
Abstract
To assess the clonal structure of Staphylococcus aureus in the United States, we performed a molecular epidemiological study of 1,055 S. aureus isolates from a nationally representative clinical isolate collection from 2004–2008. Resistant and susceptible isolates were typed with multilocus sequence typing, tested for the presence of Panton-Valentine leukocidin (PVL), and serotyped. USA300 (multilocus sequence typing clonal complex 8, PVL positive, and methicillin-resistant) was the most frequently isolated clone, expanding from 12% of all isolates in 2004 to 38% in 2006. The USA300 clone increased significantly in frequency among both outpatients and inpatients. USA300 increased in both skin and soft-tissue and invasive infection isolates. The second most frequently observed clone was clonal complex 5, PVL-negative, and methicillin-resistant, and its frequency was stable from 2004–2008. The methicillin-susceptible S. aureus in the study was polyclonal, and decreased in frequency as it was replaced by USA300.
Introduction
The rise of the USA300 clone and other changes within the S. aureus population have been documented in various accounts. Nevertheless, the overall clonal structure of S. aureus in the United States and the expansion of USA300 have not been adequately analyzed in a statistically well-powered study covering all nine census regions, a range of ages, specimen sources, and patient settings. To address this, we designed a molecular epidemiologic study of S. aureus in the United States from 2004–2008.
Materials and Methods
Clinical isolates
We obtained 1,066 S. aureus isolates from Eurofins Medinet (Table 1). This isolate collection is the largest in the United States and comes from over 200 clinical laboratories distributed among all nine U.S. Census Bureau Regions across the country. Participating laboratories submit single patient, nonconsecutive, and nonduplicate isolates as part of various surveillance initiatives. These surveillance initiatives were designed to capture the staphylococcal phenotypes in proportions that reflect the distributions seen within the larger database of The Surveillance Network. Eurofins collects clinical infection isolates from both inpatients and outpatients, and includes isolates from a variety of specimen sources and diverse patient demographics. When we commenced this study in 2009, the collection consisted of 14,151 clinical infection isolates from the years 2004, 2006, and 2008. We randomly sampled this collection without replacement to generate our study sample. Of the 1,066 isolates sampled, we were able to complete molecular characterization on 1,055, and we report the results of those isolates.
Setting information was missing for 17 isolates.
SSTI, skin and soft-tissue infection.
Isolates were identified as MRSA or MSSA on the basis of testing for susceptibility to oxacillin (which is used as a proxy for β-lactam antimicrobial drugs, including methicillin) and classified as susceptible or resistant according to National Committee for Clinical Laboratory Standards (NCCLS) breakpoint criteria. Eurofins did not provide diagnoses, but did provide the site of specimen collection (e.g., blood or urine). To simplify analysis, certain specimen sites were grouped as follows: SSTI (skin, SSTI, and wound), Blood and Lower Respiratory (Blood, Bronchial Washing, Sputum, and Tracheal Aspirate), and Other (Conjunctiva, Corneal scrapings, Ear, Other, Unknown, Upper Respiratory, and Urine). Eurofins also provided information on the geographic location where the isolates were collected, the year of isolation, the patient's age, and the patient's location at the time of sample collection. Inpatient isolates were obtained in hospital wards or intensive care units, and outpatient isolates were obtained in clinics or emergency rooms.
Molecular characterization
MLST and PVL testing were performed as previously described. 11 Isolates were serotyped using an enzyme-linked immunosorbent assay (ELISA) using conjugate rabbit antibodies specific to capsular polysaccharide (CP)5 and CP8. Microtiter plates were coated overnight at 4°C in polyclonal antibodies against either CP5 or CP8 diluted 1:500 in phosphate-buffered saline (PBS). Plates were then washed and blocked with PBS–bovine serum albumin 1%. Samples were added in the form of a suspension of killed whole cells. Positive controls consisted of suspensions of killed whole cells of strain Wright (CP8) or strain Lowenstein (CP5); purified CP5 and CP8 were also used as positive controls. Plain PBS and suspensions of killed whole cells of USA300 strain FPR3757 were used as negative controls. Samples were then incubated with mouse monoclonal antibodies against CP5 (diluted 1:5,000 in PBS) or CP8 (diluted 1:8,000 in PBS). Goat anti-mouse peroxidase was used as a secondary antibody, and coloration was carried out using SIGMAFAST OPD tablets (Sigma-Aldrich).
