
Abstract
We compared the prevalence of cultivable antibiotic-resistant bacteria and resistance genes in the surface microlayer (SML) and underlying waters (UW) of an estuary. Prevalence of resistant bacteria was determined in antibiotic-supplemented agar. Bacterial isolates from the UW (n=91) and SML (n=80), selected in media without antibiotic, were characterized concerning susceptibility against nine antibiotics. The presence of genes blaTEM, blaOXA-B, blaSHV, blaIMP, tet(A), tet(B), tet(E), tet(M), cat, sul1, sul2, sul3, aadA, IntI1, IntI2, and IntI3 was assessed by PCR. The variable regions of integrons were sequenced. Ampicillin- and streptomycin-resistant bacteria were significantly more prevalent in SML. Resistance levels among the bacterial collections were generally low, preventing detection of significant differences between SML and UW. The tet(E) gene was detected in two Aeromonas isolates and tet(M) was detected in a Pseudomonas isolate. Gene sul1 was amplified from three Aeromonas isolates. Prevalence of intI genes was 2.11%. Cassette arrays contained genes encoding resistance to aminoglycosides and chloramphenicol. A higher prevalence of antibiotic-resistant bacteria in the SML, although only detectable when bacteria were selected in antibiotic-supplemented agar, suggests that SML conditions select for antibiotic resistance. Results also showed that antibiotic resistance was uncommon among estuarine bacteria and the resistance mechanisms are probably predominantly intrinsic.
Introduction
The SML is distinct from underlying waters (UW), 27 since higher fluctuations in salinity and temperature occur and the exposure to ultraviolet radiation is also greater than in UW. Also, in the SML, higher amounts of organic compounds (proteins, lipids, and polysaccharides) and anthropogenic contaminants (hydrocarbons, metalloids, pesticides, and antibiotic residues) accumulate.7,11,10 Moreover, even under greater selective pressures, the SML harbors a more abundant and active bacterial community, the bacterioneuston, comparing to UW.2,16,37
The gelatinous nature of the SML combined with strong selective pressures and high bacterial densities stimulate the production of secondary metabolites. 29 In fact, for example, antibiotic producers have been frequently retrieved from the SML. 14
The production of antimicrobials, together with persistence of anthropogenic-derived antibiotic residues in the SML 39 may select for and foment the dissemination of antibiotic resistance genes (ARGs). In addition, bacterial growth in biofilms, expected to occur in the SML, might also promote horizontal gene transfer and resistance to antimicrobial agents.20,10,35 Thus, the SML may be a natural reservoir of ARGs and an incubator of new gene combinations.
However, most studies concerning antibiotic bacterial resistance in estuarine and coastal water systems focused on bulk water,18,25,13 and consequently the relevance of the SML in developing, selecting, and spreading ARGs has been almost ignored.
Estuarine systems are strongly influenced by anthropogenic activities accumulating high levels of pollutants and often becoming eutrophicated, 23 and thus gathering ideal conditions for the establishment of a distinct bacterioneuston community.
In a previous study, we have focused our attention on the phylogenetic composition of the bacterioneuston and bacterioplankton communities from a highly polluted estuarine system (Ria de Aveiro, Portugal). 2 The aims of the present study are (1) to compare the prevalence of cultivable antibiotic-resistant bacteria in the SML and UW in Ria de Aveiro; (2) to compare antibiotic resistance profiles between isolates previously obtained from SML and UW samples; (3) to assess and compare the occurrence and diversity of ARGs and integrons in the same isolates.
Material and Methods
Heterotrophic plate counts on antibiotic-selective media
SML and UW samples were collected on three occasions (C1, C2, and C3) in 2008 at three sites named Cais do Chegado (CC), Costa Nova (CN), and Cais do Sporting (CS) within the estuary Ria de Aveiro (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr) as described by Azevedo et al. 2 Briefly, SML samples (250 ml) represented by a water layer of 60–100 μm were collected with an acrylic or a glass plate. 2 UW (1 L) were collected at ∼0.4-m depth in 2-L autoclaved bottles.
From each water sample, 100 μl were directly spread onto EA and GSP plates without added antibiotics. Also, 1 ml of each sample was serially diluted in 0.9% NaCl and 100 μl of each dilution was spread in the same media to obtain colony counts between 30 and 300 per plate. Total heterotrophic plate counts (HPC) were determined using these plates.
