
Abstract
The emergence and spread of antibiotic-resistant bacteria reflects both, a gradual, completely Darwinian evolution, which mostly yields slight decreases in antibiotic susceptibility, along with phenotypes that are not precisely characterized as “resistance”; and sudden changes, from full susceptibility to full resistance, which are driven by a vast array of horizontal gene transfer mechanisms. Antibiotics select for more than just antibiotic resistance (i.e., increased virulence and enhanced gene exchange abilities); and many non-antibiotic agents or conditions select for or maintain antibiotic resistance traits as a result of a complex network of underlying and often overlapping mechanisms. Thus, the development of new antibiotics and thoughtful, integrated anti-infective strategies is needed to address the immediate and long-term threat of antibiotic resistance. Since the biology of resistance is complex, these new drugs and strategies will not come from free-market forces, or from “incentives” for pharmaceutical companies.
Introduction
To emphasize the size and pace of the problem, it is worth mentioning that during the preparation of this article, three important crises developed which directly or indirectly involved antibiotic resistance: (1) the detection, first in the United Kingdom and subsequently in other Western countries, of multiresistant enteric bacteria, traced down to India, carrying the gene for a very-broad-spectrum beta-lactamase, NDM-1, among other things 95 ; (2) an outbreak of food poisoning with an enteroaggregative, Shiga-toxin-producing Escherichia coli (O104:H4) exhibiting blended virulence profiles (the exposure to some antibiotics might induce the mobilization of virulence genes carried in prophages) and producing an extended-spectrum beta-lactamase 133 ; and (3) a Japanese strain of Neisseria gonorrhoeae that is resistant to ceftriaxone, which is the “last resort” treatment of gonorrhea 129 (along with an increase in cefixime resistance in the United Kingdom from 0% in 2005 to 20% in 2010). Many other silent problems underlie these headline stories, such as the spread of colistin-sensitive-only strains of Pseudomonas aeruginosa 38 or the growing presence of methicillin-resistant Staphylococcus aureus (MRSA) in the community.
Forgotten Concepts of the Darwinian Evolution of Resistance
A common view of the evolution of resistance is an oversimplification that goes similar to this: One or a few organisms that are resistant to a given antibiotic that, by chance, emerge from within a large bacterial population are selected through the use of that antibiotic, say, during the treatment of an infectious episode (although this scenario is considered to be rare 108 ). These few resistant variants quickly multiply, filling the niche previously occupied by the decimated, susceptible population, and as a result, many or most of the bacteria are now resistant to that drug. Repeated exposure to different antibiotics increases multiresistance, which is currently common. Although this view is basically true, a number of potentially important details have been missed, which may be as important as the simple selection of an already fully resistant organism.
We should first consider that the word “resistance,” from the clinical point of view, does not encompass the full variety of altered susceptibility phenotypes selected by the presence of antibiotics. Clinically, an isolate is deemed resistant when the minimal inhibitory concentration (MIC) of the corresponding antibiotic is above a breakpoint which is set by agencies or committees, such as CLSI or EUCAST, that attempt to associate this phenotypic change with clinical failure. The particular case of diminished susceptibility that does not amount to this full resistance will be discussed next. Other phenotypes associated with clinical failure that do not necessarily modify the MIC are selected by antibiotics. For example, tolerance (a significant increase in the concentration needed to kill bacteria while MIC remains unchanged), which can arise by starvation 58 or mutations 21 could even protect against chemically unrelated antibiotics 68 ; subsistence (the ability of some bacteria to use antibiotics as a source of carbon) reduces the local concentration of antibiotics, thereby detoxifying and protecting surrounding organisms 14 ; and the peculiar dependence, only known to involve glycopeptide antibiotics, such as vancomycin, allows the growth of enterococci or staphylococci only in the presence of the antibiotic. 120 A separate paragraph should be devoted to the persistence of biofilms, 105 which may or may not be associated with the persistence of a small fraction of a susceptible bacterial population exposed to antibiotics, caused by phenotypic switching in growth rates. 12 The data from studies on the prevalence of resistance almost never include these phenotypes, as they cannot be detected with the routine methods in clinical microbiology labs. Many of these phenotypes have been described as “noninherited resistance.” 104 These phenomena also contribute to the discrepancies between the results of in vitro susceptibility testing and the clinical efficacy of antibiotics.
Biofilms are complex microbial consortia whose clinical relevance has only recently been analyzed. Most infectious episodes are caused by bacteria growing as biofilms, particularly those affecting tissues that are susceptible to fluxes, such as the tissues which line the respiratory and urinary tracts. 49 Biofilms persist after treatment with antibiotics, even if they are formed by bacteria that show a susceptible phenotype when growing freely (i.e., planktonically), as they are in routine susceptibility tests. The reasons for this persistence are not well understood, vary from species to species, and might involve the diminished permeation of the antibiotic through the biofilm, an altered bacterial physiology, and/or an antiapoptotic phenotype. 66 In some cases, the genes involved in antibiotic resistance, such as P. aeruginosa tssC1, which is associated with type VI secretion, 158 are specifically induced during biofilm growth. The initial step in the formation of a biofilm is the adherence of bacteria, which is also the first step of virulence; furthermore, biofilms are also resistant to the immune system, making biofilm formation an inherent virulence mechanism. By keeping bacteria closely attached, biofilms also facilitate horizontal gene transfer (HGT) (discussed next). Moreover, many of these drugs also induce the formation of biofilms themselves.
