
Abstract
The objectives of this study were to compare the in vitro adhesion and invasion of human epithelial cells, motility, and toxin production characteristics of Campylobacter-susceptible strains and their fluoroquinolone- or macrolide-resistant mutants. Susceptible strains and resistant mutants demonstrated similar adhesion capacities to epithelial cells. For Campylobacter coli, fluoroquinolone-resistant mutants with Thr86Ile or Asp90Asn substitutions showed a higher rate of invasion of Caco-2 cells than their isogenic parental strain. Fluoroquinolone resistance did not impact C. coli motility. Mutants harboring Asp90Asn had greater cytotoxic activity than the parental strain. Macrolide resistance had no impact on the studied characteristics of C. coli. For Campylobacter jejuni, fluoroquinolone-resistant mutants had slightly different invasiveness levels and significantly lower motility than the isogenic parental strain. C. jejuni macrolide-resistant mutants with A2074G substitution in the 23S rRNA gene had a higher invasiveness level than its parental strain, but mutants with A2074C in 23S rRNA and G221A in rplD showed reduced motility and similar invasion levels to the susceptible strains. Neither fluoroquinolone nor macrolide resistance appears to affect C. jejuni cytotoxicity. In conclusion, mutations that are frequently encountered in Campylobacter-resistant strains can enhance the invasiveness in Caco-2 cells.
Introduction
Fluoroquinolones act by interfering with DNA gyrase that catalyzes the negative supercoiling of double-stranded DNA. Mutations in the quinolone-resisting determining region (QRDR) of the gyrA gene cause resistance to fluoroquinolones by altering the amino acid sequence near the putative active site of the GyrA subunit. In Campylobacter, specific mutations in GyrA such as Thr86Ile, Asp90Asn, and Asp90His were linked to fluoroquinolone resistance. 1
Macrolides bind to the 50S bacterial ribosomal subunit and inhibit the elongation of peptide chains by interfering with protein synthesis. In Campylobacter, chromosomal mutations, primarily 23S rRNA point mutations (e.g., A2074C, A2074G, and A2075G), account for most macrolide resistance. 19 Recently, it was reported that modifications in the rplD and rplV genes encoding ribosomal proteins L4 and L22, respectively, act in synergy with the CmeABC efflux pump, thereby conferring macrolide resistance to Campylobacter. 7
Previously, we and others showed that antimicrobial resistance could enhance fitness 28 or impose23,24,37 a fitness cost on the in vitro growth, on the persistence on food surfaces, or on the capacity of Campylobacter to colonize chickens. In addition to these studies, we next decided to explore some of the characteristics of Campylobacter, possibly influencing the virulence of this organism. Although the virulence strategies are not fully understood, motility, adherence to, and invasion of epithelial cells and cytotoxin production are thought to play a role in the virulence of Campylobacter 34 and can be studied relatively easily. 22 Thus, we decided to compare these properties in our previously characterized susceptible strains and resistant mutants.
Materials and Methods
Bacterial strains and growth conditions
Strains used in this study were isolated from healthy chicken caeca from a slaughterhouse. They were isolated by culture on Karmali agar plates supplemented with selective agents (Oxoid, Dardilly, France), and identified to species level by multiplex PCR. 12 Strains were routinely cultured in the Mueller-Hinton broth (MHB; Biorad, Marnes la Coquette, France) or on Karmali agar plates at 42°C, under microaerobic conditions (5% O2, 10% CO2, and 85% N2).
In vitro selection of fluoroquinolone- and macrolide-resistant mutants
Fluoroquinolone-resistant mutants harboring the Thr86Ile mutation in their GyrA subunit (Cc103T and Cj126T) were constructed by transformation using a 306-bp fragment of the QRDR of the gyrA sequence from a resistant strain as described previously. 37 The other fluoroquinolone- and macrolide-resistant mutants were selected using ciprofloxacin (Cip; Sigma, St Quentin-Fallavier, France) and erythromycin (Ery; Sigma) as selective agents, respectively, as described by Payot et al. 31 and Caldwell et al. 8 with minor modifications.
Antimicrobial susceptibility testing
The minimal inhibitory concentrations (MICs) of antibiotics were determined for all strains and mutants using the agar dilution method (CLSI, M31-A3). 10 According to EUCAST recommendations, 15 strains with MIC(Cip)>1 mg/L were considered to be resistant to fluoroquinolones. C. coli strains with MIC(Ery)>8 mg/L and C. jejuni strains with MIC(Ery)>4 mg/L were considered to be resistant to macrolides.
