
Abstract
Staphylococcus pseudintermedius is an opportunistic pathogen that is one of the most frequent causes of infections in dogs. In Europe, there are increasing reports of methicillin-resistant S. pseudintermedius (MRSP), and in Sweden, MRSP has also been more frequently isolated during recent years. However, there is limited knowledge regarding the epidemiology and genetic relationship among the Swedish isolates. This study therefore investigated the genetic relationship of MRSP isolated from companion animals in Sweden. In the study, MRSP isolates taken in the period January 2008–June 2010 from a total of 226 dogs and cats were characterized by spa typing, pulsed-field gel electrophoresis, and antimicrobial susceptibility testing. In addition, the geographical distribution of the isolates based on year of isolation and genetic typing was determined using a geographical information system. One multiresistant clonal lineage dominated among Swedish MRSP isolates, corresponding to the European winning lineage ST71-J-t02-SCCmec II-III. Furthermore, the geographical dissemination of MRSP corresponded to areas with high dog densities, centered on the three major cities in Sweden where the largest animal hospitals are situated.
Introduction
In Sweden, the number of confirmed clinical cases of methicillin-resistant S. pseudintermedius (MRSP) in animals has increased since MRSP was first detected in 2006 and in the period 2006–2010 was about 400. 5 The increasing prevalence is a major concern due to limited antimicrobial treatment options, because almost all isolates are multiresistant.5,23 In Europe, one lineage MLST type ST71 spa-type t02-carrying SCCmec II-III appears to dominate, whereas there are indications that another lineage ST68 spa-type t06 carrying SCCmec V has emerged in North America.7,23,27 However, knowledge about the epidemiology and genetic relationship among MRSP isolates isolated from animals in Sweden is very limited. The clonal spread of MRSP differs from that of methicillin-susceptible S. pseudintermedius (MSSP), which has been shown to have a high genetic polymorphism, although the occurrence of antimicrobial resistance is also common in MSSP.5,20
As a zoonotic bacterium, S. pseudintermedius appears not to be common, nor is S. intermedius, but there are reports of infections with varying degree of severity in humans.9,24,32,33,36 In addition, it has been reported that MRSP may have the potential to colonize humans and that people living and/or working with animals are more prone to carry MRSP.16,22,35
The aims of the present study were to examine the genetic relationship between MRSP isolates obtained from dogs and cats in Sweden and to position the Swedish situation in relation to an international context. These aims were achieved through molecular characterization and applying the geographical information system (GIS).
Materials and Methods
Clinical bacterial isolates and susceptibility testing
The study included isolates or specimens submitted for analysis to the Swedish National Veterinary Institute by veterinary clinics or private laboratories in the period January 2008–June 2010. Only the primary isolate from each individual was used. All MRSP isolates were verified as being S. pseudintermedius and carrying the mecA gene, using conventional polymerase chain reaction (PCR).1,29 The sample sites yielding MRSP were identified from the submission forms or by direct contact with the laboratory.
Minimum inhibitory concentrations (MICs) of the antimicrobial agents were determined using the VetMIC GP-mo microdilution panels (National Veterinary Institute, Uppsala, Sweden) with the following antimicrobial classes: aminoglycosides, β-lactam antimicrobials, chloramphenicol, fluoroquinolones, fusidic acid, lincosamides, macrolides, tetracyclines, and trimethoprim. The antimicrobials tested, together with their concentration ranges, are given in Table 1. S. aureus ATCC 29213 served as a quality control strain. The MIC breakpoints used for classification of isolates as resistant were those recommended in CLSI documents M100-S19 and M31-A3,10,11 with the exception of fusidic acid, for which the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoint for S. aureus was used. Isolates were defined as multiresistant if resistant to three or more antimicrobial classes.
Isolates from canine and feline clinical submissions. Resistance breakpoints are indicated with vertical black lines.
The dilution ranges tested for each antimicrobial are those contained within the white area. Values above this range indicate MIC values higher than the highest concentration in the range. Values below the lowest concentration tested indicate MIC values smaller or equal to the lowest concentration in the range.