A bacterial cell that does not carry the genes for a full-resistance phenotype, such as those encoding antibiotic-inactivating enzymes or altered cellular targets could bear other changes that slightly increase its MIC (Fig. 1). In many cases, this reduced susceptibility does not result in clinical failure, but can facilitate the emergence of such a fully resistant variety. This idea is more consistent with classic Darwinian evolution than the sudden emergence of a fully resistant organism. A number of genes and physiological responses have been implicated in this low-level resistance: Plasmid-mediated resistance to fluoroquinolones increases the MIC by 20-fold, 143 but the MIC remains below the resistance breakpoints; responses to oxidative stress that diminish the accumulation of xenobiotics, including antibiotics, within the bacterial cell also reduce bacterial susceptibility. 45 but not enough to surpass the resistance breakpoints; and exposure to bile up-regulates genes that confer resistance to polymyxin in enterohemorrhagic E. coli. 97 Many mutations have been identified as factors that reduce the susceptibility of E. coli to different antibiotics, which accumulate, yielding a full-resistance phenotype. 67 In many clinical conditions, such as the dramatically high concentrations of some antibiotics in the urine, these small changes are irrelevant. However, in other scenarios, such as in tissues not reached by antibiotics of poor distribution (or a growing variety of intracellular infections, where the infecting bacteria are not reached by inhibitory concentrations of antibiotics and can furthermore efficiently exchange genes 34 ) or when using substandard drugs 125 or inadequate dosing (adjusting antibiotic doses for bodyweight in adults has been proposed to not only increase the clinical success of treatments, 53 but can also optimize antibiotic exposure, thereby reducing the emergence of resistance), this reduced susceptibility might provide just enough protection for bacteria to survive and gain traits to develop a full-resistance phenotype. 73 This might be particularly true for bacteria in the open environment, where subinhibitory concentrations of antibiotics might select for low-level resistance 7 ; these subinhibitory concentrations may lead to full-resistance phenotypes, due to oxidative-induced mutagenesis,91,148 and could induce the expression of virulence mechanisms. 107 However, some paradoxical effects should also be considered: Very high concentrations of beta-lactams, for instance, can diminish its bactericidal effect and lead to clinical failure, a phenomenon known for more than 50 years as the “Eagle effect.” 118
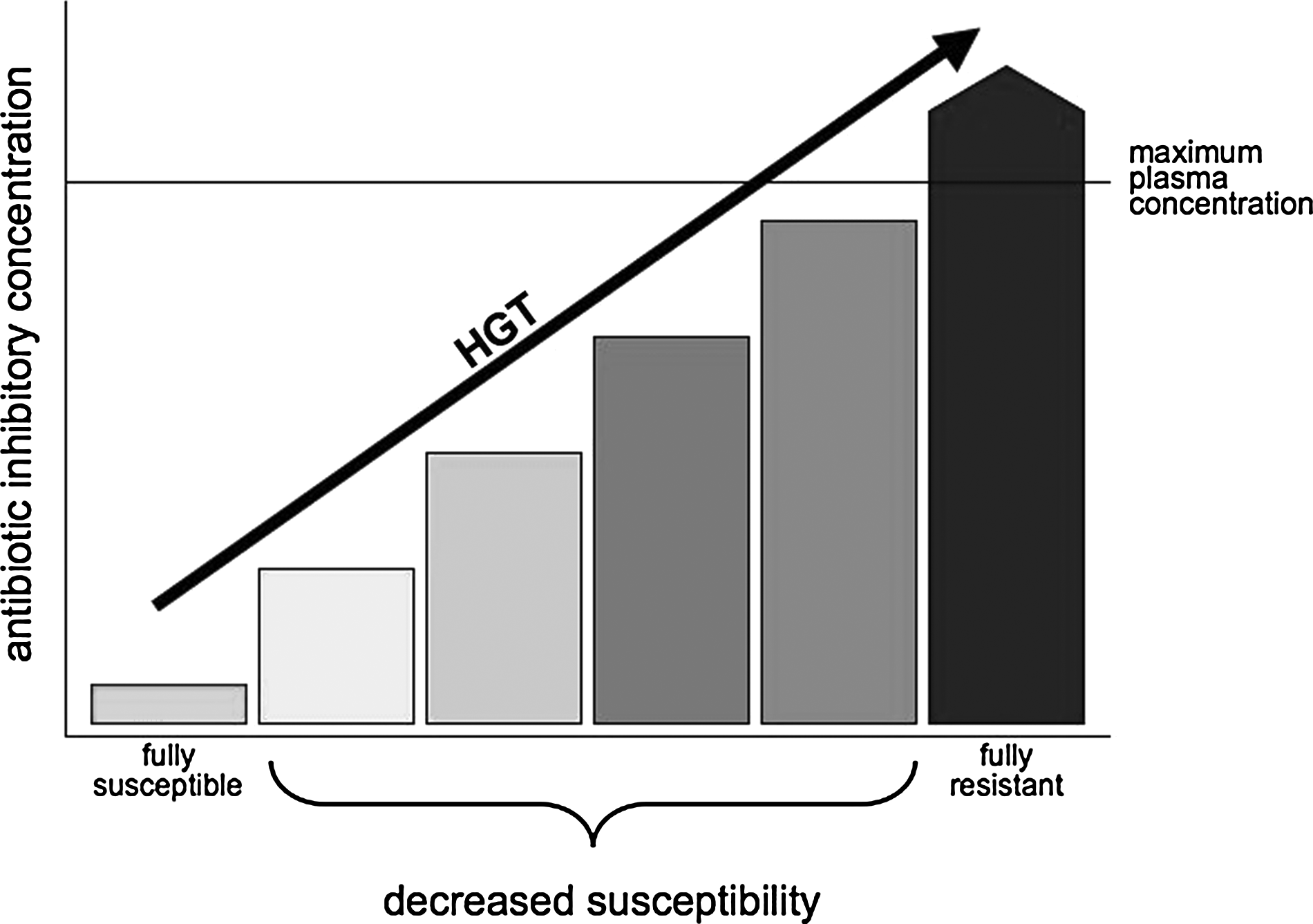
Ladder from susceptibility to resistance. The consideration of bacteria as either, fully susceptible or fully resistant, misses a variety of intermediate stages. Full resistance can be defined as an increase in the antibiotic concentration required to inhibit or kill a given bacteria, leading to therapeutic failure, usually because such concentrations are higher than those attained in plasma or tissues. At the other end, fully susceptible bacteria are inhibited by very low antibiotic concentrations, often in the range of tenths or hundredths of micrograms per milliliter. The sudden transition from full susceptibility to full resistance occurs, and is often the result of horizontal gene transfer (HGT). In addition, different physiological or genotypical changes can lead to subtle increases in inhibitory concentrations, which are slightly higher than those observed in the full-susceptible phenotype but are not enough to prevent inhibition under most clinical conditions. However, such changes can facilitate further increases, become additive, and diminish the clinical efficacy of antibiotics under conditions such as poor antibiotic distribution, inadequate dosage, or suboptimal drugs.
The first side-effect of these phenomena is that antibiotics do not only select for antibiotic resistance (Fig. 2): The ability to form biofilms or to endure other forms of environmental stress can be coselected through the use of antibiotics. These traits, in turn, can increase bacterial virulence, creating a more dangerous pathogen with diminished susceptibility to antibiotics. 40 A wide variety of traits are involved in both antibiotic resistance and virulence, 114 which can be selected for using antibiotics. These mechanisms aside, many examples of virulence genes are linked to resistance determinants in a single genetic element. 90 The coevolution of resistance and virulence has been implicated in the successful development of some dangerous pathogens in the hospital environment, such as Enterococcus faecium. 60 The coevolution of resistance has also resulted in the notion of “super-bugs,” which are multiresistant, highly virulent, and highly transmissible strains, such as some recent MRSA isolates. 79 The role of antibiotics in selecting for HGT abilities will be discussed in detail next.