Resistant bacteria were cultivated by the same spreading procedure in the same culture media supplemented with ampicillin (50 μg/μl), tetracycline (20 μg/μl), and streptomycin (10 μg/μl). These concentrations are above the breakpoints defined by the Clinical and Laboratory Standards Institute (CLSI) 8 for most phylogenetic groups and were chosen to select for highly resistant bacteria. Colony-forming units (CFU/ml) were counted in triplicate (three independent plates) after 4 days of incubation at 22°C in the dark.
Bacterial isolates
A total of 352 bacterial isolates were obtained from SML and UW within the estuary Ria de Aveiro in a previous work, corresponding to 10 to 20 isolates selected from each sample. 2 From those, 13 isolates representing 12 genera were impossible to recover in successive cultures and were excluded from the current study. Isolates were typed using REP-PCR with primers REP-1R and REP-2I as described previously (Versalovic et al.). 38 PCR reactions were carried out with Taq polymerase, nucleotides, and buffers from Promega. The PCR reaction mixtures (25 μl) contained 1×PCR buffer, 200 μM of each nucleotide, 3 mM MgCl2, 5% dimethylsulfoxide, 7.5 pmol of each primer, 1 U of Taq polymerase, and 50–100 ng purified DNA. Amplification was carried out as follows: initial denaturation for 7 min at 95°C, followed by 30 cycles of denaturation for 1 min at 94°C, annealing for 1 min at 40°C and extension for 8 min at 65°C and by a final extension of 16 min at 65°C. The products were electrophoresed for 90 min under a constant voltage of 80 V on a 1.5% (w/v) agarose gel containing 0.5×TAE (20 mM Tris–acetate, pH 7.4, 10 mM sodium acetate, 0.5 mM Na2EDTA) and DNA markers purchased from MBI Fermentas. The gel images were acquired using a Molecular Imager FX system (Bio-Rad Laboratories) and analyzed using the software package GelCompar 4.0 (Applied Maths). From 339 isolates, a total of 171 isolates displaying different REP profiles were identified based on the partial sequence of the 16S rRNA gene. 2 Isolates (80 isolates from SML and 91 from UW) affiliated with 34 genera within six classes (Table 1) and were included in the present study.
Isolates selected as representative of different REP profiles.
SML, surface microlayer; UW, underlying waters.
DNA extraction
Isolates were cultured in Luria-Bertani agar plates (Merck) and were incubated at 30°C overnight. The total DNA was obtained by resuspending two isolated colonies in 100 μl of buffer B1 (50 mM Tris-Cl pH 8.0; 50 mM EDTA pH 8.0; 0.5% Tween 20; 0.5% Triton X-100) and 100 μl of master-mix (1 ml of buffer B1; 2 mg/ml of Lysozyme; 4.5 mg/ml of Proteinase K: 7.5 U of RNAse A). Cell suspension was mixed vigorously and incubated for 30 min at 37°C. Following, 70 μl of B2 (3 M Guanidine Hydrochloride; 20% Tween 20) were added and the mixture was incubated for 30 min at 50°C. Subsequently, the DNA Extraction Kit (#K0513–MBI; Fermentas) was used according to the manufacturer's instructions.
Antimicrobial susceptibility testing
Antimicrobial susceptibility was tested by the agar disk diffusion method as recommended by the CLSI. 8 Isolates representing different REP profiles (n=171) were tested for susceptibility to 9 antimicrobial agents representing six classes of antibiotics: (1) β-lactams—ampicillin (10 μg), imipenem (10 μg), cephalothin (30 μg); (2) aminoglycoside—streptomycin (10 μg), gentamicin (10 μg); (3) phenicols—chloramphenicol (30 μg); (4) tetracyclines—tetracycline (30 μg); (5) the combination sulfamethoxazole/trimethoprim (25 μg) and (6) quinolones—nalidixic acid (30 μg). Disks were purchased from Oxoid. Antimicrobial activities were determined as clear zones of inhibition around the antibiotic disks, after incubation at 30°C for 24 hr. Isolates were classified as sensitive, intermediate, or resistant taking into account the CLSI guidelines. 8 Whenever genus-specific guidelines were not available, guidelines for the Enterobacteriaceae were used for Gram-negative strains and guidelines for Staphylococcus spp. were used for Gram-positive strains. Escherichia coli ATCC 25922 was used as a quality control strain.
