
Abstract
Conditions facilitating resistance to quaternary ammonium compounds (QACs) were investigated in Staphylococcus aureus SK982 exposed to benzalkonium chloride (BAC; a member of QACs) under various circumstances. S. aureus SK982 carrying the qacA gene encoding for resistance to QACs was grown in the presence of stable or gradually increasing concentrations of BAC, or it was exposed to this antiseptic in the exponential phase of growth. Bacteria cultivated in the highest BAC concentrations that did not retard their growth comparing to the untreated control were subjected to real-time quantitative polymerase chain reaction analysis for relative expression of the efflux genes qacA and norA. Under such conditions, S. aureus SK982 tolerated a relatively low stable concentration of BAC (1.22 mg/L) when compared with a gradually increasing antiseptic concentration (tolerance of 4.88 mg/L). However, in both cases, qacA expression was not significant. The culture exposed in the exponential phase of growth tolerated the highest concentration of BAC (9.76 mg/L) as also accompanied by significant overexpression of qacA. Expression of norA was relatively low regardless of the conditions tested. It seems that under the short-term conditions, the phase of bacterial growth is more important for the expression of BAC resistance than the capability to adapt to this antiseptic. This study provides a deeper insight into the relevance of the qac genes in conferring resistance to QACs.
Introduction
So far, resistance to disinfectants at a genetic level has been described in quaternary ammonium compounds (QACs). QACs are cationic surface-active agents widely used in both food and medical environments as well as in various medicaments (e.g., eye drops and skin cleansing). Main advantages of QACs are their high stability, low corrosivity, and relatively low toxicity.24,26 The qac genes belong to the first described and epidemiologically most important genetic determinants of resistance to QACs. 12 The involvement of an efflux-based system in qac-mediated resistance was first observed by Jones and Midgley 17 and Tennent et al. 36 In accordance with that, an energy-dependent efflux pump relying on the proton motive force was later confirmed as a mechanism of resistance conferred by the qac genes.23,30 Further analysis of the QAC proteins revealed that these belong to a small multidrug resistance protein family (except for those encoded by qacA and qacB, which belong to the major facilitator superfamily) integrated in the cytoplasmic membrane via transmembrane segments and containing distinct subsets of amino acid residues involved in substrate recognition and binding.6,27,30
Although there is no doubt that the qac genes encode for the resistance to QACs, there are still a lot of questions requiring explanation. In our recent work, 16 we did not observe increased resistance to benzalkonium chloride (BAC, a member of QACs) in Staphylococcus epidermidis isolates carrying the qac genes. In Staphylococcus aureus, Smith et al. 34 demonstrated that differences in bactericidal concentrations of QACs between qac-positive and qac-negative isolates were statistically significant but lower than twofold. On the other hand, the authors observed that the qac genes were associated with at least four times higher resistance to another cationic biocide–chlorhexidine (CHX). It is known now that the qac genes confer resistance to more than 30 distinct cationic compounds belonging to at least 12 different chemical classes. 11 However, there are still a lot of contradictory reports concerning the resistance conferred by the qac genes as recently discussed by Jaglic and Cervinkova. 15 Therefore, the factors influencing expression of the various QAC pumps remain to be further explained.
In addition, the nor genes, encoding for resistance to fluoroquinolones, were also found to encode for resistance to antiseptics and, even more, to have similar substrates to those of the qac genes.9,37 This shows that resistance to antibiotics and disinfectants may closely interface. 22 Similarly to the qac genes, the nor genes are now recognized as those encoding for multidrug efflux pumps.25,37
The present work deals with a relevant question, namely, why the presence of QAC resistance genes in some cases does not result in resistance to QACs. In accordance with that, the goal of this study was to evaluate the bacterial response to the pressure of selected cationic antiseptics and to identify conditions that may facilitate resistance to QACs. For that purpose, tolerance to BAC, as well as transcription of the qacA and norA genes, was monitored in S. aureus cells under various circumstances.
Materials and Methods
Bacterial strain and culture conditions
S. aureus reference strain SK982 (DSM 9385) harboring the plasmid pSK1 carrying the qacA gene encoding for resistance to QACs was used in this study. Before experimental use, the strain was kept at −80°C in Nutrient Broth No. 2 (Oxoid, Basingstoke, UK) supplemented with 20% glycerol. Bacterial colonies taken from the second passage grown at 37°C for 16 hr on blood agar (Trios, Prague, Czech Republic) were resuspended in phosphate-buffered saline (pH 7.2) to a turbidity of one McFarland standard and further diluted in Brain–Heart Infusion (BHI; BioRad, Marnes-la-Coquette, France) to a final concentration of ∼1.5×105 colony forming units (CFU) per ml. Bacteria were then incubated at 37°C in BHI alone or supplemented with antiseptics as described below. After 8 hr of incubation, the cultures (including untreated controls) in the exponential phase of growth [i.e., the cultures with optical density (OD605) of 0.4–0.8, which corresponded to 1.2–2.3×108 CFU/ml; Fig. 1] were processed as described below. Then, the cultures were further incubated for 24 hr, at which time OD605 reached 1.4 to 1.7, which corresponded to 0.9–1.2×109 CFU/ml as quantified by agar plate counting. In other words, the highest concentrations of antiseptics that had no influence on bacterial growth after 8 and 24 hr of incubation (when compared with the untreated controls) were considered in this study.

