
Abstract
We report the emergence of a multidrug-resistant Haemophilus influenzae strain in a patient with common variable immunodeficiency suffering from recurrent bronchopneumonia caused by H. influenzae. After the patient had received several antibiotic therapies, a strain was isolated showing resistance to ampicillin, ampicillin/sulbactam, cefazolin, cefuroxime, ciprofloxacin, and clarithromycin. Polymerase chain reaction analyses and sequencing revealed the presence of the beta-lactamase gene blaTEM-1, two mutations (A502T and R517H) in the ftsI gene encoding the transpeptidase region of the penicillin-binding protein 3, and one mutation in the ribosomal protein gene L4 (G65D) conferring resistance to beta-lactams and macrolides, respectively. Additionally, the plasmid-encoded aac(6′)-Ib-cr gene mediating slightly reduced susceptibility to quinolones and two mutations in the DNA gyrase gene gyrA and one mutation in the topoisomerase IV gene parC were identified leading to a high-level fluoroquinolone-resistant phenotype. In conclusion, the treatment of H. influenzae infections accompanied by high bacterial loads such as bronchopneumonia can be complicated by the selection of multidrug-resistant strains. Moreover, the emergence of aac(6′)-Ib-cr in H. influenzae causing low fluoroquinolone resistance levels might have contributed to the selection of DNA gyrase and topoisomerase IV mutants.
Introduction
Resistance to the beta-lactam antibiotic ampicillin in H. influenzae is primarily mediated by the production of beta-lactamases, for example, TEM-1 or ROB-1-type enzymes.9,25 An international surveillance study analyzing H. influenzae isolated from patients with community-acquired respiratory tract infection showed an overall prevalence of beta-lactamases in 15.0% and a beta-lactamase-prevalence in Germany of 6.0% of the strains. 7 H. influenzae isolates with reduced susceptibility to fluoroquinolones are very rare 2 and are usually associated with mutations in the quinolone resistance-determining region of the genes encoding DNA gyrase or topoisomerase IV (gyrA and parC, respectively). 8 Additionally, in Haemophilus parasuis, the plasmid-encoded genes qnr and aac(6′)-Ib-cr mediating reduced susceptibility to quinolones have been described. 11
The present study describes in detail the multidrug-resistance mechanisms of a H. influenzae strain causing bronchopneumonia in a patient with CVID.
Case Report
A 34-year-old female patient was repeatedly admitted to our hospital since 2007 with cough and production of purulent sputum. Physical examination and imaging of the chest revealed recurrent bacterial bronchopulmonal infections and splenomegaly. Laboratory testing results showed leukocytosis, neutrophilia, and elevated C-reactive protein each time of admission. As an underlying disease, the CVID syndrome with severe hypo-gamma-globulinemia affecting all classes of immunoglobulins (IgA, IgG, and IgM) was initially diagnosed in 2001. Parenteral immunoglobulin substitution therapy was started in 1998 and continued since March 2011 by a subcutaneous delivery system. Since 2010, the patient suffered from chronic organizing pneumonia with several episodes of exacerbation showing peribronchial and bronchiolitic shadowing with beginning abscess formation by chest computed tomography consistent to bacterial infiltrates (Fig. 1). Between March 2010 and May 2011, H. influenzae was repeatedly isolated from respiratory materials, that is, sputum, tracheal secretion, bronchoalveolar lavage, and lung biopsy. H. influenzae showing antimicrobial susceptibility to all antibiotics tested was detected for the first time in March 2010. Subsequently, H. influenzae isolates with increasing antimicrobial resistance emerged (Fig. 2). In September 2010, H. influenzae isolates showing resistance to ampicillin and ciprofloxacin were recovered from tracheal secretion, bronchoalveolar lavage, and lung biopsy. In May 2011, two isolates resistant to ampicillin, ampicillin/sulbactam, cefazolin, cefuroxime, ciprofloxacin, and clarithromycin emerged. Between 2007 and 2011, the patient was treated with several anti-infectives, including fluoroquinolones, macrolides, and beta-lactam antibiotics (Fig. 2).
