
Abstract
Tuberculosis causes close to 1.5 million deaths in the world, with new cases exceeding 9 million in recent years. Coinfection with HIV further worsens the global situation. New molecules that overcome the limitations of currently used drugs are needed. We aimed to determine whether HHC-10 is active against the Mycobacterium tuberculosis complex bacteria Mycobacterium bovis bacille calmette guerin (BCG) in vitro and in vivo. For this, HHC-10 was tested in vitro using different peptide concentrations, and in vivo, in C57BL/6 mice infected intratracheally, at two doses (1.25 and 2.5 mg kg−1, once a week, 4 weeks). Interferon (IFN)-γ, TNF-α, interleukin (IL)-4, and IL-10 mRNA transcript levels were compared between treated and nontreated mice. In vitro, HHC-10 decreased 69% and 88% the number of colony-forming units (CFU) per millileter recovered after 24-hr treatment at 50 and 100 μg/ml, respectively. In vivo, BCG CFUs in mouse lungs were reduced 77.8% and 95.8% at 1.25 and 2.5 mg kg−1, respectively. IFN-γ expression was lower in the HHC-10-treated group than that of nontreated animals. Considering genomic conservation between BCG and M. tuberculosis, the in vitro and in vivo activities of HHC-10 observed in this study suggest that the use of this peptide may be useful as therapeutic agent against tuberculosis.
Introduction
When TB is caused by infection with an antibiotic-susceptible strain, its treatment requires a combined therapy of multiple antibiotics (rifampicin, isoniazid, pyrazinamide, and ethambutol) for 6–9 months; this relatively long period of time contributes with problems for successful treatment. A further concern is the development of drug-resistant (including multi-(MDR), extensively (XDR), and totally drug resistant (TDR)) strains of Mycobacterium tuberculosis. It is very important to identify and develop new molecules that may be used as complement, especially against those strains that are drug resistant. Antimicrobial peptides (AMPs) are widely distributed in several organisms such as insects, plants, and mammals. In mammals, there are two main families: defensins and cathelicidins. 5 Intensive research has led to the development of synthetic peptides with enhanced antimicrobial activities, some of them, including capacity to stimulating host immune cells, to facilitate clearance of an infection in vivo. 4 The immunomodulatory activities of such peptides are multiple and include stimulation of chemotaxis and chemokine production, stimulation of epithelial cell migration, and many others. 15 Kandler found that LPS in combination with LL-37 led to a decrease in expression of HLA-DR, CD86, and other markers when applied to DCs. 14 These findings are consistent with the concept that AMPs modulate rather than strongly suppress or stimulate immune responses.
To our knowledge, the capacity of HHC-10 has not been explored to date, regarding its effect against microorganisms other than Staphylococcus aureus in vivo. In this work, we aimed to determine for the first time whether this cathelicidin inhibits the M. tuberculosis complex bacteria Mycobacterium bovis bacille calmette guerin (BCG) in vitro and in vivo. Our results suggest that HHC-10 could be used as future therapy to combat TB.
Materials and Methods
Determination of minimum inhibitory concentration of HHC-10 in vitro
M. bovis BCG Pasteur 1173P2 was grown in the Middlebrook 7H9 broth supplemented with 10% oleic acid-albumin-dextrose-catalase (OADC; Difco Laboratories), 0.05% Tween 80, and 0.2% glycerol (Difco) in an atmosphere of 5% CO2–95% air. The synthetic cathelicidin HHC-10 has been previously described,4,6 and it was synthesized and purchased at EZ Biolab at 95% purity with no amidation. M. bovis BCG was harvested at the mid-log phase by pelleting at 14,000 g for 10 min at 25°C and suspended in warm 7H9 broth. From a culture of M. bovis BCG reaching 0.5–0.6 O.D.600nm, we adjusted it to 0.02 O.D.600nm (106 colony-forming units [CFU]/μl). Then, 100 μl of HHC-10 were added to reach a final volume of 1 ml, to obtain concentrations of 25, 50, and 100 μg/ml. For the control, 100 μl sterile water were added. We next plated serial dilutions at time 0 and 24 hr, onto Middlebrook 7H10 agar enriched with 10% OADC and 0.5% glycerol, and incubated at 37°C for 21 days. CFUs were counted. All tests were done in quadruplicate for each concentration and control, and the experiments were repeated on different dates three times.