An example of calibration line prepared for Staphylococcus aureus SK982 grown in Brain–Heart Infusion (BHI) at 37°C. Six measurements of optical density (OD605) performed in 30-min intervals during a time frame of 7.5 to 10 hr of incubation are shown. Calibration line was drawn taking into account five measurements (black dots) as shown in the figure.
Chemicals and experimental design
Two chemically distinct surface-active agents (BAC, a member of QACs, and CHX, a member of biguanides; both from Sigma Aldrich, Brondby, Denmark) were tested at concentrations indicated below. The stock solutions of antiseptics were prepared in distilled water (in a twofold dilution range) and immediately used. Amounts of the stock solution added to the culture never exceeded one hundredth of the BHI volume. The BHI culture to which distilled water was added instead of the stock solution served as an untreated control. All testing conditions (see below) were performed in three independent experiments.
Growth in the presence of BAC and CHX
In three independent experiments, bacteria were grown at 37°C on a shaker in 10 ml of BHI containing BAC or CHX. In further three experiments, bacteria were grown in the presence of both BAC and CHX, whereas bacteria grown in the presence of BAC alone served as a positive control. Regardless of whether tested alone or in combination, the concentrations of BAC and CHX were 1.22 and 0.31 mg/L, respectively. These concentrations were the highest concentrations (in log2 scale) that did not inhibit bacterial growth during the first 8 hr (as well as 24 hr) of incubation when compared with the untreated control. After 8 hr of incubation, 500 μl of the culture (including untreated control) was harvested for RNA isolation.
Growth in the presence of gradually increasing concentrations of BAC
Bacteria were grown at 37°C under agitation in 1 L of BHI in a Lambda Minifor bioreactor (Lambda Laboratory Instruments, Zurich, Switzerland). During the first 8 hr of incubation, a total of 3.04 ml of the stock solution of BAC (1.61 g/L) was continuously applied to the culture (0.38 ml/hr) using a peristaltic pump (Lambda Laboratory Instruments), so that the concentration of BAC in the BHI culture gradually increased from the initial zero to final 4.88 mg/L (reached after 8 hr). This was the highest concentration range (gradually increasing) that did not inhibit bacterial growth during the first 8 hr (as well as 24 hr) of incubation when compared with the untreated control grown in 1 L of BHI on a shaker. After 8 hr of incubation, 500 μl of the culture (including untreated control) was harvested for RNA isolation.
Exposure of 8-hr-old cultures to BAC
After 8 hr of incubation at 37°C in 9.9 ml of BHI on a shaker, the culture was supplemented with 100 μl of the stock solution containing BAC. The tested concentrations of BAC were 1.22 and 9.76 mg/L. After further incubation for 30 min at 37°C, 500 μl of the culture (including untreated control) was harvested for RNA isolation. A concentration of 9.76 mg/L was the highest concentration of BAC, which (during 30 min of incubation) did not reduce the number of CFU for more than 0.2 orders of magnitude (as evaluated by agar plate counting) when compared with the untreated control. In addition, this concentration did not inhibit bacteria in their further growth when compared with the untreated control.
Isolation of RNA and reverse transcription
A total of 500 μl of the BHI culture incubated as described above was added to 1 ml of RNAprotect Bacteria Reagent (Qiagen, Hilden, Germany) and incubated for 10 min at room temperature (21±1°C). Bacterial cells were then pelleted at 18,407 g for 5 min (+4°C), resuspended in 100 μl of TE Buffer (10 mmol/L Tris.Cl and 1 mmol/L EDTA, pH 8.0) containing 1 g/L lysostaphin (Sigma Aldrich), and incubated for 10 min at room temperature. After incubation, 600 μl of Lysis Buffer RA1 (Macherey-Nagel, Dueren, Germany) containing 6.0 μl of β-mercaptoethanol (Sigma-Aldrich) and 350 mg of zirconia/silica beads (Biospec Products, Bartlesville, OK) was added, and the cells were mechanically disrupted in the MagNA Lyser (Roche Diagnostics, Mannheim, Germany) at 7000 rpm for 4×20 s. RNA was isolated using the NucleoSpin RNA II kit (Macherey-Nagel) according to the manufacturer's instruction.