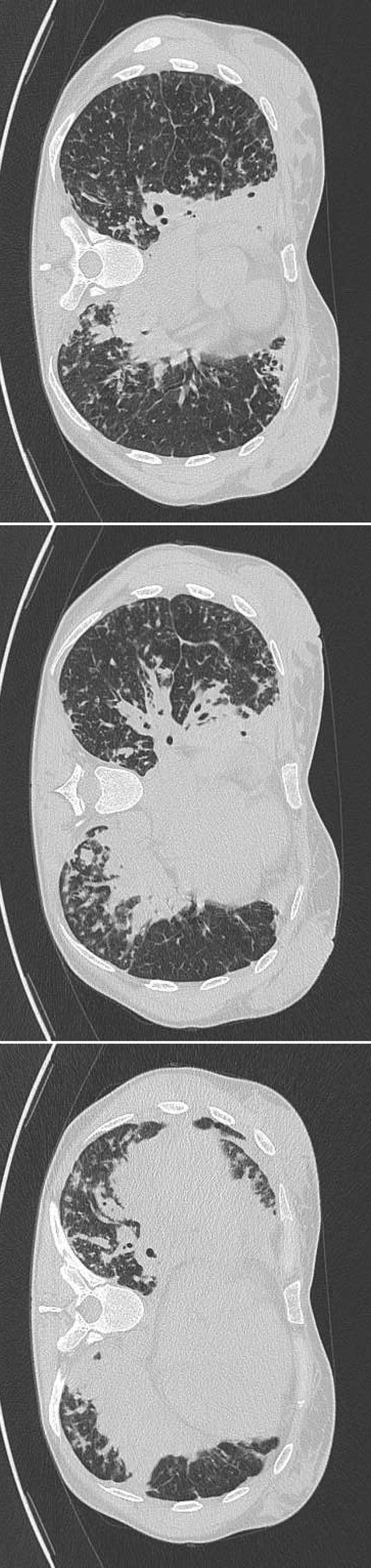
Chest computed tomography of the patient in March 2010.
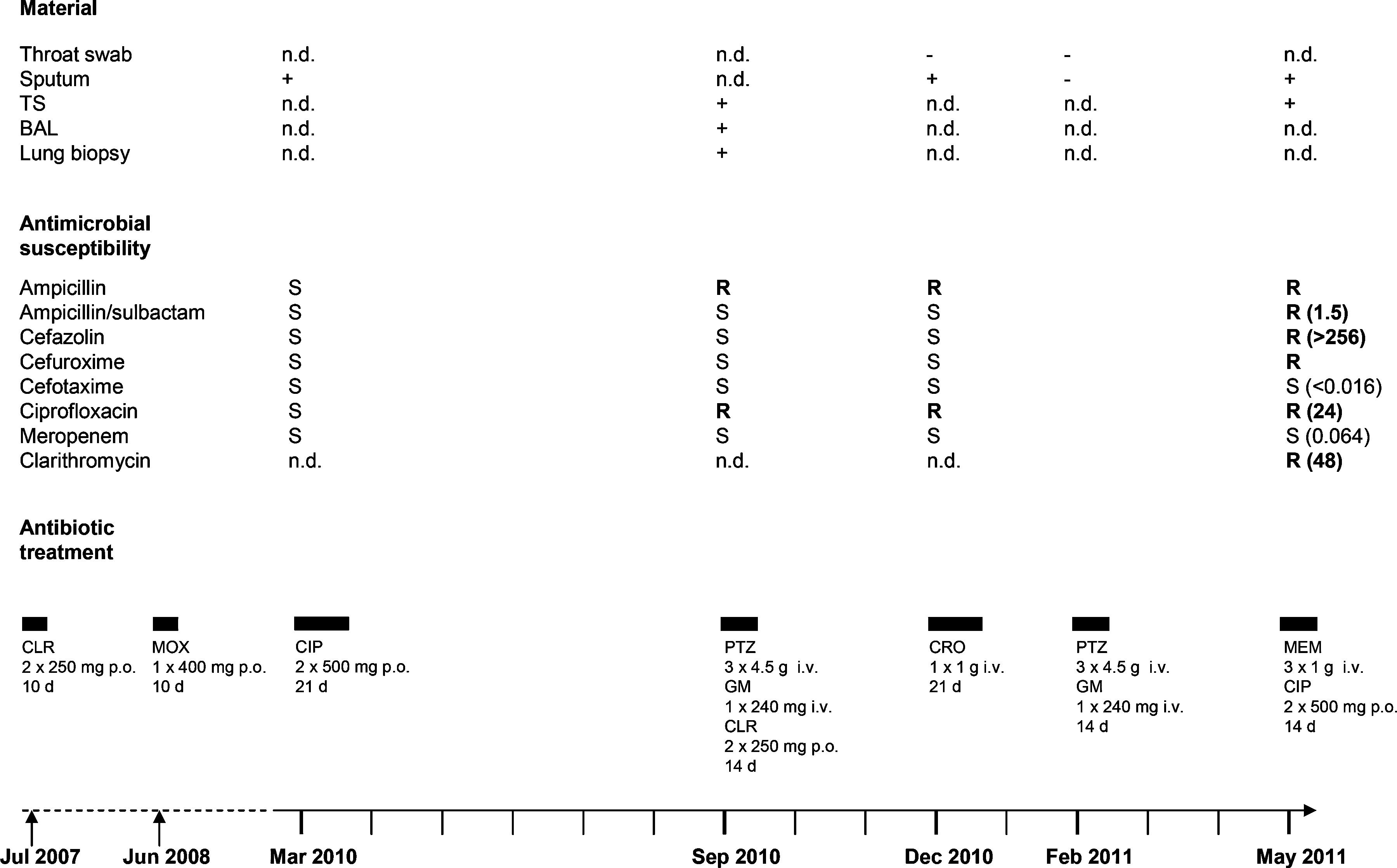
Haemophilus influenzae isolated from the patient between March 2010 and May 2011 and antimicrobial therapies 2007–2011. Minimal inhibitory concentration (MIC) values [μg/ml] determined by Etest are given in brackets. TS, tracheal secretion; BAL, bronchoalveolar lavage; n.d., not determined; +, culture positive; −, culture negative; AP, ampicillin; AUG, ampicillin/sulbactam; CZ, cefazolin; CXM, cefuroxime; CTX, cefotaxime; CIP, ciprofloxacin; MEM, meropenem; CLR, clarithromycin; MOX, moxifloxacin; CRO, ceftriaxone; PTZ, piperacillin/tazobactam; GM, gentamicin; S, susceptible; R, resistant; i.v., intravenously; p.o., per os.
Materials and Methods
Bacterial cultivation, identification, and antimicrobial susceptibility testing
Throat swabs were cultivated on Columbia blood agar (Oxoid, Wesel, Germany) for 24 hr at 37°C under aerobic conditions using streak inoculation with a bovine, beta-hemolytic Staphylococcus aureus strain to enhance the ability to recover Haemophilus spp. and placing of an optochine-containing disc (Oxoid) to detect S. pneumoniae. Additionally, throat swabs were cultivated on Endo agar (Becton Dickinson, Heidelberg, Germany) containing 0.04% carbol fuchsin solution (Applichem, Darmstadt, Germany) for 24 hr at 37°C under aerobic conditions for the detection of gram-negative organisms.
Sputum, tracheal secretion, and bronchoalveolar lavage were each cultivated on Columbia blood and Endo agar for 48 hr as described above. Additionally, these materials were cultivated on brain–heart infusion (BHI) agar (Oxoid) + 5% sheep blood + IsoVitaleX enrichment (Becton Dickinson; own