Antimicrobial resistance genes
All isolates were tested by PCR for the presence of genes conferring resistance to β-lactams, tetracyclines, chloramphenicol, aminoglycoside, and sulfonamides by using primers and PCR conditions previously reported18,19 (see also Table 2). Isolates carrying resistance genes were included in each PCR run as positive controls. The positive controls for amplification of the sulfonamide resistance genes sul1, sul2, and sul3 were as described previously.18,9 A PCR mixture with no DNA added was used as a negative control. Gel electrophoresis was performed on 1.5% agarose gel and stained with ethidium bromide.
Detection and characterization of integrons
The presence of IntI1, IntI2, and IntI3 genes, encoding class 1, 2, and 3 integrases, was investigated by PCR as previously described and using the same positive controls 30 (see Table 2). All integrase-positive isolates were examined to determine the sizes of integrons variable regions. Primer set 5′CS/3′CS was used to amplify the class 1 integron gene cassette region according to what has been described elsewhere 30 (see Table 2). PCR products were purified with the Jetquick PCR Product Purification Spin Kit (Genomed) and used as a template in the sequencing reaction carried out by the company GATC (Germany). When bands with different sizes were amplified, PCR products were cloned using a TA cloning kit (Invitrogen) according to the manufacturer's instructions. Clones representative of different amplicons were sent for sequencing. Online similarity searches were performed using the BLAST software. 1
Statistical analysis
Data retrieved from HPC on antibiotic-selective media were analyzed using the t-test through PAST version 2.04 (http://palaeoelectronica.org/2001_2001/past/issue2001_2001.htm). 15
Nucleotide sequence accession numbers
The nucleotide sequences were deposited in the GenBank nucleotide database under the following accession numbers: JX646699 to JX646702.
Results
Prevalence of antibiotic-resistant bacteria
The percentages of ampicillin-, streptomycin-, and tetracycline-resistant cultivable bacteria found in the two water layers are shown in Table 3. In general, resistance to streptomycin was the most prevalent (5.86% to 42.87% of total HPC), followed by ampicillin (2.29% to 12.37%), and tetracycline (0.87% to 7.21%). Levels of resistance were significantly higher for the SML samples (p-value<0.05) with only a few exceptions (resistance to tetracycline in the C2 and C3 campaigns). The prevalence of ampicillin-resistant bacteria in the SML was three to four times higher than in UW and streptomycin-resistant bacteria were two to three times more prevalent in the SML. For tetracycline, significant differences were only detected in the first campaign (five times more prevalent in the SML).
HPC, heterotrophic plate counts.
Antibiotic susceptibility testing
Susceptibility to all tested antibiotics was observed in 36% (61 out of 171) of the isolates. Predominant resistances were observed to cephalothin (46% of the isolates were resistant to this antibiotic), followed by ampicillin (40%), and streptomycin (34%). Isolates were most frequently susceptible to imipenem (only 6% of the isolates were resistant to this antibiotic), tetracycline (4%), and gentamicin (3%). Significant differences between isolates collected from SML and UW were not observed (Fig. 1).

Percentages of strains obtained from the surface microlayer (SML) and underlying waters (UW) that were resistant to each of the antibiotics tested. Antibiotic abbreviations: AMP, ampicillin; IMP, imipenen; TE, tetracycline; CN, gentamicin; NA, nalidixic acid; SXT, sulfamethoxazole/trimethoprim; C, chloramphenicol; S, streptomycin; KF, cephalothin.
Since antibiotic resistance profiles may be dependent on the phylogenetic groups, results were separately analyzed for the most commonly retrieved genera (Aeromonas, Acinetobacter, Pseudomonas, Psychrobacter, and Shewanella) as well as for the isolates classified as Enterobacteriaceae (Table 4). Most groups were highly resistant (resistance rates >30%) to ampicillin, cephalothin, and streptomycin. Lower levels of resistance to these antibiotics were only registered for Acinetobacter against streptomycin and for Shewanella against ampicillin. Additionally, Psychrobacter members were exceptionally sensitive (resistance rates <18%) to all antibiotics tested. All groups were highly susceptible (resistance rates <18%) to imipenem, gentamicin, and tetracycline. Resistance levels to chloramphenicol, nalidixic acid, and trimethoprim/sulfamethoxazole were generally low with a few exceptions: Acinetobacter and Pseudomonas were highly resistant to trimethoprim/sulfamethoxazole and Pseudomonas members were also highly resistant to chloramphenicol and nalidixic acid.
n, number of isolates affiliated to each phylogenetic group.
MDR, percentage of multiresistant strains (resistant to antibiotics included in 3 or more classes).