To differentiate weakly positive results from background, we calculated statistical cutoffs based on the standard deviation of negative control readings using within-plate variation and experiment-wide variation. To avoid overestimation of the frequency of nontypeable isolates, statistical cutoffs were calculated to produce an estimated 1% false-positive rate.
To confirm our serotyping results, we sequenced a fragment of the cap5D gene that contains a mutation that produces a nontypeable phenotype. 3 We amplified and sequenced this region using the following primers: cap5dF (5′-actaggcagagatcctgttg-3′) and cap5dR (5′-gcacaaatacttcgccacc-3′).
Statistical methods
Sampling weights were included in all univariate and multivariate analysis to reflect the appropriate distribution of the sample according to the US population. Weighted multivariate logistic regression was used to analyze the data and compute point estimates and 95% confidence intervals. STATA® 11 was used for all statistical analysis.
Results
Clonal structure of S. aureus in the United States, 2004–2008
We found that three categories of S. aureus accounted for nearly 90% of all sampled isolates across the three timepoints, 2004, 2006, and 2008: the CC8, PVL-positive MRSA (USA300) clone, the CC5 PVL-negative MRSA clone, and polyclonal PVL-negative MSSA (See Fig. 1). USA300 was the most frequently observed clone (N=325, 31%±3%). Among PVL-positive CC8 MRSA, 95% were sequence type 8 (the remainder were rare and/or novel variants). The second most frequently observed clone was PVL-negative MRSA from CC5 (N=296, 28%±3%), representing the major traditional HA-MRSA in our sample.11,19 Among PVL-negative CC5 MRSA, 70% were sequence type 5 (14% were sequence type 105, 2% were sequence type 225, and the remainder were rare and/or novel variants). The PVL-negative MSSA observed in our study (N=314, 30%±3%) was