in Axonnite. Moreover, enterococci remain sensitive to AgNPs regardless of being multiresistant. It confirms that AgNPs may be very active in very small concentrations against those very common multiresistant strains.
Research focusing on the development of a AgNP-containing coating would be more attractive for future infection prevention, since AgNPs act against a wide range of pathogens in minor concentrations and with a low risk of resistance development.14,15,37 It is especially important in case of enterococci, which are able to acquire resistance to multiple antibiotics. Our data suggest that even multidrug-resistant enterococcal strains are highly sensitive to AgNPs in Axonnite. Moreover, there seems to be no correlation between resistance to antibiotics and concentration of nanoparticles active.
To summarize, diverse antibacterial agents containing silver were developed to reduce bacterial adhesion and prevent biofilm production. Silver is effective against many bacterial species, but its concentration is crucial to avoid cytotoxity or genotoxicity. The AgNPs in Axonnite (NanoTech) seem to be a promising alternative for other drugs to prevent infections caused by multidrug-resistant, virulent enterococcal strains because of its low effective concentrations and low possibility for resistance development. However, there is still a need to conduct studies to learn about their effectiveness and toxicity in in-vitro and in vivo models, as other AgNPs have adverse effects. As the future application of Axonnite could be limited by the fact that it may be toxic to human cells, special attention should be paid to all aspects associated with its role in treating illness, not just its antimicrobial potential.
Footnotes
Acknowledgments
The authors wish to thank Paweł Siejko (NanoTech, Poland) for providing silver nanoparticles and for valuable advice. This work was supported by the Medical University of Lodz (grant no. 503/5-020-03/503-01).
Disclosure Statement
No competing interests are present.