Characterization of parent and mutant strains
Parental and mutant strains were characterized by sequencing of the genes implicated in fluoroquinolone resistance (QRDR of the gyrA and gyrB genes) or macrolide resistance (the three copies of domain V of the 23S rRNA, rplD, and rplV genes). Sequencing was performed as previously described.36,37
In vitro cell lines and cell growth conditions
Caco-2 cells obtained from the European Collection of Cell Cultures (09C004) were grown routinely in the Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Cergy-Pontoise, France) supplemented with 10% fetal bovine serum (FBS; Eurobio, Les Ulis, France), 1% of nonessential amino acid (NEAA; Eurobio), penicillin (Sigma) 10,000 UI/ml, and streptomycin (Sigma) 10,000 mg/ml at 37°C in a 5% CO2 humidified atmosphere. For experimental assays, Caco-2 cells were grown in 24-well plastic plates (for adhesion and invasion assays) or in 96-well plastic plates (Dutscher, Brumath, France) (for cytotoxicity assay) seeded at 105 cells per well and incubated for 24 hours (cytotoxicity assay) or 48 hours (adhesion and invasion assays) before infection.
Adhesion and invasion assays
Adhesion and invasion assays of the susceptible C. jejuni and C. coli strains and fluoroquinolone- and macrolide-resistant mutants were performed using Caco-2 cells, with five independent repeats for each experiment. A 24-hour culture of each strain was inoculated in 15 ml of MHB to obtain a final optical density of 0.01 at 600 nm. Cultures were incubated at 42°C with shaking (140 rpm) under microaerobic conditions for 18 hours. Using these cultures, an inoculum of each strain was adjusted to 0.5 McFarland in the DMEM containing 1% FBS and 1% NEAA. About 100 μl (107 CFU/ml) of this suspension was used to inoculate duplicate wells containing a confluent monolayer of Caco-2 cells. The titers of the bacterial inocula of the initial suspensions were quantified by plating 100 μl 1:10 serial dilutions in phosphate-buffered saline (PBS) on Karmali agar plates. Caco-2 cells were incubated for 3 hours at 37°C and 5% CO2 to allow bacterial adherence and invasion. To determine adherence, the cells were washed three times in PBS solution, and then the monolayer was lyzed with 0.1% Triton X-100 (Sigma) for 15 minutes at 37°C, and total bacteria (intracellular and adherent bacteria) were enumerated by serial dilutions as described above. To determine invasion, the monolayer was washed three times in PBS solution, and 1 ml of DMEM containing gentamicin at 100 mg/L was added to kill extracellular bacteria. After 2-hour incubation, the monolayer was washed three times in PBS solution and lyzed with 0. 1% Triton X-100 as described above. After serial dilution in PBS, released intracellular bacteria were enumerated by the colony count method on Karmali agar plates cultured under microaerobic conditions. Invasion ability (Inv%) was expressed as the percentage of the inoculum surviving the gentamicin treatment, and adherent bacteria (Adh%) were expressed as the total number of bacteria counted without antibiotic treatment relative to the initial inoculum.
Motility assay
The motility phenotypes of all susceptible strains and resistant mutants were tested in the Mueller-Hinton medium containing 0.5% agar (Becton Dickinson, Le Pont de Claix, France). Bacterial cells were grown on Karmali agar plates for 48 hours at 42°C under microaerobic conditions. An inoculum of each strain was adjusted to 0.5 McFarland in 5 ml of MHB. About 10 μl of this suspension was stabbed into motility agar. Plates were incubated at 42°C under microaerobic conditions, and the diameter of the resulting swarms was measured after 24 hours. The results were the mean of five separate repetitions.
Cytotoxicity assay
The cytotoxicity assay was performed as described by Coote et al. 11 with minor modifications. Briefly, cultures of wild-type strains and mutants were prepared as described above. An inoculum of each strain was adjusted to 0.5 McFarland in DMEM containing 1% FBS, 1% NEAA, and 0.2% polymyxin B (Sigma). As demonstrated by Guerrant et al., 21 polymyxin B enables cytotoxin extraction in C. jejuni. After incubation of cultures at 37°C for 1 hour, cells were centrifuged for 20 minutes at 8,000 rpm, and the supernatants filtered using 0.22-μm filters (Fisher Scientific, Illkirch, France).