polyclonal. The MSSA population was made up of 19 different CCs, four of which (CC1, CC5, CC8, and CC30) accounted for 72% of MSSA isolates. These four CCs were also the most prevalent MRSA CCs. The isolates falling outside the three dominant categories were non-CC8 PVL-positive MRSA clones (1%), non-CC5 PVL-negative MRSA clones (6%), and PVL-positive MSSA clones (4%).
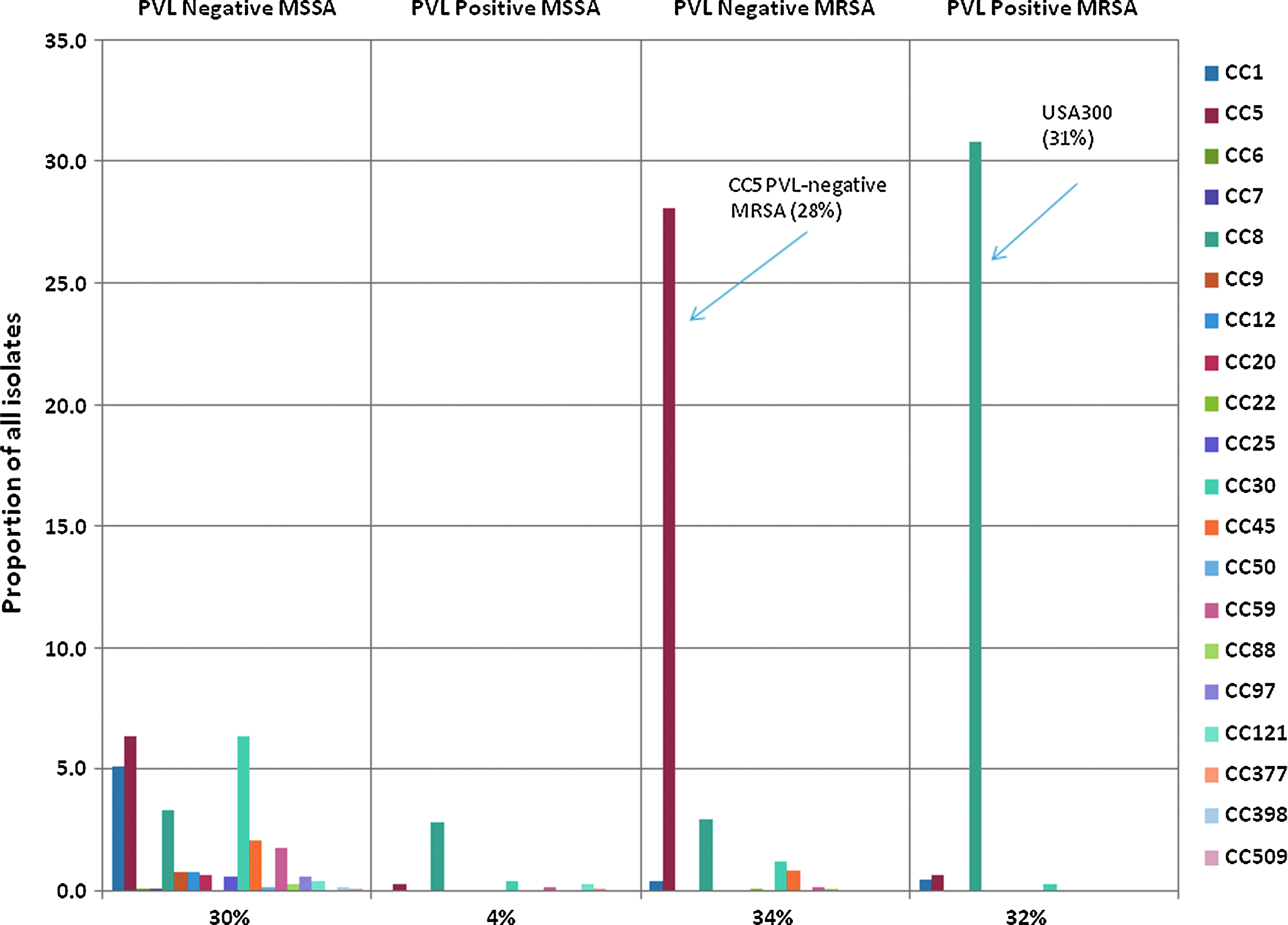
Overall frequency distribution of Staphylococcus aureus clonal complexes (CCs) by Panton-Valentine leukocidin (PVL) and methicillin-resistance status. Along the X-axis, CCs are divided into four groups on the basis of presence of the PVL genes and methicillin-resistance status. The percentage of isolates comprised by each of the four categories is listed below the X-axis. Within each category, each bar corresponds to a CC, with CCs ordered from CC1 up to CC509 from left to right. The two major clones, CC5 PVL-negative methicillin-resistant S. aureus (MRSA) and USA300 (CC8 PVL-positive MRSA), are highlighted along with the percentage of all isolates composed of that clone. Color images available online at www.liebertpub.com/mdr
Changes in clonal structure overall and by patient setting
Between 2004 and 2006, the USA300 clone significantly increased in frequency (Fig. 2a) in the United States. The increase was significant in both inpatient and outpatient isolates (Fig. 2b, c). By 2008, the frequency of USA300 among inpatients was close to that of the traditional HA-MRSA strain (27%±6.6% for USA300 vs. 33%±3.4% for CC5 PVL-negative MRSA). The increase in USA300 from 2004–2006 was accompanied by a decrease in PVL-negative MSSA (Fig. 2a). PVL-negative MSSA accounted for 44% of all isolates in 2004 and by 2006 declined to 20% (p<0.001). In 2008, the frequency of PVL-negative MSSA rebounded to 31% (p<0.001), accompanied by a slight decline in all MRSA (p<0.001).