DNA was digested with the RNase-free DNase I recombinant as recommended by the manufacturer (Roche Diagnostics) at 37°C for 10 min, followed by inactivation at 75°C for 10 min. The RNA was purified using the NucleoSpin RNA Clean-up XS kit (Macherey-Nagel), and the final concentration of RNA was adjusted to 10 ng/μl. The RNA was checked for its integrity by agarose gel electrophoresis. RNA was reverse transcribed with the random hexamer primers (Fermentas, St. Leon-Rot, Germany) using the Omniscript RT Kit (Qiagen) according to the manufacturer's instruction. The cDNA was purified using the MinElute PCR Purification Kit (Qiagen) according to the manufacturer's instruction.
Real-time quantitative polymerase chain reaction
The primers and FAM (6-carboxyfluorescein)-labeled probes are listed in Table 1. Real-time quantitative polymerase chain reaction (RT-qPCR) was performed in a total volume of 20 μl. The primers GYR and NOR were tested at a concentration of 500 nmol/L, whereas the primers GMK and QAC at a concentration of 900 nmol/L. The final RT-qPCR mixture included the 1× DyNAmo Probe qPCR Kit (Finnzyme, Espoo, Finland), one primer pair at a concentration indicated above, 500 nmol/L of a probe, and 2 μl of cDNA template. Each RT-qPCR was performed in triplicate. Each gene was amplified in a separate well, and samples from one experiment were amplified in one run. Amplification and fluorescence detection were performed in the Light-Cycler 480 Instrument (Roche Diagnostics) using 96-well PCR plates under the following conditions: initial denaturation at 95°C for 10 min followed by 45 cycles of 95°C for 10 sec, 60°C for 30 sec, and 72°C for 1 sec. Subsequent analysis was carried out using the Fit-Point analysis option of LightCycler 480 Software Release 1.5.0 (version 1.5.0.39).
Target genes.
Reference genes.
Calculation of the relative gene expression
Relative expression of the target genes qacA and norA was normalized by geometric averaging
39
of the reference genes gyrB and gmk. The relative expression ratio (R) of the target gene is calculated based on the RT-qPCR efficiency and CP deviation of the sample versus control, and expressed in comparison to the reference genes (the efficiency-corrected calculation model based on one sample).
28
Untreated culture (i.e., the BHI culture without antimicrobial agents) was set as a control:
Etarget is the RT-qPCR efficiency of the target gene transcript; Eref (ref=1; ref=2) is the RT-qPCR efficiency of the reference gene transcripts; ΔCPtarget is the CP deviation of control–sample of the target gene transcript; ΔCPref (ref=1; ref=2) is the CP deviation of control–sample of the reference gene transcripts. The amplification rate was calculated on the basis of a linear regression slope of a dilution row 14 . Six 10-fold dilutions from 105 to 100 cDNA copies per reaction were tested for the each gene.
Results
RT-qPCR validation data
The RT-qPCR efficiency was as follows: 1.852 (slope −3.737), 1.745 (slope −4.135), 1.784 (slope −3.978), and 1.771 (slope −4.028) for the qacA, norA, gmk, and gyrB genes, respectively.
Gene expression after 8 hr of growth in the presence of BAC and CHX
R values of ≥2.0 or ≤0.5 (i.e., ∣ log2(R)∣ ≥1) represent significant differences in the relative expression ratio. 41 Because our ΔCP standard deviations sometimes increased up to 0.6 (data not shown), we considered a value of ∣ log2(R)∣ ≥2 as a significant difference in the expression ratio. From this point of view, it could be therefore concluded that increased expression of the qacA gene observed in the presence of BAC alone was near the limit of significance (Figs. 2 and 3). CHX alone only slightly increased the qacA expression; however, when added to BAC, expression of the qacA gene was higher than that observed in BAC alone (Fig. 3). Changes in the norA expression were insignificant regardless of the conditions tested.