Breakpoints for classification of isolates as resistant were those recommended for Staphylococcus (S.) spp. in the Clinical and Laboratory Standards Institute (CLSI) documents M100-S19 and M31 with exception for oxacillin and fusidic acid. For oxacillin, the breakpoint for S. pseudintermedius is according to Schissler et al. (2009) 30 and approved by the CLSI subcommittee on Veterinary Antimicrobial Susceptibility Testing. For fusidic acid, cutoff defined by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) for S. aureus was used.
MIC, minimal inhibitory concentrations.
Molecular characterization
spa typing was performed as described by Moodley et al. (2009). 15 The repeats and spa types were defined using the scheme adopted by Moodley et al. (2009) and using sequence signatures 5′-AATAATTCA and 3′-GACAAGCG defined by Ruscher et al. (2010). 27 SCCmec types I–V were determined using a multiplex real-time PCR assay. 36 To differentiate between SCCmec III lacking SCC-Hg, SCCmec II-III, and SCCmec VII-241, a conventional PCR was applied. 23
Chromosomal DNA and plugs (Agarose Prep; Amersham Biosciences, Uppsala, Sweden) for pulsed-field gel electrophoresis (PFGE) were prepared according to the Harmony protocol. 18 S. aureus NCTC8325 was used as control. To establish whether Swedish isolates belonged to the main European PFGE J-cluster described by Perreten et al. (2010), 23 five isolates belonging to the dominant European lineage ST71-J-t02-SCCmec II-III were included as references.
DNA was fragmented using 20 U SmaI (Fermentas, Vilnius, Lithuania), and fragments were separated in a CHEF-DR II system (BIO-Rad Lab, Hercules, CA) with a 1.2% agarose gel (Agarose NA; GE Healthcare, Uppsala, Sweden). The gel was run for 24 h at 5.6 V cm−1 with pulsed-time ramping from 2 to 5 s and at a temperature of 14°C.
Fragment patterns were analyzed in BioNumerics® version 6.6 (Applied Maths, Gent, Belgium) using DICE coefficient and Unweighted Pair Group Method with Arithmetic Mean cluster analysis, and fragments between 9 and 117 kb were included in the analysis. Position optimization was set at 0.5% and tolerance at 1.2%. PFGE patterns with a ≥80% homology value were assigned to the same cluster.