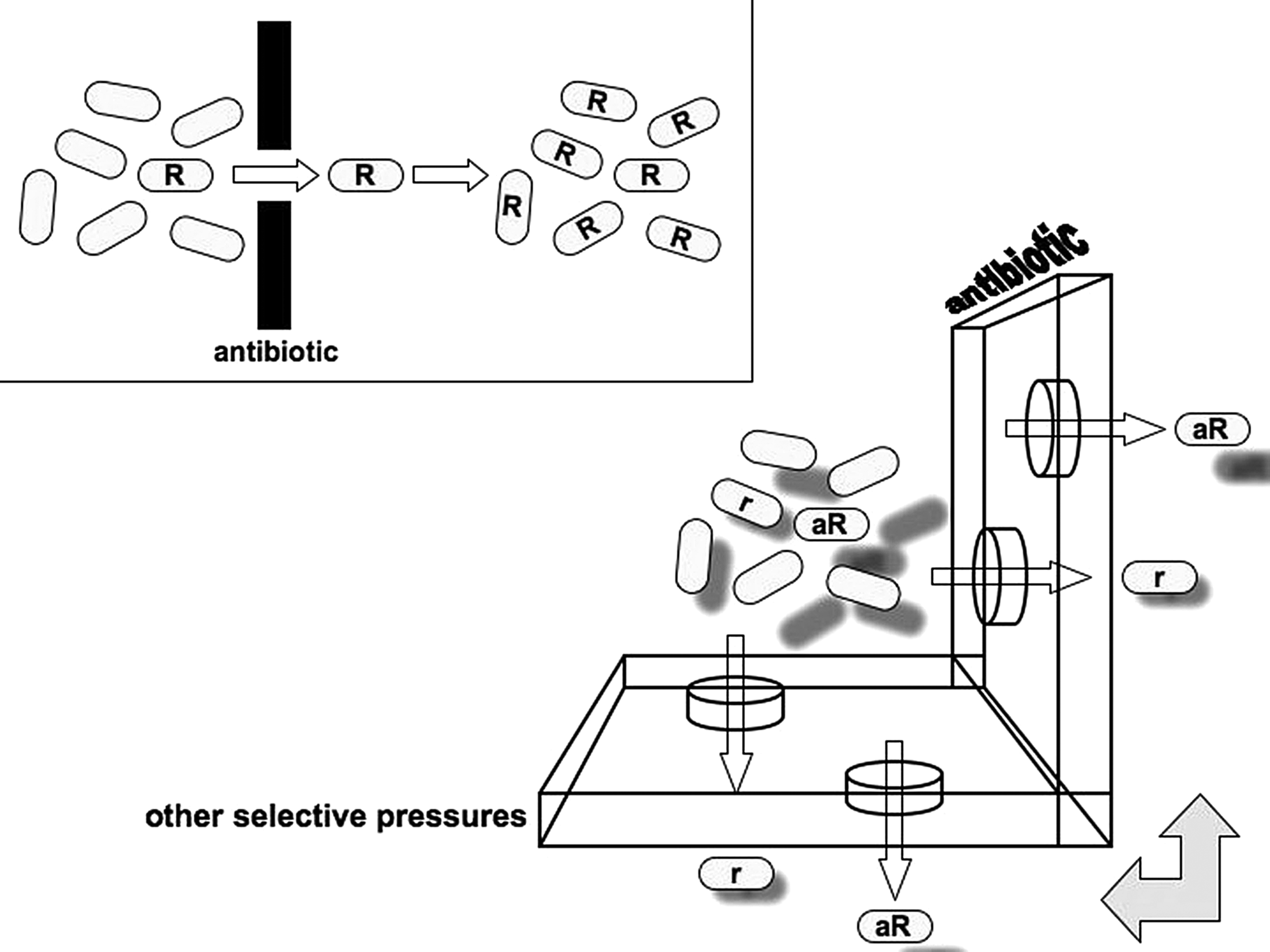
Resistance selection: a 3D view. The simple notion of resistance selection is illustrated in the upper left box: One (the one with an “R”) among many bacterial cells is resistant to a given antibiotic; when the population is exposed to such an agent, only the resistant organism survives, which replicates filling the niche previously occupied by the susceptible population. Instead of this single-dimension view, the process is multidimensional: An antibiotic may be present in different concentrations, even within a single patient, selecting for full resistance (R) or low-level resistance (r) phenotypes; resistance is often the result of additional abilities (a), such as increased gene exchange or stress response traits, which are coselected by antibiotics. Nonantibiotic compounds can also select for antibiotic resistance phenotypes, not only by linkage. Resulting populations multiply and interact, often exchanging genes. Additional layers can be drawn by the use and interaction of different antibiotics and other selective pressures.
Antibiotics serve not only as agents that select for or maintain resistance traits but also as resistance inducers. This effect goes beyond typical examples of resistance genes whose expression is induced by antibiotics (e.g., chromosomal beta-lactamase ampC in enteric bacteria, macrolide-resistance erm, and mef genes in gram-positive cocci 26 ). Antibiotics can induce phenotypes that indirectly foster the ability of bacteria to endure their presence. Exposure to the aminoglycoside tobramycin can induce the formation of biofilms in P. aeruginosa and E. coli 77 ; tetracycline and chloramphenicol can induce the expression of the multiresistance regulon marRAB 42 ; and ribosome inhibitors, such as aminoglycosides and tetracycline, induce the expression of the MexXY efflux pump in P. aeruginosa, which confers multiresistance. 84 Furthermore, several antibiotics can increase the mutability of bacteria, enhancing their chances of obtaining a beneficial mutation during exposure. 113 Fluoroquinolones 15 and beta-lactams 121 can induce the expression of SOS genes (see next), which can protect against the effects of antibiotics and activate other responses associated with the dispersal of resistance genes. Considering the fact that antibiotics are not simply anti-infection drugs but, in many cases, are signaling agents which were produced and used by bacteria long before their discovery, might help in understanding their pleiotropic effects and the risks of exposing bacteria to subinhibitory concentrations during treatment. 107 Antibiotic exposure can also induce the expression of virulence traits. Some of the examples provided earlier, such as biofilm formation and efflux mechanisms, also play a role in bacterial virulence. Fluoroquinolone exposure increases the expression of colonization factors in Clostridium difficile, particularly in antibiotic-resistant strains. 46
A second side-effect is that many other agents and conditions also select for antibiotic resistance (Fig. 2). This notion has been considered for many years, particularly when also considering the linkage issue. Since many antibiotic-resistant genes are present in plasmids that also carry other traits, selective pressures which favor these other traits could coselect for the whole plasmid, including the antibiotic-resistant genes. For instance, mer genes that encode the transporters and enzymes which are necessary to eliminate mercuric compounds are commonly linked to antibiotic-resistant genes; thus, mercury (such as the one released from dental fillings) could select for antibiotic resistance. 156 Genes that confer resistance to disinfectants go along with antibiotic-resistant genes: The qacE gene, which mediates resistance to quaternary-ammonium compounds, is a part of the conserved region of class 1 integrons; and the qrg gene, which mediates a similar phenotype, was recently identified in a Tn916-like element. 30 However, agents or conditions favoring a phenotype that indirectly causes antibiotic resistance, such as exposure to the immune system (toward which biofilm growth can provide protection) or environmental pollutants (such as ozone, which affects the expression of oxidative stress responses 87 ), can potentially coselect for antibiotic resistance. Moreover, complex environments, such as urban areas, particularly those in developing countries, often present agents or conditions fostering resistance. 136 On the other hand, this idea partially explains the occurrence of resistance genes in environments 20 and animals 86 not exposed to antibiotics. Bacteria can use inducible, nonspecific resistance mechanisms to cooperate: Resistant mutants within a population can release indole, which activates stress responses in their nonresistant neighbors, through a mechanism described as “charity,” as the production and release of indole is expensive. 102 Disinfectants have also been implicated as potential inducers or selective agents for antibiotic resistance.2,35 The obsession for aseptic environments has led to a fad for the inclusion of disinfectants in all types of household items, perhaps increasing the prevalence of antibiotic resistance among environmental bacteria. Nonantibiotic drugs that, nevertheless, have antibacterial activity, including psychotropic agents, 93 might also select for unspecific resistance mechanisms. Statins, which are widely used to lower blood cholesterol levels, have been associated with better outcomes of bacterial infections and have, indeed, bactericidal properties that occur at concentrations much higher than those achieved clinically. 19 Celecoxib, a cyclooxygenase-2-selective anti-inflammatory drug, inhibits efflux pumps, thereby increasing the activity of several antibiotics. 89 As many bacterial physiological responses toward environmental stress can also diminish their susceptibility to antibiotics, the list of agents that stress bacteria in such a way might be quite long.