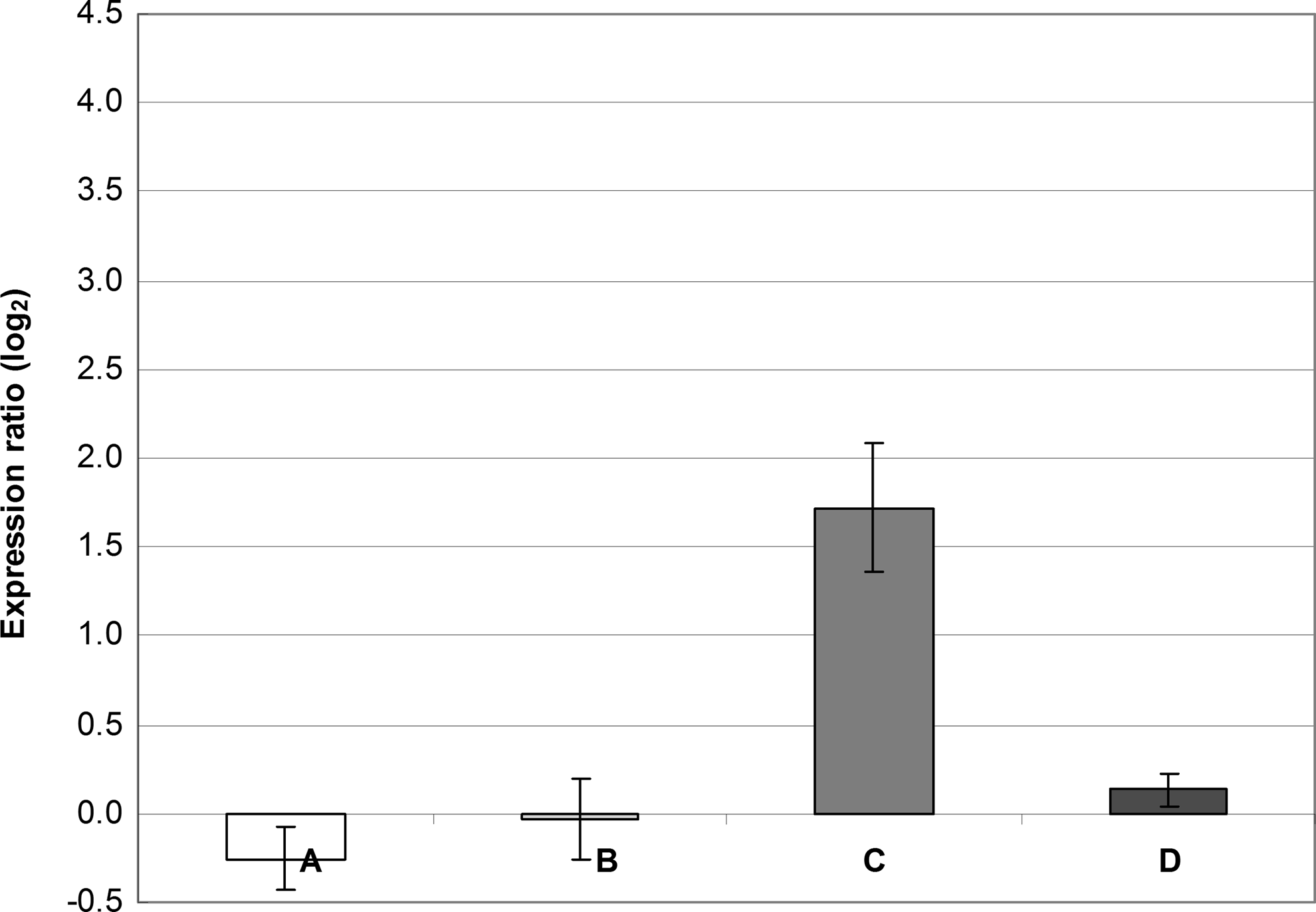
Gene expression of norA and qacA in S. aureus SK982 grown in the presence of either benzalkonium chloride (BAC; 1.22 mg/L) or chlorhexidine (CHX; 0.31 mg/L). The values of norA and qacA expression observed in BHI alone (the basal expression) were set at zero. The mean values with standard deviations (error bars) calculated from the three independent experiments are shown. (A) Expression of norA in the presence of BAC. (B) Expression of norA in the presence of CHX. (C) Expression of qacA in the presence of BAC. (D) Expression of qacA in the presence of CHX.
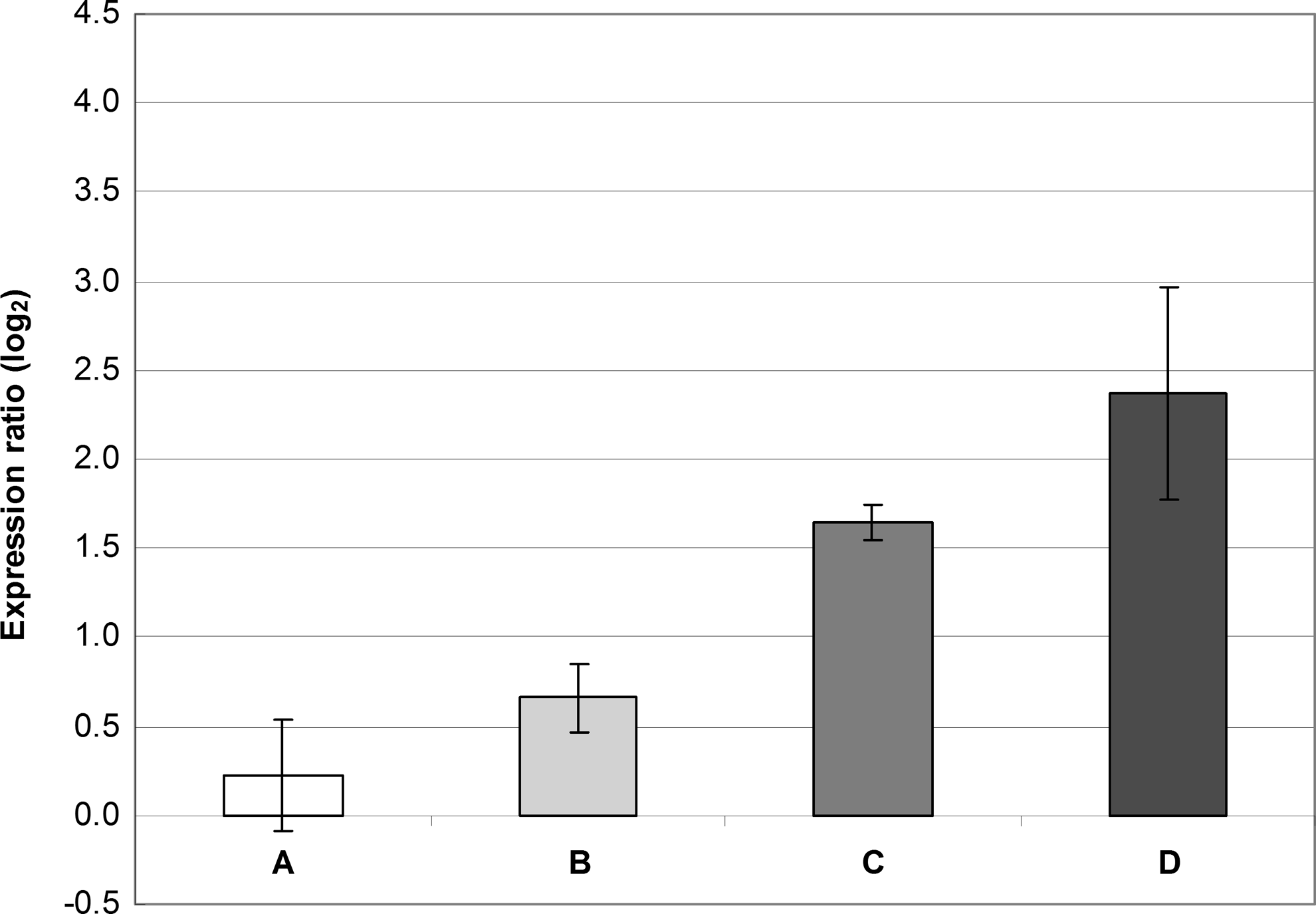
Gene expression of norA and qacA in S. aureus SK982 grown in the presence of BAC (1.22 mg/L) alone or combined with CHX (0.31 mg/L). The values of norA and qacA expression observed in BHI alone (the basal expression) were set at zero. The mean values with standard deviations (error bars) calculated from the three independent experiments are shown. (A) Expression of norA in the presence of BAC alone. (B) Expression of norA in the presence of BAC combined with CHX. (C) Expression of qacA in the presence of BAC alone. (D) Expression of qacA in the presence of BAC combined with CHX.
Gene expression after 8 hr of growth in the presence of gradually increasing concentrations of BAC
When incubated in gradually increasing concentrations of BAC, S. aureus SK982 tolerated up to 4.88 mg/L of this antiseptic. However, expression of qacA was lower than that observed in the stable BAC concentration of 1.22 mg/L and thus insignificant. Expression of norA was similar to that of qacA (Fig. 4).