Caco-2 cells were seeded at 105 cells per well in 96-well plastic plates and incubated at 37°C in the conditions described above to allow cell adherence. After 24-hour incubation, the medium was discarded and replaced by 100 μl of a twofold dilution of the Campylobacter polymyxin B extract. Cells exposed to the DMEM containing 1% FBS, 1% NEAA, and 0.2% polymyxin B and cells treated with 0.1% Triton X-100 served as negative and positive controls, respectively. Monolayers of cells in the presence of bacterial extracts were incubated at 37°C as described above. After 24-hour incubation, bacterial extracts were discarded, and cells were washed with PBS 1×, and then 100 μl of 0.075% 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT; Sigma) in PBS was added to each well. In a living eukaryotic cell, MTT is cleaved by dehydrogenase activity to produce a dark-blue Formazan product. After 3-hour incubation, the medium was removed and replaced by 100 μl of 0.04 M of HCl in isopropanol to solubilize and quantify the formazan product. An A540 measurement was taken using a Multiskan plate reader (Labsystems, Cergy-Pontoise, France), and the percentage of cytotoxicity in each well was calculated according to the following formula:
Statistical analysis
For all assays, mean values were compared using either Student's t-test or the Mann and Whitney test, depending on the normality of distributions and homogeneity of variances. Differences between the mean values were considered significant at p<0.01 (for adhesion and invasion assays) or p<0.05 (for motility and cytotoxicity assays).
Results
Characteristics of Campylobacter wild-type strains and Campylobacter-resistant mutants
The characteristics of wild-type strains and their fluoroquinolone-resistant mutants are presented in Table 1. For wild-type strains Cc103 and Cj126, the MICs of ciprofloxacin were, respectively, 0.25 and 0.06 mg/L. Fluoroquinolone-resistant mutants had MICs of ciprofloxacin between 2 and 8 mg/L. No changes between the MICs of antibiotics of other families between wild-type strains and their isogenic mutants were observed. After sequencing the QRDRs of the gyrA and gyrB genes, no mutations either in gyrA or gyrB were detected in the wild-type strains compared to C. coli NCTC 11353 or C. jejuni NCTC 11168 strains. Resistant mutants harbored Asp90Asn, Asp90His, and/or Thr86Ile substitutions in the GyrA subunit, but no mutations were detected in the GyrB subunit.
MICs (mg/L) determined by the agar dilution method.
MIC, minimal inhibitory concentration; CIP, ciprofloxacin; NAL, nalidixic acid; ENR, enrofloxacin; ERY, erythromycin; AMP, ampicillin; TET, tetracycline; Asp, aspartic acid; Asn, asparagine; Thr, threonine; Ile, isoleucine; His, histidine; FQS, susceptible to fluoroquinolones; FQR, resistant to fluoroquinolones; —, no mutation was detected in the gyrase subunit.
The characteristics of the wild-type strains and their macrolide-resistant mutants are presented in Table 2. Wild-type strains had MICs of erythromycin between 2 and 4 mg/L. Erythromycin-resistant mutants had MICs of erythromycin between 64 and >128 mg/L. In addition, all mutants were resistant to tylosin, azithromycin, and lincomycin, whereas the parent strains were susceptible. No changes were observed between the MICs of antibiotics of other families for the wild-type strains and their isogenic mutants. After sequencing the three gene copies encoding 23S rRNA, rplD, and rplV, no mutations were detected in the wild-type strains. Resistant mutants harbored A2074C/G and/or A2075G in 2 or 3 copies of genes encoding 23S rRNA and/or G221A in rplD.
MICs (mg/L) determined by agar dilution method.
rrlA, rrlB and rrlC represent the three gene copies encoding 23S rRNA. rplD and rplV encode ribosomal proteins L4 and L22, respectively.
TYL, tylosin; AZM, azithromycin; LIN, lincomycin; Cc, Campylobacter coli; Cj, Campylobacter jejuni; MacS, susceptible to macrolides; MacR, resistant to macrolides; — , no mutation was detected in the gene.