Clinically, all these interactions suggest that the utterly simplistic way we use antibiotics should be reviewed. Apart from pharmacodynamic considerations, it is clear that there are profound differences in how antibiotics select and/or induce resistance, and this information should be used to rationally choose therapeutic options. It is necessary to incorporate our knowledge of resistance mechanisms and gene mobilization to optimize antibiotic treatment. 159 The notion of fixed doses should also be reviewed, 53 to minimize the exposure to subinhibitory concentrations of antibiotics, and elimination pathways should also be considered: For instance, the overuse of oral cephalosporins, which are excreted in sweat, might account for the emergence and rapid spread of community-acquired MRSA 78 ; and the accelerated clearance of levofloxacin in obese individuals may lead to suboptimal plasma concentrations, 32 thus fostering the emergence of antibiotic-resistant mutants. Contrary to “common knowledge,” even obtaining a favorable clinical outcome when using antibiotics to treat infections does not prevent or diminish the risk of resistance. In fact, it might have the opposite effect: For example, a successful antibiotic treatment could wipe out both the pathogen and a significant portion of the microbiota, increasing the risk of colonization by new, antibiotic-resistant bacteria. 108 Similarly, the use of antibiotic combinations with synergic effects, which is obviously preferred for clinical reasons, might potentially select resistance more likely than combining antagonic antibiotics, where resistance to one drug could increase the susceptibility to the other. 111
HGT and Antibiotic Resistance
Although the gradual, low-level, noncanonical mechanisms described earlier play a relevant role in the emergence and evolution of antibiotic resistance, dramatic phenotypic and genotypic jumps, from full susceptibility to full resistance, frequently occur. The most important cause of these jumps is the acquisition of resistance determinants through HGT. A comprehensive review of known acquired resistance genes was recently published. 149 Bacteria that colonize humans, either symbiotically or parasitically, belong to a HGT network in which gene exchange occurs ∼25 times more frequently than in other environments 141 ; therefore, it is not surprising that traits selected through human activities, such as antibiotic resistance, are readily mobilized within this network.
Three main mechanisms of HGT in bacteria have been recognized for decades: conjugation, transformation, and transduction (Fig. 3). Joshua Lederberg played a key role in discovering two of them. While transduction is regarded as a rather “accidental” process and transformation provides a substrate for DNA repair mechanisms, conjugation is a complex system that mediates, among other things, the propagation of HGT itself. 8 Antibiotic resistance genes are among the operational genes considered in the “complexity hypothesis” and, thus, are more likely to be horizontally mobilized. 82 Our knowledge of the role of HGT in antibiotic resistance spread has increased exponentially, and it is also possible that the role of HGT itself has enlarged during the era of antibiotics in yet another case where “the ability to evolve may itself evolve.” 23 Many antibiotic resistance genes are likely originated from antibiotic-producing bacteria that carry a huge variety of resistance genes, including genes conferring resistance to synthetic drugs, 37 a phenomenon which was discovered nearly 40 years ago, on a biochemical rather than on a molecular basis, 17 and that continues to be re-discovered every once in a while. 36 Genes that confer resistance to natural (e.g., aminoglycosides, chloramphenicol, and tetracycline) and synthetic (e.g., trimethoprim) antibiotics have been identified from soil metagenomic DNA libraries expressed in an E. coli host. 117 In any case, the extent of old and recent HGT events and their contributions to bacterial evolution is immense. 39 Recently discovered mobile elements and physiological responses that modify HGT interact with antibiotic resistance determinants in a number of ways; some of these will be reviewed in the next few paragraphs.
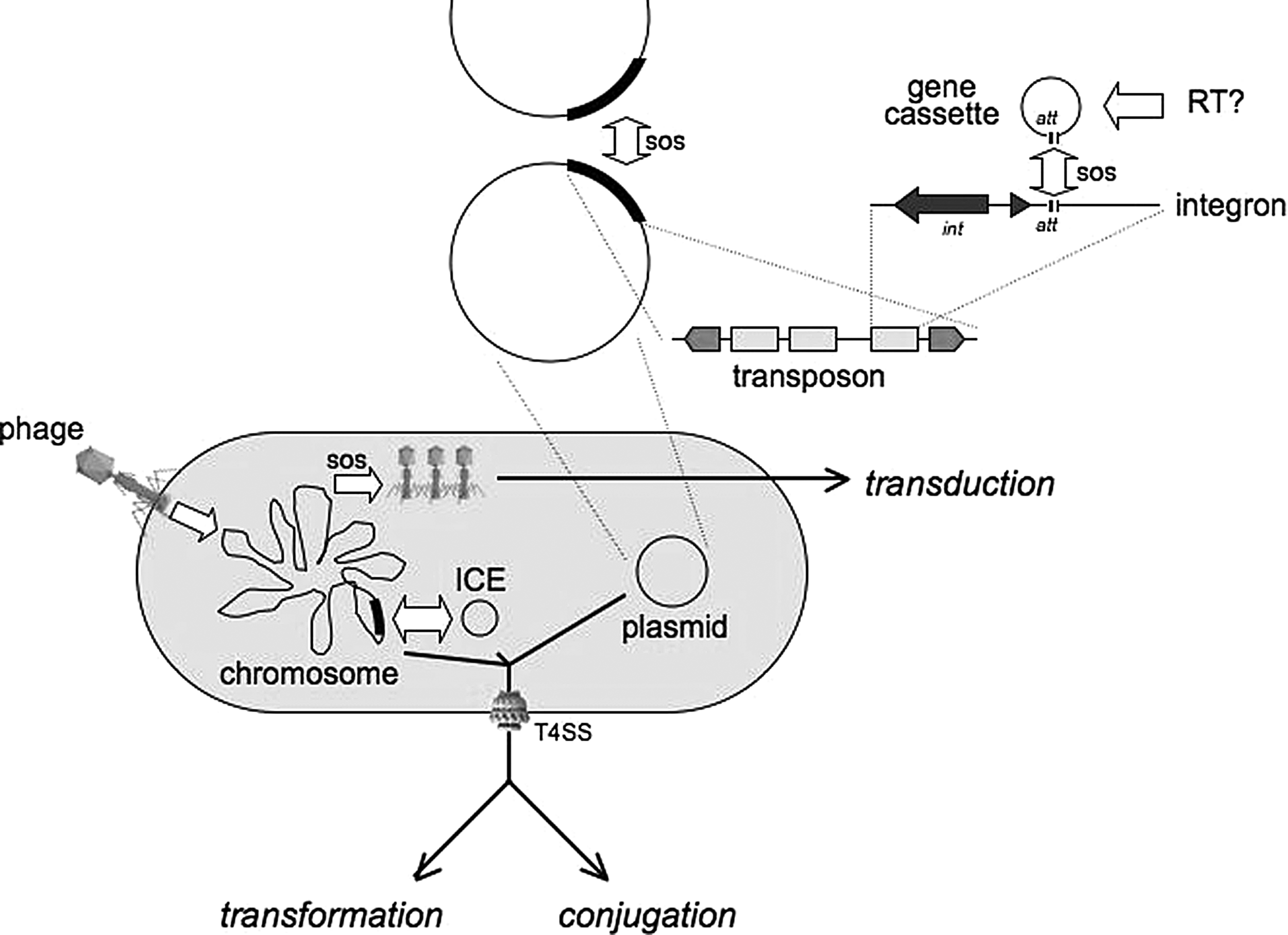
HGT players. The elements and events involved in HGT include plasmids as important vectors in inter-cellular and inter-molecular mobilization; plasmids act as collections of transposons, which, in turn, might bear integrons containing one or several gene cassettes. Gene cassettes are possibly originated through retrotranscription; and a key signal that increases cassette and transposon mobilization is the SOS response. In addition to plasmids, integrated chromosomal elements (ICEs) have been identified in a number of bacterial species; they can integrate into the chromosome and are excised before conjugal transfer. Type IV secretion systems (T4SS) play a role in the inter-cellular mobilization of DNA, either through transformation (both in secreting and in uptaking DNA) or through conjugation. Phages mediate transduction, which is also activated by SOS responses.