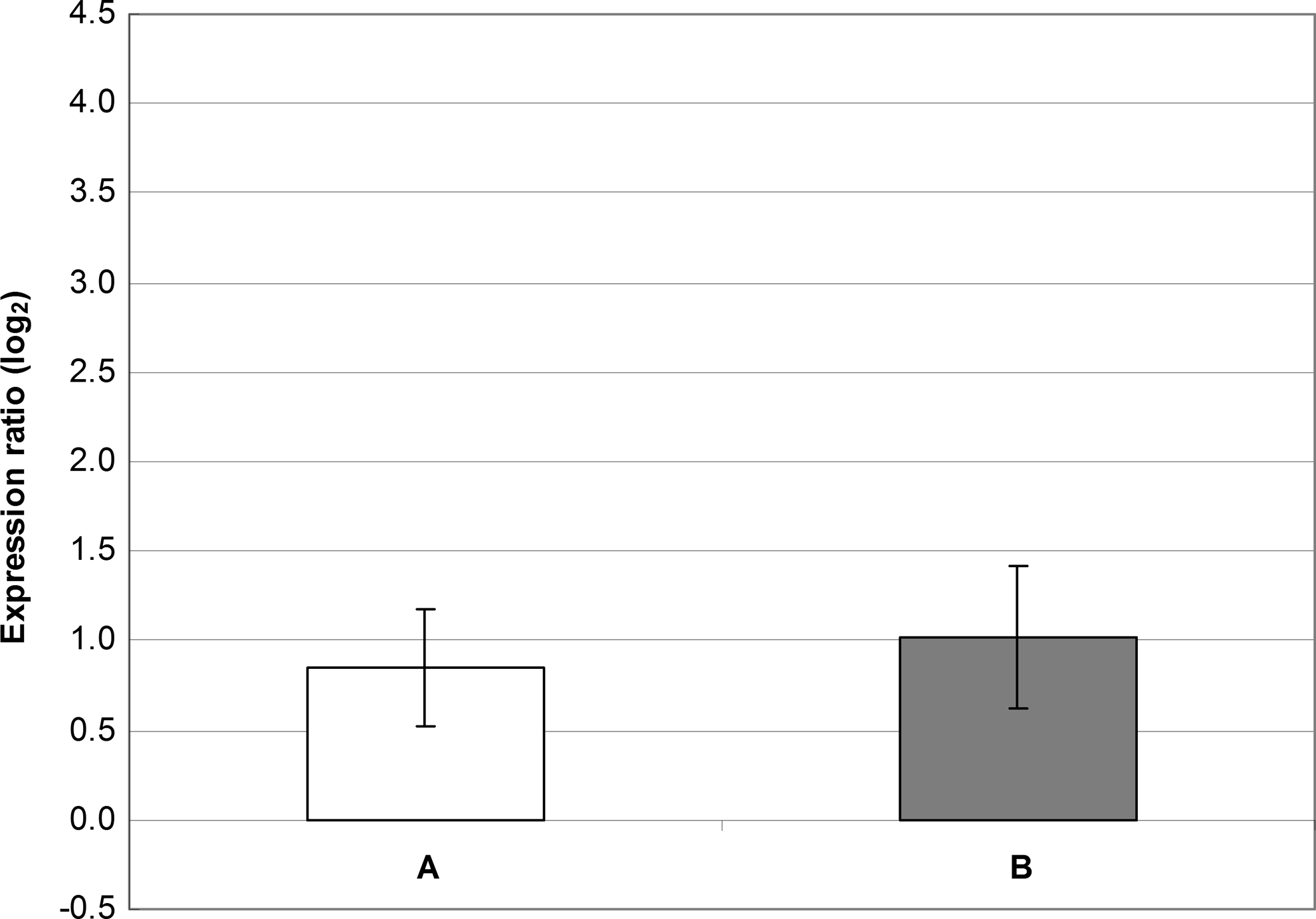
Gene expression of norA and qacA in S. aureus SK982 grown in the presence of BAC whose concentrations gradually increased (up to 4.88 mg/L) during the first 8 hr of incubation. The values of norA and qacA expression observed in BHI alone (the basal expression) were set at zero. The mean values with standard deviations (error bars) calculated from the three independent experiments are shown. (A) Expression of norA in the presence of gradually increasing concentrations of BAC. (B) Expression of qacA in the presence of gradually increasing concentrations of BAC.
Gene expression of 8-hr-old cultures exposed to BAC
Significantly increased expression of the qacA gene was observed when bacteria in the exponential phase of growth were exposed to 9.76 mg/L of BAC. Even though it cannot be excluded that such increased expression of qacA is only transient, it should be remarked that, under our study conditions, 9.76 mg/L of BAC did not inhibit bacteria in their further growth, and after 24 hr of incubation, OD605 and CFU were comparable with those observed in the untreated control. The highest concentration of BAC (1.22 mg/L) that allowed bacterial growth during the first 8 hr of incubation only slightly increased the qacA expression. This suggests a dose-dependent induction of the transcription of qacA. Changes in the norA expression were insignificant regardless of the concentrations tested (Fig. 5).
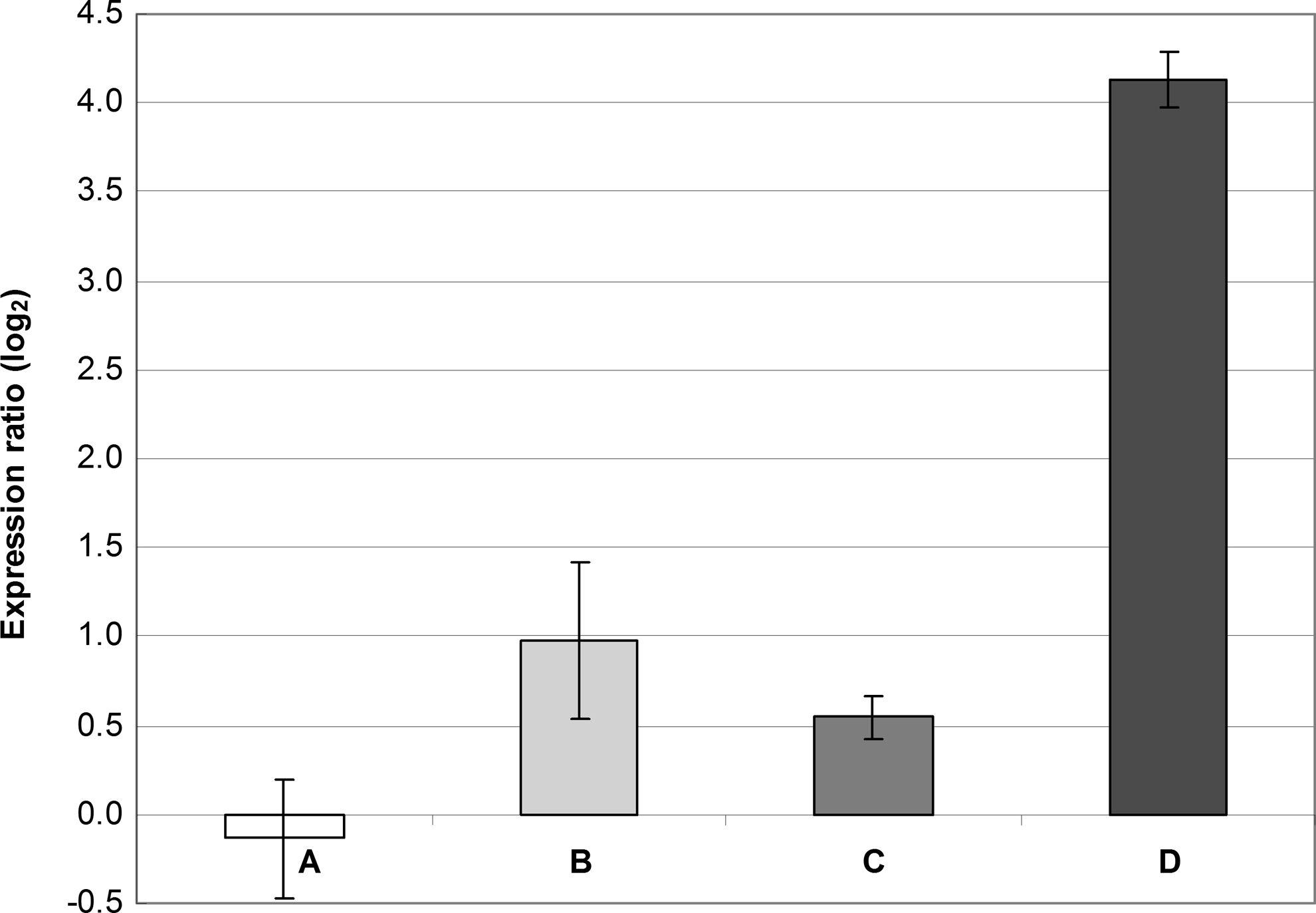
