
Abstract
Abstract
Salmonella enterica is a prevalent foodborne pathogen that can carry multidrug resistance (MDR) and pose a threat to human health. Identifying the genetics associated with MDR in Salmonella isolated from animals, foods, and humans can help determine sources of MDR in food animals and their impact on humans. S. enterica serovars most frequently carrying MDR from healthy animals, retail meats, and human infections in the United States and Canada were identified and isolates resistant to the largest number of antimicrobials were chosen. Isolates were from U.S. slaughter (n=12), retail (9), and humans (9), and Canadian slaughter (9), retail (9), and humans (8; total n=56). These isolates were assayed by microarray for antimicrobial resistance and MDR plasmid genes. Genes detected encoded resistance to aminoglycosides (alleles of aac, aad, aph, strA/B); beta-lactams (blaTEM, blaCMY, blaPSE-1); chloramphenicol (cat, flo, cmlA); sulfamethoxazole (sulI); tetracycline (tet(A, B, C, D) and tetR); and trimethoprim (dfrA). Hybridization with IncA/C plasmid gene probes indicated that 27/56 isolates carried one of these plasmids; however, they differed in several variable regions. Cluster analysis based on genes detected separated most of the isolates into two groups, one with IncA/C plasmids and one without IncA/C plasmids. Other plasmid replicons were detected in all but one isolate, and included I1 (25/56), N (23/56), and FIB (10/56). The presence of different mobile elements along with similar resistance genes suggest that these genetic elements may acquire similar resistance cassettes, and serve as multiple sources for MDR in Salmonella from food animals, retail meats, and human infections.
Introduction
Multidrug-resistant (MDR) Salmonella are frequently encountered and may reduce the effectiveness of treatments. 21 In 2007, 13% of Salmonella isolated from chickens and swine during slaughter (abattoir) and 3% isolated from retail chicken in Canada were resistant to five or more antimicrobials. Almost 10% of Salmonella isolated in Canada from humans between 2004 and 2006 also exhibited resistance to five or more antimicrobials (http://phac-aspc.gc.ca/cipars-picra/index-eng.php). In 2007, 10% of Salmonella isolated in the United States from chicken, turkey, cattle, and swine slaughter samples were resistant to five or more antimicrobials, while 5.6% of retail meat isolates exhibited similar resistances. In U.S. human isolates, 6.9% were resistant to five or more antimicrobials (http://fda.gov/AnimalVeterinary/SafetyHealth/AntimicrobialResistance/NationalAntimicrobialResistanceMonitoringSystem/ucm209340.htm).
Plasmids are often associated with antimicrobial resistance in Salmonella and may also contain additional genes that provide heavy metal or sanitizer resistance, or that aid in virulence and environmental adaptability.8,21,24,29,30 These plasmids are small, circular pieces of DNA that are often self-transmissible to other bacteria through conjugation. The transfer of plasmids from one bacterial cell to another results in the horizontal transfer of genetic material, allowing acquisition of multiple genes by the recipient.10,27,30 Plasmids with the IncA/C replicon have been linked to multiple drug resistance in S. enterica associated with food animals as well as the fish pathogen Yersinia ruckeri and Y. pestis, the causative agent of plague. 42 In addition to IncA/C, several other plasmid replicons have been associated with MDR Salmonella, including B/O, HI1, HI2, I1, N, F, and P, many of which are found to be co-resident in some MDR Salmonella. 36
This study was conducted by NARMS and CIPARS to identify the antimicrobial resistance genes and genetic elements associated with MDR in Salmonella isolated from humans, animals, and retail meats. Genetic analyses included detection of antimicrobial resistance and MDR plasmid genes by DNA microarrays and PCR detection of plasmid replicons.9,23 The detailed analysis of a collection of Salmonella from the most frequently isolated MDR serovars identified antimicrobial resistance and MDR plasmid genes present in U.S. and Canadian isolates. Cluster and linkage analysis of this data was used to identify isolates with significantly similar profiles of antimicrobial resistance and plasmid genes from the different surveillance programs. Several genetic commonalities such as the presence of IncA/C plasmids were detected in MDR Salmonella isolated from different sources, including humans, retail meat, and animals at slaughter. Comparable MDR genetics were also identified in isolates from similar animal, meat, and human sources sampled by the U.S. and Canadian surveillance programs.