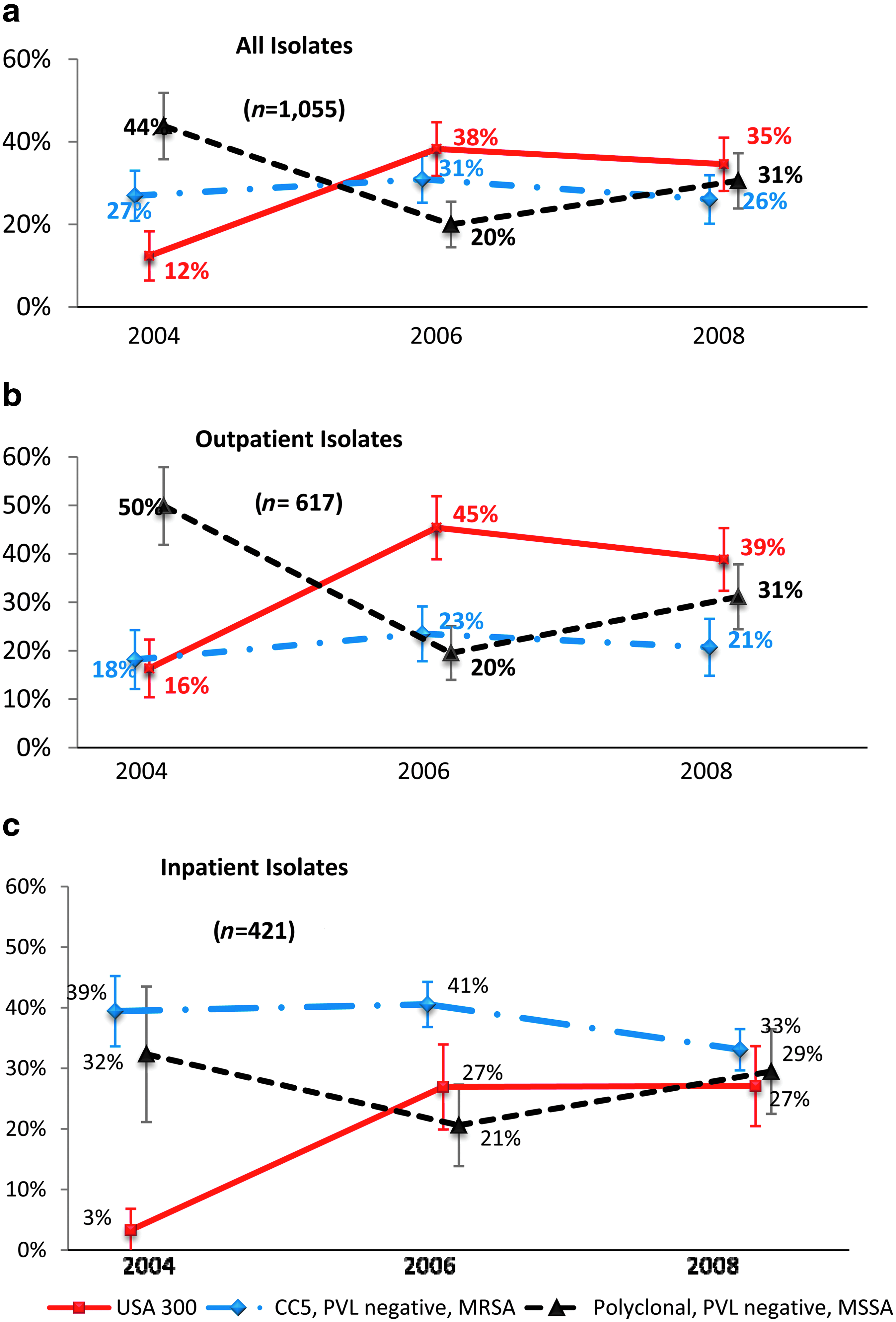
Proportion of all isolates that were USA300 (CC8 PVL-positive MRSA), CC5 PVL-negative MRSA, or Polyclonal PVL-negative methicillin-susceptible S. aureus (MSSA) over time.
In contrast to the changes observed for USA300 and polyclonal MSSA, CC5 PVL-negative MRSA had no significant changes in frequency from 2004–2008. The proportion of MRSA that was USA300 doubled from 23% in 2004 to half of all MRSA isolates by 2006 (51%, p<0.001) with the same pattern of stabilization in 2008 (no significant change, 53% of MRSA isolates).
Changes in the proportion of USA300 by specimen source, age, and geography
The rise of the USA300 clone between 2004 and 2006 was most significant among children under 18 (Fig. 3a). USA300 increased in frequency among SSTI isolates and in isolates commonly associated with invasive disease (blood or lower respiratory tract) (Fig. 3b). The expansion of USA300 occurred nationwide, with gains in all US census regions except the East-South-Central/West-South-Central region, where the frequency of USA300 in 2004 was already high at 36% (Fig. 3c). In 2004, some regions, such as the Northeast had very low frequencies of USA300, but by 2008 all census regions had effectively the same frequency of USA300 (overlapping confidence intervals).

USA300 as a proportion of all S. aureus isolates over time.
Serotypes and clones
Overall, 36% of isolates in the study expressed CP5, 19% expressed CP8, and nearly half (46%) were nontypeable for CP5 or CP8 by ELISA (Table 2). Serotypes were correlated with clones and the presence or absence of PVL.