Mice
Pathogen-free, female, 8–9 weeks old C57BL/6 mice were purchased from Harlan Laboratories (Mexico). Mice were housed in microisolator cages (HEPA class 100) and fed a standard rodent diet and water ad libitum. Experiments were performed in accordance with SAGARPA's (Ministry of Agriculture, Livestock, Rural Development, Fisheries and Food) animal care ethics approval and guidelines.
Infection and CFU enumeration
For each experiment, M. bovis BCG cultures at 0.8 O.D.600nm were harvested (14,000 g for 15 min), suspended in phosphate-buffered saline (PBS), and diluted to 107 CFU/ml. Mice were anesthetized intraperitoneally with zoletil® 50 solution (0.15 ml/kg body weight). Incising the skin and fascia in the midline exposed the trachea. A total of 105 CFUs in 50 μl PBS were injected to all mice using a 0.5-ml microsyringe (Becton Dickinson). The skin was closed with type-5-0 absorbable chromic catgut surgical thread (Atramat). AMP HHC-10 was diluted in sterile water and administered subcutaneously in the upper back, over the shoulders, of experimental groups of mice. Treatment was started 3 weeks postinfection (when the highest bacillary load was reported for C57BL/6 mice intratracheally infected with BCG 10 ) at doses of 1.25 and 2.5 mg kg−1, once a week during 1 month. Control mice received sterile water only. Three mice were sacrificed at the indicated time points. Three lungs and spleens were collected and divided from all mice at 24 hr, 1, and 3 weeks postinfection, and 4 weeks post-treatment with HHC-10. The half part of the lung and spleen of each mouse were triturated by Polytron (tissue tearer pattern # 985370-395) and centrifuged (5000 rpm for 1 min) to determine mycobacterial CFUs after plating onto 7H10 agar plates. The remaining half of the organ was cryopreserved at −80°C for RNA isolation. For histopathology, the lungs were perfused with formaldehyde 10%, sectioned, and stained using hematoxylin–eosin. Three different pathologists evaluated these sections separately, in a blind manner. Infection experiments were repeated two times in different months with similar results, and figures in the Results section are those from one representative experiment.
Quantitative real-time polymerase chain reaction
Three lungs of experimental groups, from two different experiments (six samples in total), were used for isolating RNA and synthesis of cDNA. Real-time polymerase chain reaction (PCR) was performed using the StepOne real-time PCR system (Applied Biosystems). Tissue was homogenized in 1 ml of TRIzol® reagent (Invitrogen) and extracted according to the manufacturer's instructions, treated with RNase-Free DNase (Promega), and cDNA prepared with the AffinityScript QPCR kit (Agilent Technologies). We used 108 ng cDNA for quantitative real-time PCR (qPCR). Primer/probe was selected from the Applied Biosystems TaqMan® Gene Expression Assays product line. Transcript levels of the proinflammatory cytokines such as interferon (IFN)-γ (RefSeq NM_008337.3) and tumor necrosis factor-alpha (RefSeq MN_013693.2), anti-inflammatory cytokines such as interleukin (IL)-4 (RefSeq NM_021283.2) and IL-10 (RefSeq NM_010548.2), and endogenous control rRNA 18S (RefSeq NR_003278.1) were assessed by qPCR with the TaqMan Universal PCR Master Mix (Applied Biosystems). The cell-cycling conditions used were initial denaturation at 95°C for 10 min, followed by 40 cycles each at 50°C for 2 min, 95°C for 10 min, and 60°C for 1 min. Quantities of the specific mRNA in the sample were measured in accordance with the corresponding gene-specific standard. The data were analyzed using the comparative Cq method and normalized against rRNA18S expression. Expression of each cytokine and the internal reference 18S was determined for each experimental group and compared to the nontreated group.
Statistical analysis
Data from the CFUs and qPCR were analyzed as follows. Kruskal–Wallis tests were performed for each data set to choose the appropriate group comparison test. We compared treated versus non-treated groups for CFUs recovered in lungs and spleens, as well as for the relative expression of each cytokine. After one-way analysis of variance, results that showed significant differences were submitted to pairwise comparisons by Tukey's or Dunn's post-test. Two-sided p-values of <0.05 were considered statistically significant. Statistical analyses were performed using the GraphPad Prism version 5.03 for Windows (GraphPad Software).