Transduction is, perhaps, the HGT mechanism that is least considered for antibiotic resistance. Although phages are very common in the environment (up to 2.5×108/mL in seawater, for instance 18 ), the abundance of these viruses within our bodies was only recently revealed: Sequences of ∼1,000 different phages were found in DNA extracted from human feces. 134 Therefore, the likelihood of these phages to mediate HGT between bacteria from our own microbiota seems high. Phages transfer other clinically relevant genes, from the textbook story of diphtheria or cholera 72 toxins, to the intergeneric transfer of staphylococcal toxin genes, to Listeria monocytogenes. 27 However, there is little evidence for the phage-mediated transfer of antibiotic resistance genes. A recent report of beta-lactamase and methicillin-resistant genes in phage DNA samples from the fecal material of farm animals 31 proves that transduction might indeed play a role in the spread of antibiotic-resistant genes. Furthermore, streptococcal prophages are induced by antibiotics that are typically fed to farm animals, and resistance genes can be detected in these viromes. 4 This issue is particularly worrisome, as phages are now conceived as agents to be used in the fight against antibiotic-resistant infections; the massive release of potential vectors of resistance determinants could further increase the gene flux that mobilizes resistance and virulence traits. However, the phage genomes identified to date contain minute proportions of host genes, 92 indicating that transduction is indeed an accidental phenomenon.
The very discovery of transformation was associated to clinically relevant traits, as a virulence gene was mobilized between pneumococcal strains in Griffith's experiments, which caught the attention of Avery's team. These early experiments showed that this kind of HGT occurs within living animals. However, due to the homology restrictions of most transformation processes, this mechanism has been associated more with gene repair than with gene exchange, 43 although naturally occurring plasmid transformation and the acquisition of large DNA fragments flanked by regions of high sequence similarity have been reported. 146 Furthermore, only a handful of clinically relevant bacteria, such as Streptococcus, Haemophilus, Acinetobacter, and some Pseudomonas, are known to naturally achieve transformation competence. However, at least one important antibiotic-resistant trait has been spread through transformation: penicillin resistance in Streptococcus pneumoniae. Transformation also provided the first known example of horizontally acquired fluoroquinolone full resistance. Resistance to fluoroquinolones is typically mediated through mutations in the target genes, gyr and/or par, that encode topoisomerases; these mutations are recessive, hence simply acquiring the mutated gene through HGT would not confer a resistance phenotype. 57 However, the acquisition of genes through transformation is usually followed by substitution by recombination; therefore, mutant target genes can actually confer fluoroquinolone resistance. 13 Transformation is also thought to be limited by the short life of naked DNA in an open environment, as transforming DNA is either rapidly taken up or degraded after its release. However, a phenomenon recently described in Acinetobacter baumannii might circumvent such limitations. This species releases outer membrane vesicles containing DNA that can be transformed into another cell; plasmids containing carbapenemase genes were mobilized through this peculiar mechanism. 138 A somewhat related phenomenon involving membrane “particles” was also recently reported in E. coli. 28 A variety of gram-negative bacteria produce outer membrane vesicles, although these vesicles have primarily been studied for their involvement in virulence. 51 Finally, the notion of the potential availability of resistance genes during therapy for the transformation of pathogenic (or commensal) bacteria, as such genes are present in preparations of antibiotics of natural origin, 154 was dismissed, because there was no evidence of such transfer. 99 However, this idea was perhaps too quickly dismissed, as our ability to detect such phenomena is so low that events of great clinical relevance would have certainly been missed. 76
Years ago, it was suggested that antibodies induced through antipneumococcal vaccines might interfere with the function of the manganese-permease complex, Psa, which, in turn, might promote penicillin tolerance. 127 Interestingly, psa− mutants exhibited reduced transformation efficiencies. However, pneumococcal capsular-polysaccharide antibodies affect the expression of quorum sensing mechanisms and enhance the transformation frequency of S. pneumoniae, 157 which emphasizes the unpredictable effects that anti-infectious efforts, not only antibiotics, can have in the physiology and genetic composition of bacteria.
Since its discovery, bacterial conjugation was considered as being mediated exclusively through plasmids. Conjugation was first demonstrated between F+ and F− strains of E. coli. Subsequently, plasmids that are capable of conjugative transfer between E. coli and Salmonella were called “promiscuous.” Currently, the “molecular moral” is more relaxed and recognizes not only conjugation between distantly related bacteria, but also inter-kingdom transfer: bacteria-archaea, 48 bacteria-yeast, bacteria-plant, 8 and even bacteria-mammal cells. 152 Conjugation also occurs from a dead bacterial donor and among intracellular bacteria.54,153 Bacteria inhibited by bacteriostatic antibiotics can act as static recipients of conjugation, accumulating resistance genes, particularly under cycles of the presence–absence of antibiotics. 155 In the early 1980s, a new form of mobile genetic elements that was capable of mediating conjugation were discovered in enterococci, called conjugative transposons. Some years later, similar elements were identified in the gram-negative anaerobe Bacteroides. Currently, a huge catalog of integrated conjugative elements (ICEs) has been detected from many different bacterial clades. ICEs mainly exist as a part of the host cell chromosome, into which they are integrated on arrival through the activity of recombinases encoded within the ICE itself; and can excise as a circular molecule that is transferred conjugatively. Some ICEs can recombine, forming hybrids that are segregated through conjugation, containing new combinations of antibiotic-resistant genes. 63 ICEs mediate a number of antibiotic-resistant and virulence phenotypes, 96 and they are also capable of mobilizing coresident plasmids, even those that are considered nonmobilizable (i.e., lacking mobilization-oriT functions). 101
Type IV secretion systems (T4SS) crosslink several instances of HGT with virulence: the mobilization of DNA from donor to recipient cells during conjugation, either between bacteria or from bacteria to plant cells; the uptake of DNA, leading to transformation, and even the release of DNA into the extracellular milieu to facilitate this kind of genetic exchange; and the translocation of several proteins into eukaryotic cells that mediate the virulence of bacteria as diverse as Helicobacter pylori, Neisseria gonorrhoeae, and Legionella pneumophila. 56 The crucial role of T4SS as a translocator of the genes that encode the T4SS themselves, as a part of the conjugation machinery, and of other genetic information and proteins involved in resistance and virulence, makes T4SS a target for future anti-infection approaches.