A small fraction of serotyping results were ambiguous and were excluded from this table.
Nontypeable for CP5 or CP8.
CC, clonal complex; CP, capsular polysaccharide; MSSA, methicillin-susceptible S. aureus; MRSA, methicillin-resistant S. aureus; PVL, Panton-Valentine Leukocidin.
To verify the nontypeable phenotype of USA300, we sequenced a fragment of the cap5D gene from 33 nontypeable USA300 isolates to test for the presence of an insertion mutation that prevents expression of capsule. 3 All 33 isolates had the insertion mutation that disrupts the reading frame of cap5D. In addition, we found the mutation among all USA300 isolates that expressed capsule type 5 or 8 according to the ELISA test, indicating that these may have been false positives.
Discussion
This study reveals a population structure for S. aureus in the United States in which CA-MRSA, HA-MRSA, and MSSA are present in roughly equal proportions, the CA-MRSA and HA-MRSA are each dominated by a single clone, and the CA-MRSA clone (USA300) and the HA-MRSA clone (CC5 PVL-negative MRSA) are distinct. We found that MRSA tended to be distributed in a few high frequency clones, while MSSA was highly polyclonal. This finding is consistent with other studies, which noted that MRSA clones first emerged only 60 years ago, while MSSA clones are more ancient,2,12 and perhaps only a subset of clonal lineages have a suitable genetic background for stable carriage of SCCmec.6,14
One of our most important findings is that USA300 increased significantly among inpatients. While our study did not distinguish between hospital-acquired inpatient isolates and hospital-onset inpatient isolates, it is notable that by 2008, USA300 was responsible for nearly as many infections among inpatients as was the traditional HA-MRSA clone. This observation is consistent with other studies that found USA300 in the hospital setting.15,24
Our data on S. aureus capsule expression differs from some previous estimates, but is consistent with recent studies of CA-MRSA in the Unites States. CP5 and CP8 are the only capsule types common in clinical isolates, and nontypeable strains were estimated to account for ∼15%–20%.20,23 The nontypeable phenotype of USA300 has been reported before,21,26 and our results confirm that at least 96% of USA300 isolates are nontypeable, and that this phenotype is linked to a single mutation in the cap5D locus. We suspect that the percentage of USA300 isolates that are unencapsulated is actually 100%, but that our assay produced a small number of false positives. This is consistent with our finding that every USA300 isolate we tested carried a mutated version of cap5D. Our results are also consistent with those of Sutter et al., 26 revealing that, since the emergence of CA-MRSA and USA300 in the Unites States, the frequency of nontypeable S. aureus in the United States is significantly higher than previously believed. We noted a small number of isolates that reacted weakly to antibodies against both CP5 and CP8, but these are probably artifacts, so the results are not shown. When choosing statistical cutoffs to differentiate weak capsule-expressing isolates from background noise in our ELISA method, we did not want to overestimate the number of nontypeable isolates, so we were more likely to report false positives than false negatives. The literature on S. aureus microcapsule expression does not support the existence of isolates that express both CP5 and CP8, and they are in fact considered allelic.3,20,22,23
It is interesting that capsule expression and PVL are negatively correlated, because both capsule and PVL are involved in avoidance of phagocytic killing by human polymorphonuclear leukocytes.7,10,22 This observation suggests that PVL's role in virulence may be more important in strains, such as USA300, that lack capsule. This is consistent with results showing that S. aureus cytotoxicity derives from multiple, partially redundant virulence factors. 18
In summary, MRSA infections in the United States are predominantly caused by two major clones, while MSSA is polyclonal. The USA300 clone of MRSA has increased significantly nationwide, and has increased in locations and infections previously associated with HA-MRSA. The rise of USA300 may be attributable to its unique characteristics, including its lack of CP5 or CP8 and the presence of PVL.
Footnotes
Acknowledgments
We gratefully acknowledge the GlaxoSmithKline United States sequencing facility under the management of Ganesh Sathe, especially Elizabeth Thomas and Stephanie Van Horn. We thank Stuart Burstin for constructive feedback on earlier versions of the article, and Linda Miller, Linda Mundy, Nicole Scangarella-Oman, Emily Lu, and Cosmina Hogea for stimulating discussions. We also thank Daniel Sahm and Eurofins Medinet for providing the isolates for this study.
Disclosure Statement
This work was funded by GlaxoSmithKline and all authors report no other conflicts.