The overall frequency of multiresistant isolates (resistant to three or more classes of antibiotics) was 14% (24 isolates out of a total of 171 isolates). In terms of multiresistance, results obtained for SML and UW isolates were also similar (Fig. 2). For multiresistant isolates, 16 antibiotic resistance patterns were observed (Table 5). The most common resistance profile included resistance to ampicillin, nalidixic acid, streptomycin, cephalothin, chloramphenicol, and trimethoprim/ sulfamethoxazole (two isolates from SML and two from UW). The comparison between phylogenetic groups revealed that multiresistance was more frequent among Pseudomonas strains (Table 4).
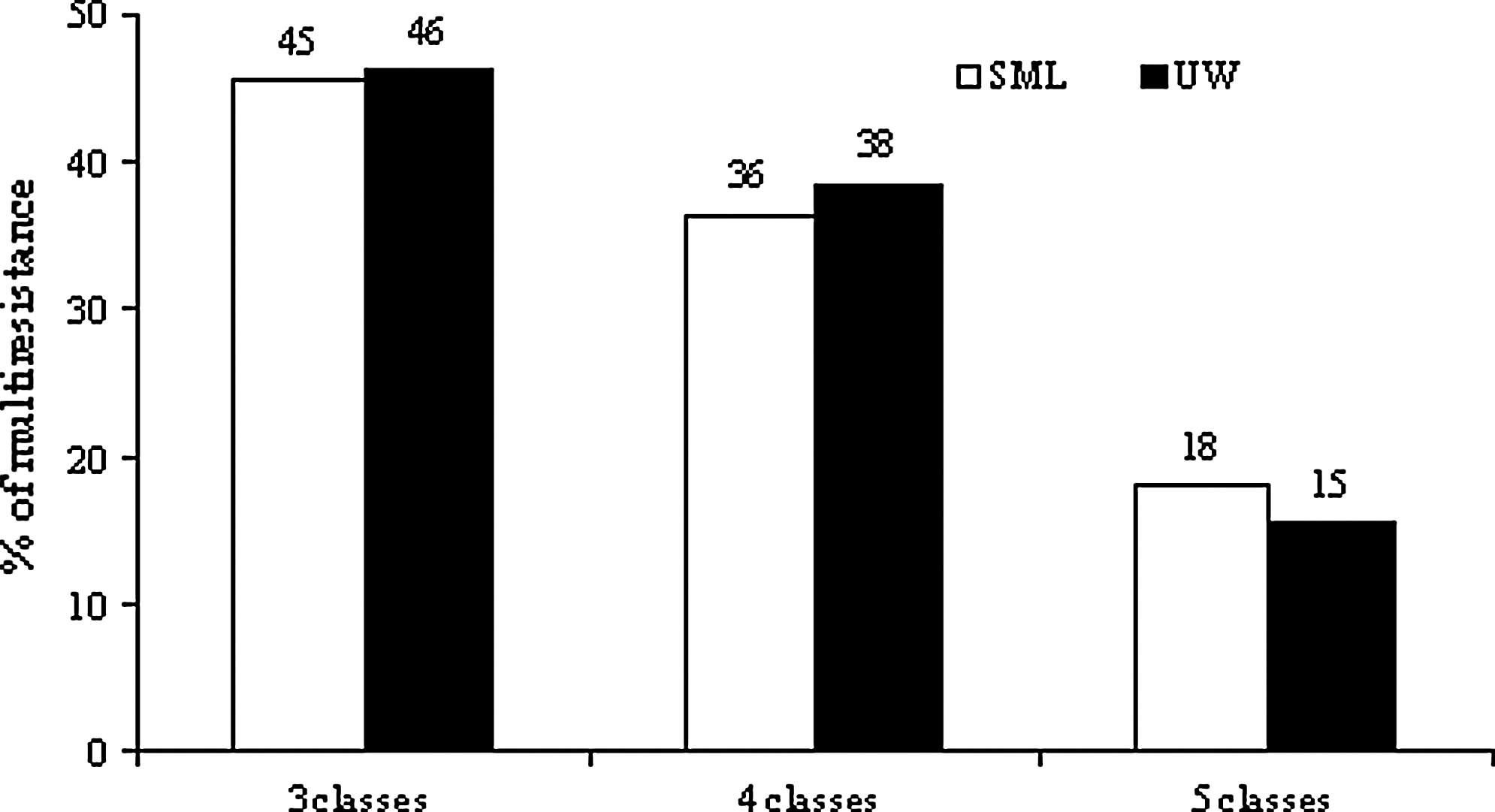
The percentage of multiple antibiotic-resistant bacteria from the SML (bacterioneuston) and UW (bacterioplankton).