Gene exchange within biofilms has been recently explored, as bacteria in environmental and clinical conditions commonly form biofilms. The close contact of bacteria in biofilms is believed to foster horizontal transfer, particularly through conjugation. Experiments using plasmid-encoded fluorescent proteins that are only expressed in recipient cells elegantly allowed the assessment of the mobilization of conjugative plasmids in biofilms. 71 Conjugative plasmids themselves carry determinants that induce bacteria to form biofilms, perhaps fostering conjugative transfer. 65 The results of recent studies with IncP-1 plasmids have shown that this transfer mainly occurs at air-liquid interfaces in biofilms and that oxygen availability influences the extent of conjugative transfer. 94 However, the impact of biofilm-forming ability on the acquisition of plasmids was not evident when comparing biofilm-producing with nonproducing clinical isolates of P. aeruginosa (although biofilm-producing strains carried more antibiotic resistance traits 44 ). Transformation can also be enhanced in biofilm environments. Pheromones that govern genetic competence in Streptococcus mutans work better when these bacteria are growing in biofilms. 106 Extracellular DNA in biofilms might also be a source of DNA for transformation. 147 Other environmental conditions that foster HGT, apart from biofilms, can be found during infectious processes. As previously discussed, conjugation can occur between intracellular bacteria; and inflammation, elicited by enteropathogens, generates a set of conditions that bring transconjugation efficiencies to nearly 100% among Enterobacteriaceae. 142 Once again, virulence, HGT, and antibiotic resistance overlap in diverse, unpredictable ways.
Inter-cellular gene mobilization has important barriers, mechanisms that can be considered as a sort of “immunity” for parasitic DNA. Restriction enzymes play a role in these defenses, reducing the efficiency of HGT. 146 Clustered, regularly interspaced short palindromic repeats (CRISPR) and associated genes provide protection, on first exposure, to phage infection, and also induce plasmid loss. 62 The plasmid host range varies depending on the ability of the plasmid to use the DNA-replication machinery of different host bacteria for self-replication. Under antibiotic stress, HGT presents a clear advantage for bacteria. It is conceivable that antibiotics select for more efficient HGT, either as bacteria devoid of restriction and other “immunity” or as mobile elements capable of circumventing such barriers. Plasmids with a wide host range, such as those of the incompatibility groups IncN, IncP, and IncW, 145 might become more prevalent among antibiotic-exposed bacteria (there have been more and more cases of epidemic plasmids115,128); while inter-molecular mobile elements might gain inter-cellular mobility in plasmids with a wide host-range transfer, irrespective of the long-term stability of such plasmids. Mutations in restriction pathways occur naturally in S. aureus, facilitating the highly efficient transfer of vancomycin resistance from Enterococcus faecalis. 144 These changes might not be present in whole populations of bacteria, as maintaining mobile elements and HGT mechanisms in the absence of direct selective pressure could be expensive. However, subpopulations with enhanced HGT abilities, such as self-destructive cooperation, 1 might maintain these trait should the need arise.
By the 1970s, plasmids were found to act as collections of transposons, many of which included antibiotic resistance genes. Gathering resistance transposons in a single plasmid was soon identified as a primary evolutionary engine of multiresistance. During the 1980s, several groups were following the traces of a different type of mobile element, which was finally identified by Ruth Hall, and named integron. Integrons are “assembly platforms” that gather, continuously rearrange, and ensure the expression of small mobile elements called “gene cassettes,” which contain genes for virulence (particularly the super-integrons of Vibrio cholerae) and for antibiotic resistance. 116 Integrons played a role in the early outbreaks of resistant Shigella in the 1950s 41 and are now abundant among enteric bacteria in clinical settings, yet they seem to have emerged in other, unrelated organisms and perhaps were only maintained in enterics because of antibiotic pressure. 47 Gene cassettes have their own, independent, and surprising origin: They seem to have emerged from the reverse transcription of mRNA molecules. 135 Gene cassettes are contained within integrons that are commonly located within transposons, such as Tn21, and transposons mobilize into plasmids, resulting in a nested or “matrioshka” effect (Fig. 3). 8 Class 1 integrons, the most common ones among clinical isolates, and frequently found in mobile genetic elements, were most likely derived from a chromosomal precursor that was captured by a Tn402-like transposon. 139 Gram-positive bacteria, that do carry integrons, 124 have their own gene rearrangement machineries: The insertion sequence IS257, which flanks a wide variety of antibiotic-resistance genes in staphylococci, including the mec gene for methicillin resistance, is crucial for the evolution of multiresistance in this gram-positive pathogen. 55
Although there are many different physiological conditions that regulate inter-molecular and inter-cellular gene mobilization, one condition is common to many of these mechanisms: DNA damage. The transition from the lysogenic to the lytic phase of some bacteriophages is regulated through the SOS response (a set of cellular activities triggered by DNA damage, usually revealed by the presence of single-stranded DNA), as is the expression of the transposase enzyme of some transposons 5 potentially increasing the rate of transduction and transposition, respectively, in response to DNA damage. Recently, the intricacies of the DNA damage response, HGT, and antibiotic exposure have increased. As previously discussed, fluoroquinolones and beta-lactams induce the SOS response and, consequently, several instances of HGT. In the particular case of V. cholerae, other antibiotics, such as tetracycline, chloramphenicol, and aminoglycosides, are also capable of inducing SOS. 11 The expression of the integrase gene of integrons is increased 4.5-fold in E. coli and 37-fold in V. cholerae through the SOS response; this induction can foster the acquisition and, most importantly, the rearrangement of gene cassettes, so that the necessary ones end up closer to the integron promoter, ensuring the expression of relevant resistance phenotypes. 69 The momentary presence of single-stranded DNA, just after conjugation, in the recipient cell, induces the SOS response and, with it, the expression of integrases and rearrangements in integron cassettes. 10 In Streptococcus gordonii, hydrogen peroxide-induced DNA damage induces the release of extracellular DNA, along with transformation competence; the released DNA promotes biofilm formation and is long enough to mobilize antibiotic resistance genes. 81 The notion that the therapeutic activity of bactericidal antibiotics is mediated through the generation of oxygen radicals 91 might also explain the ease with which resistance determinants mobilize. In the end, antibiotics, and many other chemical agents, induce SOS responses that directly protect against such agents or increase the mobility and rearrangements that are necessary to optimize the use of the resistance gene pool.