Results
Antimicrobial activity of HHC-10 in vitro against mycobacteria
We first assessed HHC-10 activity in vitro against M. bovis BCG by determining the reduction of viable bacilli by CFU assay using different peptide concentrations (0, 25, 50, and 100 μg/ml), and CFUs/ml were determined at time zero and 24 hr later, as described in the Materials and Methods section. At time zero, HHC-10 did not affect CFU/ml at any concentration; however, the peptide significantly reduced the number of viable bacilli (Fig. 1) after 24-hr treatment (p<0.05). A reduction in the CFU/ml by 69% and 88% at 50 and 100 μg/ml, respectively, compared with the control was observed.
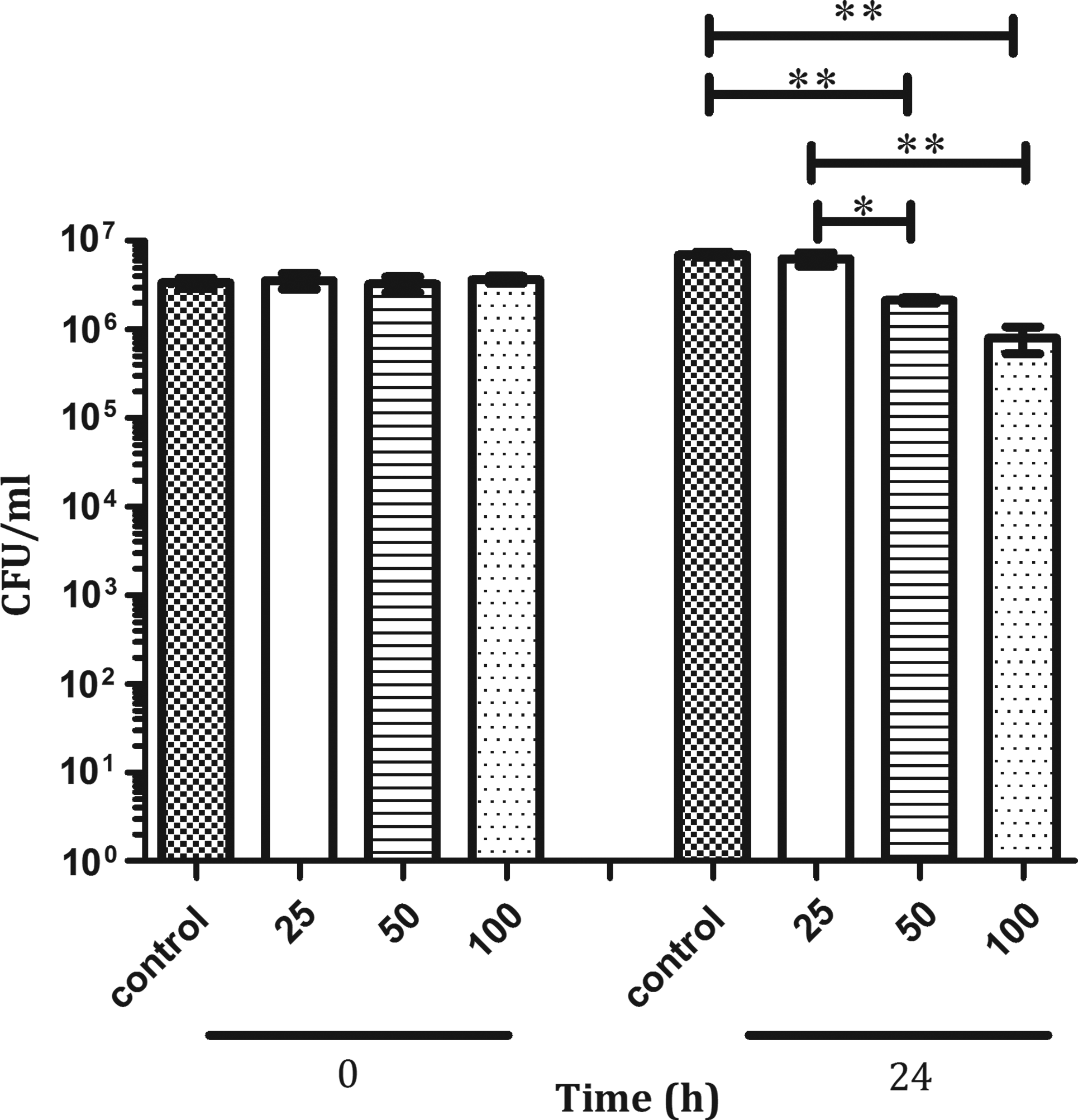
Mycobacterium bovis BCG survival after incubation with HHC-10. Experiments were performed using various concentrations of HHC-10. The bacterial survival was measured by colony-forming unit (CFU) assay. Experiments were performed as described in the Materials and Methods section. Results shown are for one representative experiment. Asterisks indicate statistical significance *p<0.01; **p<0.001. X-axis shows HHC-10 concentration in μg/ml.