Gene expression of norA and qacA in S. aureus SK982 exposed to BAC in the exponential phase of growth (i.e., after 8 hr of incubation). The values of norA and qacA expression observed in BHI alone (the basal expression) were set at zero. The mean values with standard deviations (error bars) calculated from the three independent experiments are shown. (A) Expression of norA exposed to 1.22 mg/L of BAC. (B) Expression of norA exposed to 9.76 mg/L of BAC. (C) Expression of qacA exposed to 1.22 mg/L of BAC. (D) Expression of qacA exposed to 9.76 mg/L of BAC.
Discussion
Although the qac genes have been considered to be important genetic determinants of resistance to QACs, and thus monitored by many authors (e.g., Bischoff et al. 4 and Bjorland et al. 5 ), there is still a gap of knowledge regarding their practical significance. Low or no differences in the susceptibility to QACs between qac-positive and qac-negative bacteria of various species have been reported by several authors.16,19,34 Romao et al. 29 even demonstrated that the qacEΔ1 gene does not play an important role in resistance to BAC in Pseudomonas aeruginosa. One of the explanations could lie in the fact that qac-mediated resistance is targeted to a broad spectrum of distinct cationic compounds to the exclusion of a high-level specificity to QACs.2,11 Moreover, qac-mediated resistance could be affected by specific conditions, under which bacterial cells are exposed to the qac substrates. The latter possibility was analyzed in the current study, in which relative expression of qacA was monitored under different circumstances.