A peculiar exception to the role of HGT is observed, paradoxically, in the most successful human pathogen, Mycobacterium tuberculosis. This organism is thought to infect one-third of humans, with more than 9 million new cases of tuberculosis and nearly 2 million deaths each year. 132 Multiresistance is a common and increasing trend. Multidrug-resistant tuberculosis (MDR-TB) is caused by organisms at least resistant to isoniazide and rifampicin, and extensively drug-resistant cases (XDR-TB) are also resistant to a fluoroquinolone and an injectable drug, such as capreomycin, kanamycin, or amikacin. 61 Although the very prevalence of these varieties has been difficult to assess, due to the lack of resources typical of the most affected countries (most MDR-TB cases develop in India, China, and Russia), it is considered that nearly 2 million people are affected by MDR-TB, of which 2%–20% are XDR, depending on the country. 123 However, M. tuberculosis seems to be completely apart from the gene flux that is common for almost all other bacterial pathogens, as there is no evidence of recently acquired genes or mobile genetic elements in this species. Moreover, contrary to the commonly held belief that M. tuberculosis was acquired from domesticated animals, it now seems likely that the opposite is true. 59 One can argue that, having reached such a successful status as a pathogen, and given the physiological and population-dynamics conditions that make it possible to gain resistance through mutations, 112 HGT is much more of a burden than an advantage for this bacterial species. However, M. tuberculosis was not always like that: A M. prototuberculosis apparently gathered pathogenicity islands from gamma proteobacteria, before the events that led to the clonal-like evolution of the current M. tuberculosis. 137 Close relatives of M. tuberculosis, such as M. smegmatis, carry mobile genetic elements, and conjugate in biofilm environments 126 ; there is also evidence of HGT in M. canettii. 59
Although “non-canonical” mechanisms that diminish antibiotic susceptibility bring the evolution of resistance more into a Darwinian model, as discussed earlier, HGT is perhaps the biggest mediator of antibiotic resistance. Mobile genetic elements and HGT are often considered small-scale equivalents of what bacteria and contagion mechanisms are to humans: Plasmids and other elements act more similar to parasites to bacteria, and HGT are just the means for them to spread, despite the efforts of bacteria to restrain this mobilization. However, this view is at the very least incomplete, in the same way as it would be to consider all bacteria that colonize us as parasitic. The relationships between bacteria and mobile elements range from parasitic, wherein bacteria are utilized as vehicles for the propagation of mobile elements, to symbiotic, wherein bacteria and plasmids coevolved into associations that increased their mutual fitness. Thus, antibiotics serve as external “post-segregational killing” mechanisms (plasmid-mediated systems, such as codified toxin-antitoxin pairs that kill bacteria which spontaneously lose their plasmids) that prevent the loss of plasmids and other mobile elements, 75 perhaps even forcing the eventual coevolution of both cellular and subcellular forms of life.
New Antibiotics: Do we Really Need Them? How Can we Obtain Them?
It is generally accepted that we urgently need new antibiotics. 25 However, there are diverging opinions in this regard. I would surmise that we need new drugs, as bacteria are increasing their resistance at a formidable pace, due to the no less formidable array of mechanisms discussed earlier, and as infectious diseases are slowly re-gaining their lethality. Recently, a rather imprecise initiative to obtain 10 new antibiotics by 2020 (10×20 initiative 80 ) has been proposed. The WHO developed a “policy package” that recognizes the burden of antibiotic resistance, but is rather a set of statements. 103 Furthermore, we need agents that are completely new, not just new generations of old drugs: It takes some years to modify an old antibiotic backbone to provide a slight advantage and to have it licensed for clinical use, whereas much less time is required for bacteria to modify old mechanisms to broaden its resistance spectrum, as was observed with the rapid emergence of tens of extended-spectrum beta-lactamases. Since 2000, only three antibacterial drugs have been developed from a new antibiotic class (including daptomycin, a lipopeptide similar to colistin, and retapamulin, a pleuromutilin derivative, similar to other drugs previously used in animal health). From among the 39 compounds under clinical evaluation, including topic disinfectants, only 10 are considered to belong to novel antibiotic classes. 24 The pharmaceutical industry has not met the need for new antibiotics, for a number of reasons that will be discussed later (although some researchers argue that this depressed trend is common for all drug classes 122 ). Perhaps a more Keynesian approach is needed to return pharmaceutical research to its tracks.
Large transnational pharmaceutical companies, known as big pharma, perceive antibiotics as a bad investment, despite the fact that the global antibiotic market was estimated at U.S.$25 billon 10 years ago, 151 and thus abandoned R&D in the field for many years. 131 From 20 large pharmaceutical companies that were conducting antibiotic R&D in 1990, fewer than 5 are currently engaged in antibiotic research. On the one hand, there are plenty of antibiotics, making for a strong competition, and most antibiotics are cheap, except for those devoted to particular multiresistant organisms, which will never be blockbusters. On the other hand, infectious diseases are almost always short-lived ailments that require short-term treatments, rarely exceeding a week (along with a trend to shorten the duration of treatments even further 64 ). Furthermore, there are legions of experts continuously stating that the use of antibiotics should be curbed to slow the pace of resistance (although curbing antibiotic use not necessarily reduces resistance 52 ). When new antibiotics, or new versions of old ones, arise, they are often considered as “last resort” drugs during most of the patent life of the drug because of the fear of developing resistance. Thus, the marketing scenario is rather grim: Total antibiotic sales have fallen by 1.6% annually since 2005, and this trend will continue or worsen through 2019 in the top seven markets (United States, Japan, France, Germany, Italy, Spain, and the United Kingdom). Since financial gain is the only reason for big pharma to conduct R&D, and societies have essentially placed the responsibility for all drug R&D in the hands of big pharma, we are facing a terrible problem. Since public overtaking of antibiotic R&D as a matter of public interest seems to be contrary to our capitalistic soul (a recent effort in the United States to finance drug research with public funds is ridiculously poor, and eventually all promising molecules will be transferred to big pharma), an awkward new term is emerging in this discussion: “incentives.” The argument goes similar to this: Since antibiotics are bad business, societies should provide incentives for big pharma to conduct R&D in this field, such as extended patent protection, “wild card” patents, or increased prices. 130 In the United States, the Generating Antibiotic Incentives Now (GAIN) Act is aimed at changing patent protection and the approval of antibiotics. 100 Societies are, therefore, held hostage: If they want new antibiotics, a ransom must be paid. However, drug companies are very wealthy, and the development of new antibiotics is not going to lead to financial losses but rather will result in underwhelming profits compared with the profit from drugs for the management of diabetes or erectile dysfunction. Why not conduct antibiotic research just because it is the right thing to do, and in the interest of human health, as their mottos claim? Perhaps it is time to realize that, as a society, we are facing a grave health crisis, and that thinking outside the free market box is required to solve this problem. The sole notion of “incentives” is already contrary to free market forces; therefore, the insufficiency of these forces to drive antibiotic R&D is inherently accepted when proposing incentives. However, instead of “positive” incentives, societies and governments might need to force big pharma to develop new antibiotics. For example, if drug companies do not seriously commit to this effort, severe restrictions on promotion and marketing strategies and price limitations for their products in other therapeutic areas could be issued. Companies that do not engage in antibiotic R&D might face reduced patent protection for their new products, and simple variants of older drugs, that is, the “me too” drugs, could be denied licensing. The very concept of patent protection for antibiotics, that might even be driving resistance, 74 should be analyzed, and new protection instruments should be developed. At the same time, not-for-profit institutions should recover their leading role in developing at least these types of drugs. 33 The current era of ample leeway for big pharma should end, 110 as it has not delivered the products that societies need most.