HHC-10 treatment of C57BL/6 mice reduced bacterial load of M. bovis BCG
To determine the capacity of HHC-10 to diminish mycobacterial load in vivo, we used intratracheal infection of C57BL/6 mice with M. bovis BCG as recently reported, 10 basically because we do not have the capacity to infect mice with M. tuberculosis nor via aerosol in our laboratory. Infection was allowed to progress for 3 weeks. Afterward, mice were treated four times, once a week, with HHC-10 at 1.25 and 2.5 mg kg−1 doses for 4 weeks; control groups received only sterile water. Before starting peptide treatment, three mice from each group were sacrificed at 24 hr, 1, and 3 weeks postinfection (Fig. 2A) to determine CFUs. We found that our treatment diminished viable bacilli in the lungs (Fig. 2A). These reductions were 77.8% and 95.8% at 1.25 and 2.5 mg kg−1, respectively. These reductions were statistically significant (p<0.01).

CFU enumeration in the lung and spleens of infected mice treated with HHC-10. ), HHC-10 at 1.25 mg kg−1 ( ) and 2.5 mg kg−1 (
) and 2.5 mg kg−1 ( ) per mouse was given subcutaneously for 1, 2, 3, and 4 weeks. The experiments were repeated on different dates two times, only one representative experiment is shown, and the second experiment yielded similar results. Asterisk represents statistical significance *p<0.05 and **p<0.01.
) per mouse was given subcutaneously for 1, 2, 3, and 4 weeks. The experiments were repeated on different dates two times, only one representative experiment is shown, and the second experiment yielded similar results. Asterisk represents statistical significance *p<0.05 and **p<0.01.
Mycobacterial migration and establishment in spleen were observed at 1 week postinfection, and a significant increment (p<0.01) at 3 weeks postinfection was found (Fig. 2B). After 4 weeks treatment, a significant increase (p<0.05) in mycobacterial load was observed in the spleen of group of mice that received 1.25 mg kg−1 with respect to the control group. No significant change was observed in the spleen in the group of mice that received 2.5 mg kg−1 with respect to the control group.
We performed histopathological analysis of lung sections (Fig. 3) of our different groups and found minimal inflammation in BCG-infected mice (control, non-HHC-10 treated), medium-size granulomas with mild perivascular inflammation (1.25 mg kg−1 HHC-10), and medium-size granulomas with moderate inflammation (2.5 mg kg−1 HHC-10).

Micrographs of lungs from C57BL/6 mice infected with BCG. Mice were treated with HHC-10 at the indicated doses, or vehicle (control). Magnification is 50×.
Cytokine expression in lungs after HHC-10 administration to M. bovis BCG-infected mice
To investigate the capacity of HHC-10 to modulate gene expression of cytokines well known to affect TB outcome, that is, IFN-γ, TNF-α, IL-10, and IL-4, we determined their levels by reverse transcription and quantitative real-time PCR. After 4 weeks of treatment, inhibition of all cytokines expression, but IL-4 (not detected), was observed at both doses relative to control mice (Fig. 4). However, only IFN-γ showed a statistically significant reduction (50% and 70% at 1.25 and 2.5 mg kg−1, respectively) in comparison with the nontreated infected mice (Fig. 4).