For studies of relative gene expression in staphylococci, the reference genes gyrB and gmk have been already used.1,7 The selection of reference genes is still a topic of many discussions. Contradictory findings on suitable reference genes have been reported, as well as expression of reference genes depends on conditions tested and can vary during time.10,37,38 To compensate a possible variability in the expression of reference genes during time, we compared expression profiles at the same time point of incubation. False changes in the expression of target genes due to potentially increased or decreased expression of reference genes were corrected by untreated control, which was set up as a zero expression.
We observed that 2.44 mg/L of BAC inhibited the growth of S. aureus SK982 at least for the first 8 hr of incubation, although 24-hr-old cultures reached the optical density similar to that observed in the untreated control (data not shown). The concentration of BAC that inhibited bacterial growth for 24 hr (OD605 did not increase during this period) was 4.88 mg/L. This indicates that development of the resistance to BAC is driven by the capability to adapt to this antimicrobial. In S. aureus carrying the qac genes, a stepwise adaption to twofold increasing concentrations of QACs was recently described by Smith et al. 34 This correlates with the former observation that although resistance to QACs is generally low among qac-positive staphylococci, the first resistant strains were isolated after their exposure to a QAC disinfectant. 35 Furthermore, Heir et al. 13 demonstrated that under the pressure of sublethal concentrations of BAC, qacG-containing staphylococci adapted to this antiseptic; however, their isogenic qacG-negative (QAC sensitive) variants developed efflux-based increased cross-tolerance to several other compounds, including BAC.