Impact of antibiotic resistance on adhesion and invasion characteristics tested with Caco-2 cells
Impact of fluoroquinolone resistance
All the examined strains and mutants adhered to Caco-2 cells within a range of 0.01%–1% of the starting bacterial inoculum (Fig. 1). No significant difference was observed between the adhesion of wild-type strains and those of their isogenic fluoroquinolone-resistant mutants, either for C. coli or C. jejuni. The invasiveness of Campylobacter strains and mutants into Caco-2 cells was found to lie within a wide range of 0.0001%–0.01% of the starting bacterial inocula (Fig. 1). For C. coli, as shown in Fig. 1a, the fluoroquinolone-resistant mutant Cc103A harboring mutation Asp90Asn in its GyrA subunit had a significantly higher (p=0.009) Caco-2 cell invasion mean than the susceptible strain (Inv% of 0.00059±0.00034 for Cc103 vs. 0.02±0.011 for Cc103A). Similar results were obtained for mutant Cc103T harboring substitution Thr86Ile in its GyrA subunit. This mutant had a significantly higher (p=0.009) Caco-2 cell invasion mean than the susceptible strain (Inv% of 0.00059±0.00034 for Cc103 vs. 0.011±0.0095 for Cc103T). However, no significant difference was observed between the rate of invasion of the wild-type strain and that of the resistant mutant Cc103A+T harboring the two mutations Asp90Asn and Thr86Ile in its GyrA subunit. For C. jejuni, as shown in Fig. 1b, fluoroquinolone-resistant mutants Cj126A, Cj126T, and Cj126A+T had a slightly lower invasiveness rate into Caco-2 cells than did the susceptible parental strain Cj126. However, this decrease in invasiveness was not significant (p>0.2).

Adhesion and invasion of wild-type strains and fluoroquinolone-resistant mutants to intestinal epithelial cells (Caco-2).
Impact of macrolide resistance
All macrolide-susceptible strains and their resistant mutants adhered to Caco-2 cells within a range of 0.01%–1% of the starting bacterial inoculum (Fig. 2). No significant difference was observed between the adhesion of wild-type strains and that of the macrolide-resistant mutants, either for C. coli or C. jejuni. The invasiveness of Campylobacter strains and mutants into Caco-2 cells was found to lie within a wide range of 0.0001%–0.01% of the starting bacterial inocula (Fig. 1). In C. coli, as shown in Fig. 2a, the macrolide-resistant mutant Cc454m harboring the A2075G substitution in the three gene copies encoding 23S rRNA had a Caco-2 cell invasion mean similar to that of the susceptible strain Cc454. Similar results were obtained for the mutants C3830m and Cc4053m harboring mutations A2074C and A2074G in the three gene copies encoding 23S rRNA, respectively. In C. jejuni, as shown in Fig. 2b, resistant mutant Cj126m had a slightly lower Caco-2 cell invasion mean than the susceptible strain, but the reduction was not significant (p=0.2). However, Cj126m2 harboring G221A in rplD as well as A2074C and C2551G in one copy gene encoding 23S rRNA showed a significant drop (p=0.009) in the invasion mean of Caco-2 cells compared to the susceptible strain (Inv% of 0.0084±0.0072 for the wild-type strain Cj1216 vs. 0.0012±0.00031 for the resistant mutant Cj126m2). On the other hand, the resistant mutant Cj008m harboring A2074G substitution in two gene copies encoding 23S rRNA showed a significant rise (p=0.009) in the mean invasion of Caco-2 cells compared to its wild-type strain Cj008 (Inv% of 0.00029±0.00026 for the wild-type strain Cj008 vs. 0.0033±0.0028 for the resistant mutant Cj008m).
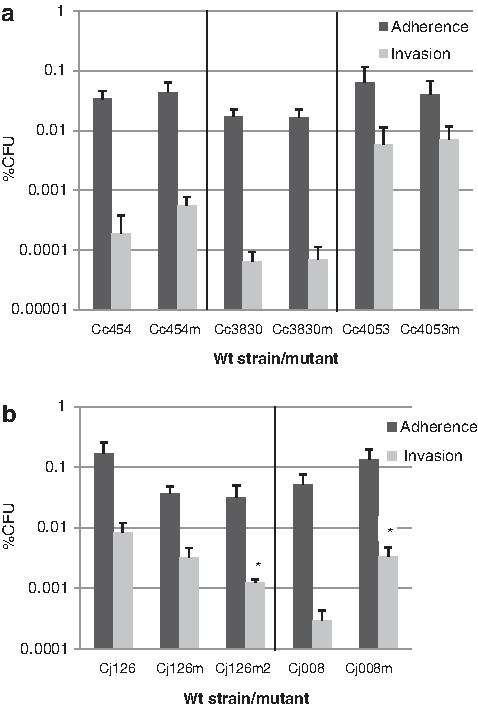
Adhesion and invasion of wild-type strains and macrolide-resistant mutants to intestinal epithelial cells (Caco-2).