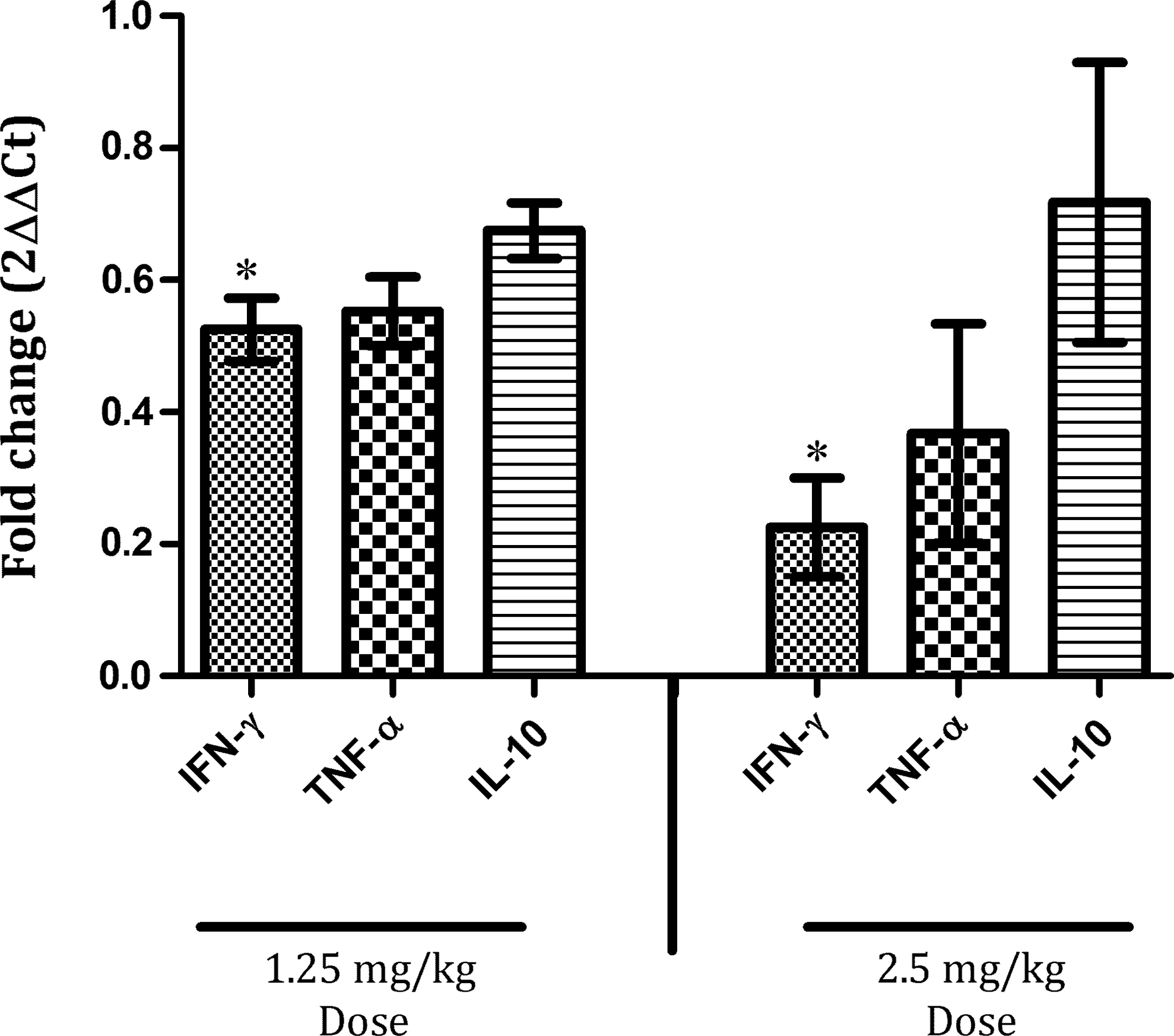
Peptide HHC-10-modified cytokine expression in the lungs of BCG-infected C57BL/6 mice. Mice were treated subcutaneously with peptide HHC-10 and control (1.25 and 2.5 mg kg−1) for 1 month and analyzed for cytokine gene expression by quantitative real-time polymerase chain reaction. The data were analyzed using the comparative Cq method and normalized against 18S rRNA. Asterisk represents statistical significance when compared with the control nontreated and group treated (*p<0.0001).