Geographical information system
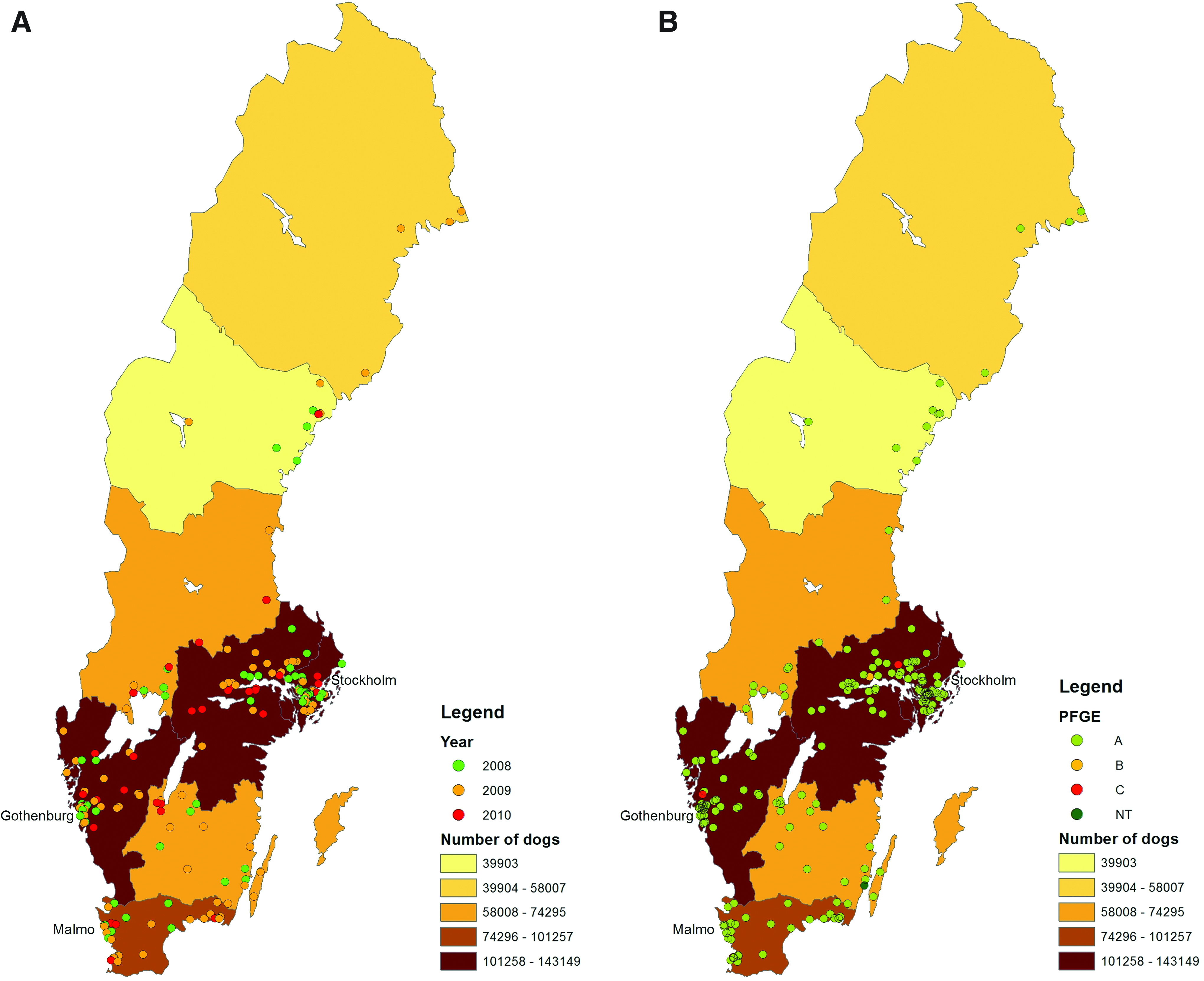
Each isolate was allocated the geographical coordinates representing the corresponding geometric centers (centroids) of the 5-digit-postcode area from which it originated using various techniques in the software ArcGIS 10 (ESRI; Redlands, CA). The zip code used was that of the animal owner, and not the hospital. The position of isolates with the same zip code was manually shifted slightly inside the zip code area to ensure visibility. Information for all cases was stored in a geodatabase, from which separate thematic maps were generated for each year and for each PFGE type and spa-type. Ocular inspection was then used to decide which maps should be presented separately and which maps could be collapsed into one.
The dog population was also displayed on the maps by using the number of dogs in each NUTS2 area of Sweden. 31
Results
Isolates, molecular typing, and GIS
In total, the study included isolates from 215 diseased individuals, 8 carriers, and 3 isolates of unknown origin (214 dogs and 12 cats). The sampling sites yielding MRSP are shown in Table 2. The MRSP isolates were assigned to four different PFGE clusters, but the majority of the isolates (216/226; 96%) occurred in one cluster, cluster A (Table 3). However, there is a possibility that this dominating cluster should be split in two, because 28 isolates (13%) of the 216 with 80% similarity formed a subcluster. Two distinct PFGE patterns were more common in cluster A, with 35 isolates (16%) and 30 isolates (14%), respectively. Cluster A corresponds to the cluster J described by Perreten et al. (2010). 23 Cluster B comprised 2 isolates, cluster C 4 isolates and cluster D 1 isolate, whereas 3 isolates were nontypeable using SmaI degradation.
Pooled samples taken from multiple sites like pharynx, gingival, and perineum.
Bacterial strains sent to the National Veterinary Institute for methicillin-resistant S. pseudintermedius confirmation.
R and bold values indicate strains classified as resistant and S=strains classified as sensitive to the tested antimicrobials.