Impact of antibiotic resistance on Campylobacter motility
The motility phenotypes of wild-type strains and resistant mutants were determined using Mueller-Hinton motility agar. As shown in Fig. 3, growth zone diameters for all susceptible strains and resistant mutants ranged between 2.2 and 5 cm. For C. coli, no significant difference was observed between the motility of each wild-type strain and mutant regardless of the resistance trait being to the fluoroquinolone or the macrolide. For C. jejuni, a significant difference was observed between the motility of wild-type strain Cj126 and that of the mutants resistant to fluoroquinolones or macrolides. Mutants Cj126A, Cj126T, and Cj126A+T harboring Asp90His, Thr86Ile, and both substitutions, respectively, in GyrA subunit showed reduced motility (p=0.009) compared to the susceptible wild-type strain (the growth zone diameter for Cj126 was 3.78±0.21 cm, whereas growth zone diameters for resistant mutants ranged between 2.41±0.49 cm and 2.92±0.21 cm). Similar results were obtained for macrolide-resistant mutants. Mutants Cj126m and Cj126m2 harboring G221A in rplD and A2074C and/or C2551G in one gene copy or 23S rRNA also showed reduced motility (p=0.009) compared to the susceptible wild-type strain (the growth zone diameter for Cj126 was 3.78±0.21 cm, whereas growth zone diameters for resistant mutants ranged between 2.61±0.51 cm and 2.67±0.39 cm). However, no significant difference was observed between the motility of strain Cj008 and that of its macrolide-resistant mutant Cj008m harboring A2074G in two gene copies encoding 23S rRNA.
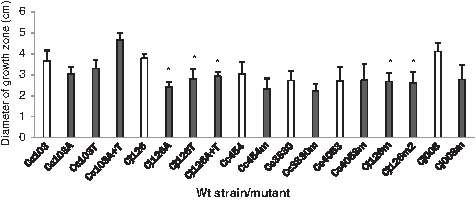
Growth zones of wild-type strains and resistant mutants on Muller-Hinton motility agar at 42°C. White bars: Growth diameter of wild-type susceptible strains. Black bars: Growth diameter of resistant mutants. The asterisk (*) represents a statistically significant difference between a mutant and its parental strain.
Impact of antibiotic resistance on the cytotoxic activity of Campylobacter
Polymyxin B extracts of all susceptible strains and resistant mutants were tested for cytotoxic activity against Caco-2 cells. Bacterial extracts displayed cytotoxicity at a range of 93%–100% over the 24-hour period, indicating high toxin activity for all strains. However, except for mutant Cc103A, the titers determined by twofold serial dilutions of the cell-free supernates showed that there was no difference in cytotoxic activity between susceptible strains and their respective resistant mutants. Nevertheless, toxin production of the mutant Cc103A was at least two times higher than that of the wild-type strain Ccc103 (p<0.05), indicating that mutation Asp90Asn in the GyrA subunit could enhance cytotoxic activity in C. coli.
Discussion
Previous studies13,25 have shown that infections with antibiotic-resistant Campylobacter lead to more severe diseases in humans, although other reports17,35 did not confirm this relationship. However, all of these studies were based on clinical and epidemiological data that cannot clearly attribute the impact of antibiotic resistance on Campylobacter virulence, thereby neglecting genetic diversity between susceptible and resistant strains. The study of the virulence characteristics of a large panel of susceptible and resistant field strains could be conducted, but in an attempt to investigate whether resistance could modify some of the characteristics of Campylobacter that may influence its virulence, we took advantage of a collection of susceptible and isogenic resistant mutants to confidently evaluate the impact of resistance on some potential mechanisms of virulence. Mutants had been obtained either by selection on supplemented media or by transformation with small isogenic DNA fragments (except the specified mutation), resulting in globally similar genetic environments between susceptible strains and their mutants. Although the number of strains was rather limited, the two most frequent fluoroquinolone mutations and their association were studied for each Campylobacter species, giving thus six different mutants, and for macrolides, the various mutations that have been previously reported in field strains or laboratory mutants were included in our study (three copies A2075G, three copies A2074C, two copies of A2074G, three copies A2074G, association of A2074C, and mutations in rrlD and/or rrnB).