Discussion
Synthetic peptides showing improved bactericidal effects and/or capacity to stimulate protective immune responses could provide an alternative for shortening long-lasting drug regimens and help to reduce the global MDR and XDR-TB burden. 7 The synthetic cathelicidin HHC-10 has been reported previously to have a bactericidal effect in vitro against multidrug-resistant pathogens (also called superbugs) such as Pseudomonas aeruginosa, Escherichia coli, Klebsiella pneumonia, and S. aureus, among others, 6 where it exhibited minimum inhibitory concentrations (MICs) of 0.3–11 μM against most drug-resistant microorganisms. As we found significant decrease in bacterial replication in vitro for 50 and 100 μg/ml (34–68 μM), it is clear that drug-susceptible BCG is more resistant in vitro to HHC-10 than drug-resistant Pseudomonas, E. coli and S. aureus.
In vitro results showed a significant reduction of viable BCG in a dose-dependent manner reaching 69% reduction at 50 μg/ml and 88% at 100 μg/ml with respect to the control (Fig. 1). In our research group, we found that the synthetic cathelicidins HHC-10 and Bac2A inhibited M. tuberculosis H37Rv with a MIC of 100 μg/ml. Interestingly, the same groups of AMPs were capable of inhibiting the clinically isolated multidrug-resistant CIBIN99 strain with half of concentration MIC of 50 μg/ml (Cappoen et al., manuscript submitted). Other AMPs have shown bactericidal effects against mycobacteria such as M. tuberculosis. The defensin-class HNP-1 and protegrin-1 were shown to be bactericidal in vitro (ranging from 86% to 99% killing) against M. tuberculosis (H37Ra) at 15 μM (50 μg/ml) and 23μM (100 μg/ml), respectively. 18 Interestingly, the killing capacity was equal to that observed when clinical isolates were tested (reviewed by Flores-Valdez and Chopra 7 ). We can see that HHC-10 activity against BCG is similar to that of HNP-1 and protegrin-1 against M. tuberculosis. On the other hand, Martineau and cowokers tested different concentrations of LL-37 (0.1–100 μg/ml) and found that this cathelicidin inhibited the grow of M. tuberculosis in vitro with an MIC of 100 μg/ml. 17 We therefore tested 100 μg/ml and lower concentrations of HHC-10 based on the reported activity for another cathelicidin.
Lungs are the main organs attacked by M. tuberculosis. As shown in Fig. 2, our infection model corresponds to that previously reported by Fulton and coworkers. 10 A regimen of four administrations of HHC-10, once a week, reduced by 77.8% (1.25 mg kg−1 dose) and 95.8% (2.5 mg kg−1 dose), the number of viable bacilli in mice lungs (Fig. 2A). Although in lungs, a significant reduction of viable bacilli was seen; in the spleen, we found an increase at 1.25 mg kg−1 dose of peptide in comparison with nontreated mice. These results were unexpected, and considering the route of administration, it may simply be that the peptide was not able to reach the spleen, although this remains to be experimentally determined. It may be possible that the function of the peptide is restricted to the lungs due to a localized effect. According to Fulton's mouse model, there is dissemination of M. bovis BCG from lungs to lymph organs such as the spleen, 10 similar to what we observed (Fig. 2B).
Despite the fact of HHC-10 reducing on average 99.6% the bacterial load when administered via intraperitoneal at 4 mg kg−1 in a mouse model infected with S. aureus, we decided to test doses of 1.25 and 2.5 mg kg−1 basically because we were infecting mice with an organism (BCG) that during the course of infection is gradually, and completely (spleen) or not (lungs) eliminated. 10 Reasons for the observed halted replication include direct mycobacterial killing (Fig. 1) and perhaps HHC-10 promoted granuloma formation (Fig. 3), although experiments where markers for granuloma formation are evaluated should be performed. Analysis of local cell populations might help in explaining this finding.
It has been suggested that cathelicidin LL-37 uptake augments the bactericidal activity of macrophages infected with M. tuberculosis. 21 It could well be that HHC-10 possess some of the activities described for LL-37. Although we did not determine how long does HHC-10 remain present in vivo, it has been reported that LL-37 has an estimated 3.4-day half-life in C57BL/6 mice. 1 Given we were able to observe effects against BCG even after administering HHC-10 only once a week, we think that it might remain in vivo for a time equal to or greater than LL-37. New experiments where treatment regimen varies (change in peptide dose, times the peptide is administered, and testing against virulent mycobacteria) as well as determining, for example, serum and spleen cytokine levels, or immune cell recruitment during M. tuberculosis infection, will shed more light on the feasibility of using HHC-10 as therapy against TB.