To evaluate the capability of S. aureus SK982 to adapt to QACs during a short-term period, it was grown in the presence of BAC whose concentrations gradually increased during the first 8 hr. Under such conditions, the strain tolerated 4.88 mg/L of BAC (the concentration that normally inhibited bacterial growth if added in the lag-phase), and after 24 hr of incubation, OD605 and CFU were comparable between the treated and untreated cultures. Therefore, there is no doubt that a certain level of adaption to BAC occurred. However, the level of qacA transcription was surprisingly lower than that observed in the stable BAC concentration of 1.22 mg/L. In other words, although S. aureus SK982 tolerated higher concentrations of BAC when gradually exposed to this antiseptic, expression of the qacA gene was found to be relatively low. This indicates that mechanisms other than those mediated by the qac genes may play an important role in the short-term adaption to BAC. McDonnell and Russell 24 demonstrated that cell wall modifications may be involved in the development of bacterial resistance to biocides. However, further investigation in this field needs to be performed.
On the other hand, in spite of the fact that stable BAC concentration of 4.88 mg/L inhibited bacterial growth for 24 hr, 9.76 mg/L of BAC did not inhibit bacteria when added in their exponential phase of growth, and overnight cultures had OD605 and CFU comparable to those of the untreated control. From this point of view, it appears that under the short-term conditions, the phase of bacterial growth is more important for the expression of QAC resistance than the capability to adapt to these antiseptics. It should also be taken into account that an increased number of bacteria in the exponential phase of growth could have negatively affected the efficacy of BAC. However, unlike disinfectants that are unstable in the presence of organic matter, such as chlorines and peroxides, QACs are generally considered as stable agents predominantly acting on the cytoplasmic membrane, causing its disorganization and cell lysis. 24 Therefore, a more probable explanation of a potentially lower efficacy of BAC in cultures exposed in the exponential phase of growth could lie in an increased probability of selection of spontaneous mutants expressing increased resistance to this antiseptic. In any way, a significant overexpression of qacA observed under such conditions may indicate its involvement in the growth-phase-dependent resistance to BAC. The finding that resistance of bacteria depends on their growth was also observed by Berney et al. 3 ; however, the authors reported the opposite phenomenon; that is, resistance to physical disinfection decreased in the fast growing cells.
Another phenomenon observed in the cultures exposed to BAC in the exponential phase of growth was relatively low expression of qacA under the pressure of 1.22 mg/L of this antiseptic (approximately three times lower than that observed in cultures exposed to 1.22 mg/L of BAC in the lag phase; Figs. 2, 3, and 5). This may suggest that in nonadapted bacterial cells (i.e., those exposed in the exponential phase of growth), other factors than qacA are also involved in the bacterial tolerance; that is, the pressure of 1.22 mg/L of BAC induced only low increase in qacA expression comparing to the cultures that were cultivated for 8 hr in the presence of the same antiseptic concentration.
Russell et al. 31 previously demonstrated that adaption to CHX may increase resistance to QACs. Similarly, in the study of Smith et al., 34 substantially increased transcription of the qacA gene was observed in the presence of CHX. In the current study, CHX increased the qacA expression when combined with BAC (Fig. 3), suggesting that these two chemicals may act in a synergistic fashion. However, this finding was not reflected in the phenotype, as tolerance to BAC did not increase in the presence of CHX.
The role of the nor genes in mediating resistance to QACs has not yet been fully understood. In the current study, no significant overexpression of norA was observed regardless of the conditions tested. Nevertheless, increased cumulative norA expression was previously demonstrated in the presence of BAC using the β-galactosidase assay. 18 Furthermore, DeMarco et al. 9 tested various agents, including QACs, and found that among S. aureus multidrug efflux genes, the nor genes were most frequently overexpressed. The authors, however, did not specify whether the QAC agents belonged to those that increased the nor expression, but, interestingly, they reported no overexpression among the qac genes. This may suggest a possible interference between the nor and qac genes as already discussed by Theis et al. 37
In conclusion, resistance to BAC is strongly affected by conditions under which bacteria are exposed to this antiseptic. The mere presence of the qac genes does not necessarily imply increased resistance that could be relevant for practice. However, the qac genes seem to be most involved in resistance to BAC in bacteria exposed to this antiseptic in the exponential phase of growth.
Footnotes
Acknowledgments
The authors thank Mgr. Maria Vass, Ph.D., for English proofreading. The work was supported by the Ministry of Agriculture of the Czech Republic (project no. MZe0002716202) and by the Ministry of Education, Youth and Sports of the Czech Republic (projects no. 2B08074 and CZ.1.05/2.1.00/01.0006-ED0006/01/01-AdmireVet).
Author Disclosure Statement
All authors report no conflicts of interest relevant to this article.