Em, Erythromycin; Cm, Chloramphenicol; Cl, Clindamycin; Tc, Tetracycline; Fu, Fusidic acid; Gm, Gentamicin; Km, Kanamycin; Ci, Ciprofloxacin; Tm, Trimethoprim; NT, Non-Typeable.
In total, 5 different spa types were identified (Table 3): t02 (210 isolates, 93%), t06 (4 isolates), t10 (2 isolates), t29 (2 isolates), and t35 (1 isolates), whereas 6 isolates were nontypeable regarding spa. Isolates belonging to spa types, t02, t06, t10, and t29, carried the mecA on a SCCmec II-III, with the exception of one isolate belonging to t10, which carried a nontypeable SCCmec. The isolates belonging to t35 carried a nontypeable SCCmec, whereas the isolates that were nontypeable using spa carried a SCCmec IV (4 isolates) or a nontypeable SCCmec (2 isolates). All isolates typed as t02-SCCmec II-III, t06-SCCmec II-III, t10-SCCmec II-III, and t29-SCCmec II-III corresponded to PFGE cluster A (Table 3), with the exception of the two isolates, one t02-SCCmec II-III and one t10-SCCmec II-III, which constituted cluster B. The isolates that were nontypeable with spa and carried SCCmec IV corresponded to cluster C, and the isolate that was nontypeable using both spa and SCCmec constituted cluster D. One of the isolates studied was nontypeable with all three methods applied.
Figure 1 shows the annual geographical distribution of isolates and of PFGE patterns. On ocular inspection, there seemed to be no differences in the annual distribution, whereas the spatial distribution of isolates with PFGE pattern A corresponded roughly to the distribution of the background dog population. The rarer PFGE pattern isolates also occurred within the areas with the highest density of dogs. The geographical dissemination of MRSP centered on the three major cities in Sweden.
Susceptibility testing
The resistance and distribution of MICs for the different antimicrobial classes are shown in Table 1. All isolates were multiresistant, the majority of isolates (93%) being resistant to five or more antimicrobial classes, and one isolate was resistant to all antimicrobials tested (Tables 1 and 3). The resistance to the different classes varied from 8% for tetracycline to 100% for kanamycin. There was also a variation in MIC values for β-lactam antimicrobials, where one isolate was below the breakpoint for oxacillin (0.12 mg/L). For cephalotin, most MICs clustered on >8 mg/L, but seven isolates displayed lower MICs for cephalotin varying from 0.25 to 4 mg/L. With one exception, a t02-SCCmec II-II, these isolates carried a SCCmec IV or a nontypeable SCCmec, and five of the isolates were nontypeable with spa. The isolate with MIC of 0.12 mg/L for oxacillin had an MIC of 0.5 mg/L for cephalotin and was a t10 with nontypeable SCCmec and PFGE.
Two resistance profiles dominated among the isolates (Table 3), all belonging to PFGE cluster A. Of these, 90 (42%) isolates were susceptible only to tetracycline and fusidic acid, and 107 (50%) were susceptible to tetracycline, fusidic acid, and chloramphenicol. Three isolates were only resistant to fusidic acid. The multiresistant phenotypes in combination with their molecular characterization are shown in Table 3.
Discussion
In 2006, the first MRSP isolates were described in Sweden, and during that year, 13 dogs were confirmed with MRSP. In addition, two nosocomial outbreaks of MRSP occurred at two of Sweden's largest animal hospitals. 6 Two representative isolates from the outbreaks were included in the survey by Perreten et al. (2010), 23 and both belonged to the major European lineage. The current study included all confirmed MRSP isolates from dogs and cats in Sweden during 2008, 2009, and the first 6 months of 2010, and revealed that the clear majority of these belonged to the same clonal lineage (Table 3) described by Perreten et al. (2010). 23 International studies have clearly shown that there is one dominant clonal lineage within Europe, while another lineage appears to have emerged in North America.23,27 However, recently, Osland et al. (2012) reported that the Norwegian MRSP population has a more heterogeneous clonal distribution. 21 The reason for such a difference between geographically close countries is unclear and needs further attention. In the current study, less-common strains were also identified (Table 3). It is possible that these strains might be more successful later on and constitute a larger part of the Swedish MRSP population. It is worth noting that the majority of the less-common strains were generally nontypeable by spa typing, which can be due to differences in the primer sites. However, in our laboratory, it has been observed that MSSP is generally nontypeable when applying the current spa-typing scheme (data not shown), as described first by Moodley et al. (2009), where the authors also noticed a low typeability rate for MSSP. Together with the results from the current study, this may indicate that the spa gene appears only to be present in some members of the species, casting doubt on its usefulness in epidemiological investigations, at least for lineages other than the dominant European type.