The ability of AMPs to mediate the attraction of various immune cell types, modulation of anti-inflammatory cytokines, and promoting the adaptive immune response have been demonstrated extensively. 13 All these kind of immune mechanisms are regulated by chemokines and cytokines. The presence of IFN-γ is crucial for immunity against pathogens such as M. tuberculosis. It is well known that mice harboring mutation in the IFN-γ receptor enhances its susceptibility to mycobacterial infection. 8 Moreover, IFN-γ influences dendritic cell maturation by upregulating costimulatory molecules, antigen processing, and presentation. However, an important event related with the immunity toward mycobacterial infection is the enhancing of antimicrobial properties in macrophages, the natural cell host for M. tuberculosis. Our finding of a significant reduction of IFN-γ transcription upon HHC-10 administration to infected mice (Fig. 4) could be the consequence of either reduced bacillary load not further triggering this cytokine expression, or the direct consequence of the peptide reducing immune functions. It is important to note that similar effects on the immunomodulation of proinflammatory cytokines have been observed in mice infected and treated with LL-37.2,6 Further studies of more cytokines are needed to determine why despite IFN-γ decreased expression, there was containment of BCG replication in lungs.
Macrophages are the main cells that produce TNF-α, which is important as has been reported previously in the mycobacterial control during acute infection. 9 Mice lacking TNF-α or the TNF-α receptor succumb rapidly after infection. 3 Furthermore, it has been suggested that that these IFN-γ and TNF-α act synergistically to induce nitrogen intermediates that eventually reduce bacterial burden. 8 On the other hand, IL-10 acts as an antagonist of IFN-γ and TNF-α and is considered a deactivation molecule in infected macrophages. This cytokine is produced mainly by macrophages and CD4+T cells during mycobacterial infection. IL-10 may play a role in the regulation and prevention of excessive inflammatory related with Th1 cytokines. The presence of IL-10 inhibits APC as well APC functions during infection. 12 However, mice lacking IL-10 were not more resistant to acute mycobacterial infection compared to wild type. 19 It seems that other mechanisms are related with susceptibility to infection and required further exploration.
To our knowledge, this is the first report indicating the therapeutic potential of HHC-10 against experimental mycobacterial infection in mice. However, further studies are required to determine how effective is HHC-10 or other cathelicidins against M. tuberculosis, including definition of the mode of action. As already noted, mycobacteria often lack genes homologous to those conferring AMP resistance in other bacteria. 7 However, it has been reported that inactivating the lysX gene in M. tuberculosis renders the bacteria unable to produce lysinylated phosphatidylglycerol in the cell envelope, and sensitive to cationic peptides as occurs in S. aureus and L. monocytogenes. 16 Furthermore, the lysX mutant showed defective growth in the mouse and guinea pig lungs, as well as reduced pathology relative to wild-type bacteria. 16 We would expect the M. tuberculosis lysX mutant to be sensitive to HHC-10, thus suggesting that it might interact with the cell envelope, a hypothesis that deserves to be explored.
Finally, should HHC-10 comply with more extensive pharmacokinetics, pharmacodinamics, preclinical, and clinical studies, and if proven to be safe and effective, it could contribute to shorten the treatment time of current therapies, or be used in drug-resistant cases.
Footnotes
Acknowledgments
The authors gratefully acknowledge Dr. Erika Marino and Q.F.B. Michel de Jesús Aceves-Sánchez for technical assistance in this work; Prof. Rogelio Hernández-Pando, Dr. Hugo Esquivel, and Dr. Vladimir Espinosa for independent assessment of histopathological samples; and Prof. Robert Hancock for kindly donating some synthetic cathelicidins for preliminary experiments. CONACYT grant 81018 to M.A.F.V. allowed development of this project. Y.Y.Ll.G. Received a CONACYT M.Sc. fellowship (number 247076.). C.P.R. is recipient of a postdoctoral fellowship from CONACYT (290616-CIATEJ. A.C.)
Disclosure Statement
No competing financial interests exist