The most common pattern of multiple-drug resistance among both water collections. Antibiotic resistance patterns specific of bUW and cSML isolates.
AMP, ampicillin; KF, cephalothin; IMP, imipenem; TE, tetracycline; C, chloramphenicol; CN, gentamicin; NA, nalidixic acid; S, streptomycin; SXT, sulfamethoxazole/trimethoprim.
Screening for antibiotic resistance and integrase genes
The presence of genes conferring resistance to beta-lactams, tetracyclines, chloramphenicol, aminoglycoside, and sulfonamides was analyzed by PCR in all the 171 isolates (Table 2). Genes blaTEM, blaOXA-B, blaSHV, blaIMP, tet(A), tet(B), tet(E), tet(M), cat, sul1, sul2, sul3, and aadA were chosen as they have been the most frequently detected acquired genes both in clinical and environmental isolates. Surprisingly, despite the considerable high levels of resistance against beta-lactams and aminoglycosides, genes conferring resistance to these antibiotics were not detected. Genes conferring resistance to chloramphenicol were also not found. Among seven isolates displaying resistance to tetracycline, tet(E) was detected in two isolates belonging to genus Aeromonas, and tet(M) was detected in one Pseudomonas isolate. The gene sul1 was amplified from three isolates belonging to genus Aeromonas, and sul2 was detected in one Pseudomonas isolate.
The intI1 gene was present in 2.11% of the isolates, all affiliated with Aeromonas, while genes intI2 and intI3 were not detected in any isolate. The variable region of class 1 integrons was successfully amplified with the primers 5′CS/3′CS. Sequencing analysis revealed four different arrays: aadA1, aadA13, catB8 and catB8, aadA1. The last two arrays were amplified from the same isolate.
Discussion
We compared the persistence of antibiotic resistance among cultivable bacteria from the SML and UW in an estuarine system. We hypothesized that SML would be a reservoir of antibiotic resistance.
Specifically, we compared the prevalence of tetracycline-, ampicillin-, and streptomycin-resistant bacteria between both water layers. These antibiotics have been used for long not only in human medicine, but also in agriculture, aquaculture, and veterinary. 6 Accordingly, resistance to these drugs is highly disseminated in several environments.3,34 Results obtained during this study showed significant differences between SML and UW in terms of prevalence of resistance to all tested antibiotics. Differences may arise from the fact that SML accumulates pollutants, including antibiotics,20,39 and antibiotic concentrations in the environment are the main factors contributing to the selection of resistant bacteria. 24 In addition to the selective pressure exerted by antibiotics themselves, other compounds, such as heavy metals, disinfectants, or pesticides also contribute to the maintenance of antibiotic resistance. 22 SML has been described to accumulate nickel, copper, arsenic, chlorinated pesticides, and polychlorinated biphenyls.11,42 On the other hand, producers of antimicrobial compounds have often been described among particle-attached bacteria rather than free-living bacteria.14,40,41 Particle-attached bacteria may be enriched in SML due to accumulation of higher amounts of organic particles in this water layer. 10
Besides determining the prevalence of antibiotic-resistant bacteria in both layers using antibiotic-supplemented agar, during this study, we also compared antibiotic resistance profiles of isolates from SML and UW. Isolates were obtained in nonselective agar without an antibiotic. No significant differences in terms of resistance profiles or prevalence of multiresistance were detected between SML and UW isolates. These apparently contradictory results are probably due to the fact that generally resistant bacteria were a minor component of the SML and UW bacterial communities preventing the detection of differences between culture collections. In fact, antibiotic resistance levels among our bacterial collections were generally low and multiresistance was infrequent. For example, members of the genus, Psychrobacter, were highly sensitive to all antibiotics tested. Psychrobacter was the most frequently retrieved genus representing almost 17% of the total number of isolates. Although other studies reported considerable higher resistance levels in estuarine bacteria, 43 those studies were either focused on resistant bacteria selected on antibiotic-supplemented agar 12 or on specific bacterial groups known for their high resistance levels to several antibiotics. 18 When unbiased selections were conducted, results were similar to the ones obtained during this study.32,33
Comparative information on antibiotic resistance between SML and UW is still scarce. Even so, a few studies reported differences between SML and UW in terms of prevalence of resistant bacteria.20,22 Those studies were also based on counting antibiotic-resistant bacteria in antibiotic-supplemented agar.