production) in an anaerobic jar using a sachet absorbing atmospheric oxygen with a methylene blue indicator strip for 48 hr at 37°C and using a bacitracin-containing disc (Becton Dickinson) to enhance the ability to recover Haemophilus spp. The lung biopsy was cultivated in a tryptic soy broth (Oxoid) containing beef liver for 48 hr at 37°C under aerobic conditions. For subcultivation, Columbia blood and Endo agar (each incubated under aerobic conditions, 48 hr, 37°C) and BHI agar + 5% sheep blood + IsoVitaleX enrichment (incubated as described above) were used.
H. influenzae was identified by growth in small satellite colonies around the S. aureus streak, resistance to bacitracin, and growth dependency on hemin (factor X) and nicotinamide adenine dinucleotide (NAD+, factor V). The latter was tested on tryptic soy agar (Oxoid) incubated under aerobic conditions for 24 hr using discs containing factor V and factor V + X, respectively (Oxoid). M. catarrhalis was identified by positive oxidase and DNase reaction. Identification of the isolates was confirmed by MALDI-TOF MS analysis performed with an AXIMA Assurance instrument (Shimadzu, Duisburg, Germany) using alpha-cyano-4-hydroxy cinnamic acid (AnagnosTec, Potsdam, Germany) as a matrix and Launchpad 2.8.4 and SARAMIS premium software (AnagnosTec). Analyses were each performed as duplicates. MALDI-TOF analysis was validated for 15 clinical isolates each of H. influenzae, Haemophilus parainfluenzae, and M. catarrhalis, all of which were identified correctly. The identity of the Haemophilus spp. and the M. catarrhalis isolates was determined using standard identification procedures such as growth dependency on factor V and X and positive oxidase and DNase reaction, respectively.