The mechanisms of Campylobacter pathogenesis include various important stages: adhesion to intestinal cells, invasion of targeted cells, and toxin production. 22 Moreover, it has been demonstrated that motility is essential for Campylobacter penetration of host epithelial cells. 20 These characteristics were therefore chosen to be investigated in our study.
Coote et al. 11 demonstrated that C. jejuni isolates adhere more effectively to human colonic Caco-2 cells than other cell types, and have a greater capacity for Caco-2 cell invasion than their invasive capacity with respect to HeLa and Vero cells. Caco-2 cells were thus used in our experiments to investigate Campylobacter virulence.
It has been demonstrated that Campylobacter virulence involves flagellum expression, as well as a set of proteins implicated in adhesion to and invasion of host epithelial cells.26,27,33 Thus, the process of bacterial infections requires efficient protein synthesis to enhance motility, invasion, and cytotoxin production. GyrA is known to be a key enzyme for DNA supercoiling and an essential protein for DNA replication and transcription. 9 Mutations in the gyrA gene could enhance or reduce the expression of multiple genes, which may consequently affect the expression of a subset of proteins involved in bacterial virulence. Surprisingly, the C. coli mutant Cc103 (but not the C. jejuni Cj126) harboring the Asp90Asn +Thr86Ile mutations showed invasion scores similar to the wild-type strain, maybe because of a modified structure of the C. coli gyrase, each mutation abolishing the effect of the other one. Similarly, multiple mechanisms ensure correct rRNA activity, which is essential for an active transcription machinery. 30 In our conditions, it seems that Asp90Asn or Thr86Ile mutations enhance the transcription activity of the GyrA subunit in C. coli, allowing a more efficient invasion or cytotoxin production in fluoroquinolone-resistant mutants. However, in C. jejuni, whenever the motility was affected, fluoroquinolone- or macrolide-resistant mutants could not show an enhanced virulence activity. These observations confirm that adhesion and invasion properties are strongly related to motility characteristics.27,33 Recently, Almofti et al. 3 showed that an erythromycin-resistant C. jejuni mutant harboring A2074C in three gene copies of rRNA adheres to and invades intestinal epithelial cells less than the susceptible strain. In another study, 4 they reported that A2074C substitution can impair the motility of C. jejuni in 0.4% agar plates, but this same mutation may have no effect in another recipient strain in the absence of macrolide selection pressure. These diverse effects of macrolide resistance could be largely attributed to the genetic background of bacterial strains as well as to the different mutations in genes implicated in macrolide resistance.
Toxin production, tested using Caco-2 cells, appeared similar in susceptible strains and resistant mutants, with the exception of Cc103A. However, Florin and Antillon 18 suggest that two different cell types should be used to detect cytotoxin production in Campylobacter. It may thus be of interest to determine whether antibiotic resistance affects cytotoxin production using other cell types.
Finally, it should be mentioned that other nondetected mutations may have arisen within the genome during the selection of resistant mutants. Such additional mutations could play a role in the difference observed between mutants and susceptible strains, but complete sequencing of the genome of all isolates and their mutants was not feasible.
To summarize, our results showed that mutations that are frequently encountered in resistant field and clinical isolates could enhance the invasiveness of Campylobacter in Caco-2 cells. Inversely, mutations conferring reduced bacterial motility were never associated with a significantly higher Caco-2 cell invasion capacity than parental strains. Studies on a larger panel of isolates with mutations at each location and combinations thereof are in progress to confirm these preliminary results concerning consequences of resistance mutations on invasiveness and bacterial mobility.
Footnotes
Acknowledgments
We are grateful to Anne Bouchardon (Anses, Ploufragan) for her remarks during the experiments. We would also like to thank Thomas Cousseau for experiments carried out during his undergraduate internship. This article was partly presented during the “International Conference on Zoonoses” in London, September 2011. This work was supported by ANSES (French Agency for Food, Environmental, and Occupational Health Safety), and Conseil Général des Côtes d'Armor. Salman Zeitouni was financed by the French Ministry of Research.
Disclosure Statement
No competing financial interests exist.