The GIS analyses showed that the geographical dissemination of MRSP was centered on the three major cities in Sweden, Stockholm, Gothenburg, and Malmö, which correspond to the highest dog density areas in Sweden (Fig. 1). In addition, the first confirmed cases of MRSP occurred in animal hospitals situated in the Stockholm area and has since managed to spread from this area throughout southern Sweden. In densely populated areas, spread of infectious diseases may be facilitated by frequent animal contacts, but in the present case, the spread could also be explained by the fact that the largest animal hospitals in Sweden are located in these areas. In 2006, almost 60% of the MRSP isolates at that time were from surgical-site infections (SSI), and the possibility that isolates in this study originated from SSI cannot be ruled out. 6 Poor infection control may therefore have facilitated the spread, as staphylococci can be spread through direct or indirect contact, with human hands, tools, or equipment serving as the indirect route of transmission. However, this does not explain why the ST71-J-t02-SCCmec II-III lineage has spread more successfully than the others. One partial explanation may be this lineage has an ability for high-level biofilm production, which may give it an advantage over other lineages. 21
In the present study, all MRSP isolates were multiresistant. In fact, the majority of them were resistant to five or more antimicrobials and were only susceptible to two classes of antimicrobials authorized for systemic treatment of dogs in Sweden: fusidic acid and tetracyclines. A similar multiresistance pattern has previously been described by Perreten et al. (2010) 23 and Ruscher et al. (2009). 26 This will exert pressure on veterinarians when selecting an optimal antimicrobial therapy, due to the limited treatment options. The current empirical antimicrobial treatment, which usually involves a β-lactam antimicrobial, will be ineffective. In acute severe cases when the rapid response to antimicrobial treatment is critical, there is also a high risk of patient death or severe outcome if empirical treatment fails. In addition, antimicrobial therapy with a substance to which multiresistant bacteria are resistant will favor their growth during treatment and therefore promote spread. Even though investigations have shown that MRSP isolates are susceptible to antimicrobials of last resort in humans, that is, vancomycin, linezolid, and the combination quinupristin/dalfopristin,23,27 it is questionable whether the veterinary use of these substances for the treatment of MRSP infections is justifiable.
All the isolates analyzed in this study were considered to be methicillin-resistant, because they all carried the mecA gene. However, one isolate had an MIC for oxacillin below the S. pseudintermedius applicable CLSI breakpoint of ≥0.25 mg/L, 30 and six isolates had deviating lower MICs for cephalotin compared with the majority. Notably, these diverging MICs were observed in isolates with other spa types and other SCCmec types than the dominant t02-SCCmec II-III. Similar observations have been described for S. aureus, 8 and a possible explanation is that different SCCmec differ in their responses to β-lactams to induce expression of the mecA gene. 14 Furthermore, differences in susceptibility to β-lactams such as cefoxitin have also been reported in different methicillin-resistant staphylococci.3,4 Therefore a possible explanation for this phenomenon might be differences in SCCmec types, staphylococcal species differences, or a combination of both, but this issue needs to be further investigated.
In conclusion, it was shown that a dominant multiresistant MRSP lineage, spa-type t02 SCCmec II-III, has successfully spread in a relatively short time through the areas of Sweden with the highest dog densities. This finding indicates an urgent need for improved infection control and decreased use of antimicrobials in Swedish small-animal medicine to limit the further spread of MRSP.
Footnotes
Disclosure Statement
No competing financial interests exist.