All H. influenzae strains recovered before the multidrug-resistant S1046 strain isolated from tracheal secretion of the patient in May 2011 were tested for antimicrobial susceptibility according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) criteria using the agar disk-diffusion method. The H. influenzae S1046 strain was tested for susceptibility to ampicillin/sulbactam, cefazolin, cefotaxime, ciprofloxacin, meropenem, and clarithromycin by the Etest method (Biomerieux, Nürtingen, Germany), whereas ampicillin and cefuroxime were tested by the agar disk-diffusion method.
Polymerase chain reaction and sequence analyses of resistance genes
The H. influenzae strain S1046 was analysed for several resistance genes. The beta-lactamase genes blaTEM and blaROB-1 as well as the acetyltransferase gene aac(6′)-Ib-cr and the fluoroquinolone resistance genes of qnr-type, including qnrA, qnrB, qnrC, qnrD, and qnrS, were amplified using primers as described previously.7,10,11 To detect mutations in the ribosomal protein genes L4, L22, and the domain V of the 23S rRNA gene, the DNA gyrase and topoisomerase IV genes, gyrA and parC, and the ftsI gene encoding the transpeptidase region of PBP3, primers were used as described15,18,23 using the total genome sequence of H. influenzae Rd strain (The Institute for Genomic Research) for comparison.
All amplicons were sequenced by using an ABI PRISM BigDye 3.1 Terminator Cyle Sequencing Ready Reaction kit (Applied Biosystems).
Results and Discussion
We report the case of a CVID patient suffering from chronic infection with H. influenzae causing multiple episodes of bronchopneumonia since March 2010. Replacement therapy with immunoglobulins since 1998 was not sufficient to prevent from bronchopulmonal infections, but probably reduced the frequency and severity of infections as described. 16 During exacerbation episodes, H. influenzae was isolated repeatedly from different respiratory materials like sputum, tracheal secretion, bronchoalveolar lavage, and lung biopsy, indicating that H. influenzae was the infectious agent of bronchopneumonia (Fig. 2). Additionally, in September 2010, H. influenzae was isolated with high bacterial counts from bronchoalveolar lavage and lung biopsy without contamination by the respiratory flora. However, H. influenzae was not detected in two throat swabs and one sputum sample (Fig. 2). In these cases, H. influenzae might have been overlooked since flora was present with high bacterial counts. In addition, throat swabs were cultivated 24 hr under aerobic conditions not including a BHI agar plate. Therefore, the fastidious bacterium H. influenzae, the growth of which can be enhanced in a CO2-enriched atmosphere, 25 possibly remained undetected.
Deficiency of immunoglobulins in CVID patients causes restriction of complement-mediated attack. Hence, patients are at high risk to develop infections with H. influenzae, which is the most common bacterium isolated during exacerbations. 21 Moreover, H. influenzae has developed several strategies to resist complement-mediated attack. 12 In addition to H. influenzae, M. catarrhalis, also a common pathogen causing respiratory infections in hypogammaglobulinaemic patients, 13 was isolated only once from the patient's sputum in May 2011. It has been described that these two pathogens collaborate to overcome the innate immune system as outer membrane vesicles produced by M. catarrhalis neutralize C3, and thus contribute to increased survival of H. influenza. 24 A possible infection of the patient with mycobacteria was excluded in September 2010 by microscopy, polymerase chain reaction (PCR), and cultivation methods.