Either positive (carrot) or negative (stick) incentives are interventions beyond free market forces. Apart from the fact that the pharmaceutical industry has not delivered new antibiotics in many years, Laxminarayan 100 presents three reasons for the need of interventions that are aimed at innovation, from the economic analysis of the current situation: (1) while broad-spectrum antibiotics are likely to generate greater revenue, narrow-spectrum antibiotics have greater societal value, as they create less selective pressure for resistance; (2) market forces are against the rational use of antibiotics, pressing for usage in the management of self-limited and even nonbacterial infections; on the other hand, resistance, while being against the interest of public health, is good for business, particularly for companies selling antibiotics; and (3) although it is clear that new antibiotic classes (or mechanisms of action) are needed, and that the diversity of antibiotics reduces the selective pressure on resistance, it is easier to just modify existing drugs that would only provide a marketing advantage instead of developing real solutions to face growing resistance. Other interventions curbing antibiotic usage might also be needed, and not just those that are aimed at re-starting antibiotic R&D. 100
Another example for the inadequacy of the free-market approach to this issue was provided by the recent NDM-1 scare. Developing countries, such as India, in which antibiotic abuse, low-quality drugs, and other problems increase the prevalence of resistance, are a source of resistance outbreaks in developed countries, due to the unexpected medical tourism trend. Rich countries, in which all big pharma companies reside, might feel that it is only important to ensure their own supply of antibiotics, leaving poor countries on their own, according to free-market doctrines. Of the 15 new antibiotics in clinical trials or awaiting approval, only six have proved or assumed activity against gram-negative bacteria 50 (MRSA and enterococci are the perceived immediate threats in developed countries). However, globalization is also a clear trend for infectious diseases that can bring the most unexpected super-bugs to developed countries. A final, albeit politically incorrect, comment regarding this issue: the large majority of ailments for which big pharma is actively pursuing new drugs are the consequence of chosen life styles (e.g., poor diets, smoking) or of our artificial desire to live much beyond the lifespan of our biochemistry and physiology. Infections, in contrast, can potentially affect any of us, and there is little a person can do to prevent the acquisition of most infectious diseases. Therefore, R&D for new antibiotics and vaccines should be strongly supported and actively pursued by governments instead of being subjected to market issues. Individuals who are not willing to abandon their lifestyles or who wish to extend their lives by a few months at an enormous expense should finance these goals themselves as a consequence of their decisions, thereby releasing massive public funds that can then be used for obtaining drugs which all individuals will inevitably need.
Another set of financial interests might be fostering resistance: those involving generic antibiotics. While new drugs should be clinically tested for safety and efficacy, generic drugs should only prove bioequivalence in a small set of healthy volunteers. The financial interests of governments and insurance companies to have cheap drugs push for maintaining low regulatory standards for generic drugs. Only a handful of studies compare generic and “original” antibiotics in terms of purity88,98; a single report shows that generic, bioequivalent vancomycin is inferior to the original one in in vivo experiments, 150 and the emergence of generic ciprofloxacin was followed by a 200% increase in ciprofloxacin resistance, although this effect was attributed to increased usage (0.13–0.33 DDD/1,000 inhabitants-days) due to the lower cost of the generic drug. 85 Unfortunately, studies aimed at assessing the clinical efficacy of generic drugs, as well as their impact on resistance prevalence, are expensive and are no longer of interest to big pharma companies. Hence, societies are held between a rock and a hard place, because the development of new antibiotics requires “incentives,” original drugs are expensive, and generic drugs are not forced to demonstrate clinical efficacy. Pharmaceutical and insurance companies, as well as government agencies, preoccupied with short-term savings, are the only winners in this scenario. While healthcare budgets are limited and the allocation of expenses is decided using cost-benefit analyses, quality-adjusted life year, and other perverse bean-counting strategies, the monies for wars, luxurious clothing, travel and accommodations for endless summits, and other extravagant expenses seem to be limitless.
R&D of New Antibiotics and of New Ways to Fight Infection
Although the search for new antibiotics is trying to evolve, from traditional strategies, testing compounds against cultures, toward new approaches that include genomic information and combinatorial chemistry, we are yet to obtain a single drug derived from these methods. 140 The identification of potential targets through an analysis of the genomes of pathogenic bacteria has sparked some controversy. Furthermore, at least one intriguing set of results questions the very availability of many more new targets that can be used in the fight against infection. 16 While developing new antibiotics, it may be worthwhile to also assess the potential for selecting resistance phenotypes; some assays have been developed for this purpose. 70 Since most antibiotics are prescribed empirically, it may also be useful to develop assays that optimize antibiotic usage, such as the measurement of procalcitonin. 22 Quick and cheap methods for assessing, in the medical office, the etiology of simple diseases, such as otitis or tonsillitis, might save millions of antibiotic prescriptions that are currently wasted against viral illnesses and which foster resistance.
Apart from “traditional” antibiotics, that is, drugs that selectively kill or inhibit the growth of bacteria, a wide variety of approaches are currently being explored to enter the “post-antibiotic era.” Some ideas include the following: (1) adjuvant agents that protect or reinforce the effect of typical antibiotics, such as efflux pump inhibitors 29 or plasmid eliminating agents 9 ; (2) the therapeutic use of bacteriophages 119 as a means of “biological control” of bacterial populations (with the grave potential backfire, mentioned earlier, of introducing HGT vectors as therapeutic means) or even as plasmid eliminating agents 83 ; and (3) agents that inhibit virulence traits 3 rather than directly killing or inhibiting bacterial growth. Despite their appeal, these options face biological or financial obstacles that make them nearly nonviable, at least under our current models of drug development. For instance, most antiresistance and antivirulence drugs will be of a very narrow spectrum and will require entirely new clinical laboratory assays to assess their usefulness, which are very different from the typical antibiogram. Furthermore, narrow-spectrum agents will also have a very narrow marketing niche, generating slim revenue; this problem might also be hindering the development of antineoplastic agents that are effective against a very particular kind of tumours. 6 The way we do R&D for new antibiotics, focusing primarily on financial objectives, is likely limiting the potential antiinfectious strategies and fostering the rapid emergence of resistance 74 ; but this suggestion was viewed and discarded as a pro-Soviet argument, 109 showing how difficult it would be to modify the current ways to discover new antibiotics.
Final Considerations
Antibiotic resistance, instead of causing a dramatic, movie-like, worldwide outbreak, is slowly decreasing our ability to harness infectious diseases, increasing the morbidity and mortality of infections at a slow but constant pace. In many ways, our awareness of these circumstances mimics that of our awareness of weight gain, which occurs so slowly that it goes unnoticed, except when we need new clothes, experience difficulties in simple tasks, and face a new set of diseases. This gradual progression masks the catastrophic scenario that underlies both antibiotic resistance and weight gain and suggests that only slight behavior modifications are needed. Instead, a radically new approach is required to face the grim reality of the increased number of antibiotic-resistant bacteria and the decreasing effectiveness of the current arsenal of antibiotics. We moved at full speed into the antibiotic era, using these drugs before a complete understanding of the biology of bacteria was achieved. The consequences are the same as driving a car rapidly, at night, without headlamps. Although there is still much to learn about bacteria, the transition to a “post-antibiotic era” might be achieved through the simple application of our current knowledge of bacterial biology, and to stop thinking of health as a merchandise. It is no longer time for low-calorie sweeteners and a few weeks of soft exercise.
Footnotes
Acknowledgment
The author thanks Jack Heinemann for his comments on an early version of this article.
Disclosure Statement
The author has received honoraria as speaker and/or participant on advisory boards regarding antibiotics, from Abbott, Bayer, Boehringer-Ingelheim, Roche, Stendhal, and Wyeth (now Pfizer); and conducted research sponsored by Bayer, Boehringer-Ingelheim, Stendhal, Pfizer, Schering-Plough (now MSD), and Wyeth (now Pfizer).