In this study, colonies were randomly selected from Petri dishes, which may have excluded some of the less common members of the community. Even so, this procedure was adopted since colony selection based on morphological traits commonly results in overrepresentation of these less prevalent members. Also, the representativeness of our culture collection was improved by collecting a high number of samples and by using two different culture media.
We assessed the presence and diversity of acquired ARGs commonly detected both in clinical and environmental isolates. As previously described, antibiotic resistance phenotypes and genotypes may not match. 18 Thus, we decided to screen for the presence of resistance genes in all the isolates independently of their resistance phenotypes. The presence of the ARGs in isolates with a susceptible phenotype is worth evaluating, since in combination with other resistance mechanisms or in a different genomic context these genes may confer or potentiate resistance.
Most of the times we failed to detect any of the inspected genes even in resistant isolates. For example, the presence of the common acquired bla genes encoding β-lactamases was not demonstrated in any of the isolates, even when an ampicillin resistance phenotype was detected. Again, our results may be due to the fact that an unbiased selection was conducted. This strategy resulted in a collection of isolates belonging to a wide range of genera, for some of which antibiotic resistance mechanisms had never been characterized. These mechanisms may be intrinsic rather than the acquired ones inspected during this study. For example, trends of antibiotic resistance were essentially the same for the genera most commonly retrieved in our study and included high levels of resistance to ampicillin, cephalothin, and streptomycin. Intrinsic antibiotic resistance mechanisms to beta-lactams have already been described, such as the production of chromosomal-encoded beta-lactamases in Aeromonas, Pseudomonas, Acinetobacter, Vibrio, Stenotrophomonas, or Shewanella. 28 Additionally, Acinetobacter, Pseudomonas, and Aeromonas are noted for their intrinsic resistance to antibiotics due to several other mechanisms, such as the production of amynoglicoside-modifying enzymes, the diminished expression of outer membrane proteins, and the overexpression of efflux pumps.4,21 For these genera, the combination of several resistance mechanisms often occurs leading to the emergence of multiresistance strains.4,12 In our study, multiresistance was particularly associated to Pseudomonas and Aeromonas.
Concerning tetracyclines, the genes tet(E) and tet(M) were detected in Aeromonas and Pseudomonas, respectively. tet(E) has previously been described and has the most frequent tetracycline resistance mechanism in Aeromonas 19 and tet(M) is also common among Gram-negative isolates, including Pseudomonas. 5
The prevalence of integrons was also assessed and class 1 integrons were detected in 2.11% of our isolates. This prevalence is comparable to what has been reported for estuarine bacteria from other geographical locations. 36 All class 1 integrons were detected in Aeromonas isolates (two from SML and one from UW samples), predominantly carrying the aadA-type cassettes (aadA1, aadA13), conferring resistance to streptomycin. These cassettes are reported as the most frequent gene cassettes in bacterial isolates 31 and have been frequently detected in Aeromonas.18,30,26
Results obtained during this study must be interpreted having in mind that only the culturable fraction of the bacterial community was considered. We are aware that only a minor fraction of the environmental bacterial community can be cultivated under laboratory conditions. 2 However, the large majority of the studies on antibiotic resistance conducted so far are based on the characterization of pure cultures. The main reasons are related to the current limitations of culture-independent methodologies. 17 On one hand, fragments amplified from total DNA are not necessarily indicative of the presence of functional genes. On the other hand, the hosts of those genes usually cannot be identified by culture-independent methods, which present a severe limitation when studying antibiotic resistance.
In summary, the prevalence of antibiotic-resistant bacteria was different between the SML and UW only when selection was conducted using antibiotic-supplemented agar. Even so, these results suggest that SML conditions select for antibiotic resistance. The overall results also showed that, although antibiotic resistance occurs among estuarine bacteria, the resistance mechanisms are different from the most common acquired mechanisms, being probably predominantly intrinsic. Although we cannot exclude the contribution of transferable genetic elements, our results suggest that SML primarily selects for intrinsic antibiotic resistance.
Footnotes
Acknowledgments
This work was financed by the Fundacão para a Ciência e a Tecnologia (FCT) through grants SFRH/BPD/65820/2009 (CSO) and SFRH/BPD/63487/2009 (IH) and SFRH/BD/64057/2009 (JSNA). We are also grateful to A. Moura (Department of Biology, University of Aveiro) for reviewing English style and grammar.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.