As shown in Fig. 2, between July 2007 and May 2011, the patient underwent several antibiotic therapies, including fluoroquinolones (moxifloxacin and ciprofloxacin), macrolides (clarithromycin), beta-lactams (piperacillin/tazobactam, ceftriaxone, and meropenem), and aminoglycosides (gentamicin). Subsequently, in September 2010, H. influenzae showing antimicrobial resistance to ampicillin and ciprofloxacin was isolated from different clinical specimens taken on the same day, although the duration and dosage of the previous therapies were appropriate (Fig. 2). Finally, in May 2011, the multidrug-resistant H. influenzae isolate S1046 was isolated from sputum and tracheal secretions showing resistance to ampicillin, ampicillin/sulbactam, cefazolin, cefuroxime, ciprofloxacin, and clarithromycin. High bacterial loads and the beginning abscess formation (Fig. 1) might have contributed to the emergence of multidrug resistance. Resistance of S1046 to clarithromycin was associated with a high minimal inhibitory concentration (MIC) value (48 μg/ml) comparable to that observed in previously described clarithromycin-resistant H. influenzae clones (MICs 32– >128 μg/ml) obtained by repeated subcultivation in a clarithromycin-containing medium. 3 PCR and sequencing analyses of the H. influenzae isolate S1046 revealed a mutation in the ribosomal protein gene L4 (Gly65Asp) conferring resistance to macrolides in clinical H. influenzae strains as described.18,20 Mutations in the L22 and 23S rRNA genes were not detected. Additionally, the multidrug-resistant H. influenzae isolate S1046 was positive for the beta-lactamase gene blaTEM-1, whereas blaROB-1 was negative. Production of TEM-1 is the most common mechanism of ampicillin-resistance in H. influenza, 25 whereas 80% of the ROB-1-positive strains additionally show resistance to cefaclor. 14 Alterations of PBP3 can also cause resistance to cefaclor as well as to cefuroxime and beta-lactamase/inhibitor combinations such as amoxicillin/clavulanate. 25 By sequence analysis of the ftsI gene encoding the transpeptidase region of PBP3, two mutations (A502T and R517H) were detected in H. influenzae S1046. As previously described, the amino acid change A502T may affect susceptibility to cefaclor, 1 whereas cefuroxime resistance can be attributed to R517H. 23 Many of the H. influenzae strains with combined beta-lactamase production and the presence of altered PBP3 show only a slight decrease in amoxicillin–clavulanate susceptibility. 4 Accordingly, the H. influenzae S1046 isolate described here showed only low-level resistance to ampicillin/sulbactam (MIC value 1.5 μg/ml).
Ciprofloxacin-resistance in H. influenzae due to mutations in the chromosomally encoded DNA gyrase or topoisomerase IV genes is very rare. 8 In the H. influenzae isolate S1046, two mutations in the DNA gyrase gene gyrA (Ser84Leu and Asp88Asn) and one mutation in the topoisomerase IV gene parC (Ser84Ile) were identified, most likely contributing to the high-level ciprofloxacin-resistance phenotype (MIC value 24 μg/ml) as described. 8 Additionally, H. influenzae S1046 harbored the transferable plasmid-mediated acetyltransferase gene aac(6′)-Ib-cr, which has been described to confer resistance to aminoglycosides and fluoroquinolones. 22 The aac(6′)-Ib-cr gene has been recently detected in H. parasuis strains from pigs in South China, 11 but it has not been reported in H. influenzae so far. Plasmid-mediated fluoroquinolone resistance genes of qnr-type such as qnrA, qnrB, qnrC, qnrD, and qnrS were not detected in H. influenzae S1046.
Overall resistance rates in H. influenzae clinical isolates from our hospital are comparably low. Among all H. influenzae (n=368) isolated between March 2010 and May 2011 at our hospital, we found isolates being nonsusceptible to ampicillin (n=36; 9.8%), ampicillin/sulbactam (n=3; 0.8%), cefazolin (n=19; 5.2%), cefuroxime (n=4; 1.1%), and ciprofloxacin (n=1; 0.3%). Compared to the overall prevalence of beta-lactamase in H. influenzae of 15.0% stated in a global study, 7 the percentage of H. influenzae isolates resistant to beta-lactam antibiotics at our hospital was low. The multidrug-resistant isolate S1046 was the only ciprofloxacin-resistant H. influenzae isolate detected at our hospital in the above-mentioned period of time. This is in accordance with an international surveillance study reporting a very low rate (0.15%) of H. influenzae isolates with reduced susceptibility to fluoroquinolones. 2
We report the emergence of a multiresistant H. influenzae strain (S1046) in a CVID patient in consequence of previous antibiotic therapies. Since the H. influenzae strains recovered from the patient before S1046 were not stored and were not available for further investigations such as epidemiological typing at a molecular level, we cannot demonstrate persistence of a single H. influenzae isolate during the investigation period, and therefore cannot exclude that the patient carried different isolates of which the isolate from May 2011 finally substituted the previous isolates.
This is the first description of a H. influenzae strain harboring the plasmid-encoded gene aac(6′)-Ib-cr. One could speculate that low fluoroquinolone resistance levels caused by this gene might have contributed to the selection of DNA gyrase and topoisomerase IV mutants and might facilitate increasing dissemination of fluoroquinolone resistance among Haemophilus spp. by horizontal gene transfer.
Footnotes
Disclosure Statement
No competing financial interests exist.