Materials and Methods
Isolate selection, culture conditions, and antimicrobial susceptibility testing
S. enterica isolates chosen for this study were from the U.S. Food Safety and Inspection Service (slaughter isolates, n=12), the FDA-CVM (retail isolates, n=9), the CDC (human isolates, n=9), the CIPARS (slaughter isolates, n=9; retail isolates, n=9) and the Public Health Agency of Canada (human isolates, n=8). The S. enterica serovars chosen for the study were the most frequently isolated nontyphoidal serovars carrying MDR found in each source. For U.S. NARMS this was Salmonella serovars Typhimurium, Heidelberg, and Newport for both human (CDC1–CDC9) and animal (USDA1–USDA12) isolates. U.S. retail meat isolates (FDA1–FDA9) were chosen from Salmonella serovars Typhimurium, Heidelberg, and Hadar. Isolates from Canadian CIPARS were serovars Typhimurium, Heidelberg, and Enteritidis from humans (CH1–CH8), Typhimurium, Heidelberg, Hadar, and Saintpaul, from animals (CA1–CA9), and serovars Typhimurium, Heidleberg, and Hadar from retail meat (CR1–CR9). Isolates resistant to the greatest number of antimicrobial compounds were chosen from the serovars selected for the respective agency. Isolates were then chosen from the serovars that were resistant to the maximum number of antimicrobial compounds. Isolates were obtained, cultivated, maintained, and stored as frozen stock cultures using standard methods. Culture media was obtained from Difco™ (Becton Dickinson and Company, Sparks, MD). All Salmonella isolates were subjected to testing via the Sensititre™ semi-automated antimicrobial susceptibility system following the manufacturer's instructions (TREK Diagnostic Systems, Inc., Westlake, OH) for susceptibility to amikacin, gentamicin, kanamycin, streptomycin, ampicillin, amoxicillin-clavulanic acid, ceftiofur, ceftriaxone, cefoxitin, sulfamethoxazole/sulfisoxazole, trimethoprim-sulfamethoxazole, chloramphenicol, ciprofloxacin, nalidixic acid, and tetracycline. Prevalence data, detailed isolation, testing methods, and breakpoints used to determine resistance are available in U.S. NARMS reports: http://fda.gov/AnimalVeterinary/SafetyHealth/AntimicrobialResistance/NationalAntimicrobialResistanceMonitoringSystem/ucm209340.htm, and Canadian CIPARS reports: http://www.phac-aspc.gc.ca/cipars-picra/pubs-eng.php
Microarray design and construction
The DNA microarrays and methods used for this analysis have been previously reported and validated.23,31,32 Briefly, the DNA microarrays consisted of 1,267 gene probes. Of those probes, 775 were designed to detect antimicrobial resistance genes found in the National Center for Biotechnology Information database. The other 487 gene probes were designed to detect plasmid genes identified in six strains of bacteria.31,32 IncA/C gene probes on the microarray were designed from plasmid sequences identified in Y. ruckeri str. YR71 pYR1, Y. pestis biovar Orientalis str. IP275 pIP1202, Photobacterium damselae subsp. piscicida pP99-018, Salmonella enterica subsp. enterica serovar Newport str. SL254 pSN254, Photobacterium damselae subsp. piscicida pP91278, and Escherichia coli p1658/97. Gene sequences from the plasmid pHCM1 found in Salmonella enterica subsp. enterica serovar Typhi str. CT18 were used to design probes for the genes in this IncHI1 plasmid. The microarrays were constructed by spotting probes onto Corning UltraGAPS amino-silane-coated slides (Corning, Inc., Life Sciences, Acton, MA) with an Qarray mini robot (Genetix, Hampshire, United Kingdom) and postprocessed following the manufacturer's recommendations (Corning, Inc.) as previously described.31,32
DNA isolation, labeling, microarray hybridization, and scoring
Total DNA from the isolates was prepared with the GenElute Bacterial Genomic DNA kit (Sigma, St. Louis, MO) following instructions for Gram-negative bacteria from 5 ml of overnight cultures grown in Luaria-Bertani (LB) broth at 37°C with shaking as previously described. Cye dye-labeled dCTP (Amersham, Piscataway, NJ), Klenow fragment (New England Biolabs, Beverly, MA) and random primers were used to label Salmonella genomic DNA overnight in 37°C water bath. 23 Labeled DNA was purified using Qiagen PCR clean-up kit (Qiagen, Valencia, CA), and hybridized to the microarray over night at 42°C in a Corning hybridization chamber as previously described. Microarrays were washed following the manufacturer's protocol for hybridization with formamide buffer (Corning, Inc.). Slides were scanned using ScanArray Lite microarray analysis system and images were analyzed using the ScanArray Express software version 1.1 (Packard BioChip Technologies, Billerica, MA). Positive hybridizations were determined and scored as previously described.23,32
Plasmid replicon typing
Three panels of multiplex PCR were used to determine the presence of 18 plasmid replicons commonly found in Enterobacteriaceae. PCR primers and parameters were previously described.9,30
Cluster analysis of isolates
Relationships between isolates based on gene content were determined by hierarchical cluster analysis based on antimicrobial resistance and plasmid genes detected by microarray analysis. Open source CLUSTER 3.0 was used for this analysis with Euclidean distance for gene content.13,16 Dendrogram was viewed using Java TreeView version 1.1.4r3 37 (http://jtreeview.sourceforge.net).
Statistical analysis
Linkage disequilibrium (LD) was calculated as an extension of Fisher's exact probability test on contingency tables 39 as instituted by the program Arlequin. 17 Standard settings were used, 10,000 steps in the Markov chain and 1,000 dememorization steps; and calculations of D, D′, and r2 coefficients were made with a significance level of 0.05.
Results
Detection of antimicrobial resistance and plasmid genes by microarray analysis
Resistance genes consistent with the observed phenotypes were detected in the isolates. A summary of the genes detected most frequently in the isolates is in Fig. 1 (complete hybridization data available in Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr); an exception was isolate CH1, where only a strB gene was detected. The most prevalent aminoglycoside resistance genes detected were alleles of aac(3) (24/56), aac(6′) (22/56), aadA (34/56), aadB (27/56), aadE (14/56), aph (21/56), and strAB (39/56). Detected β-lactam resistance associated genes included blaTEM (32/56), blaCMY-2 (30/56), blaPSE-1 (36/56), and the β-lactamase resistance transcriptional regulatory gene ampR (5/56). Chloramphenicol resistance genes included cat (36/56), floR (27/56), and cmlA (7/56). Sulfamethoxazole resistance genes in the isolates include sulI (26/56) and sulII (23/56). Frequently detected tetracycline-resistance-associated genes were tet(A) (45/56), tet(B) (8/56), tet(C) (8/56), tet(D) (7/56), and the regulatory gene tetR (50/56). Also detected in several isolates were alleles of trimethoprim resistance genes, such as dfrA (18/56).
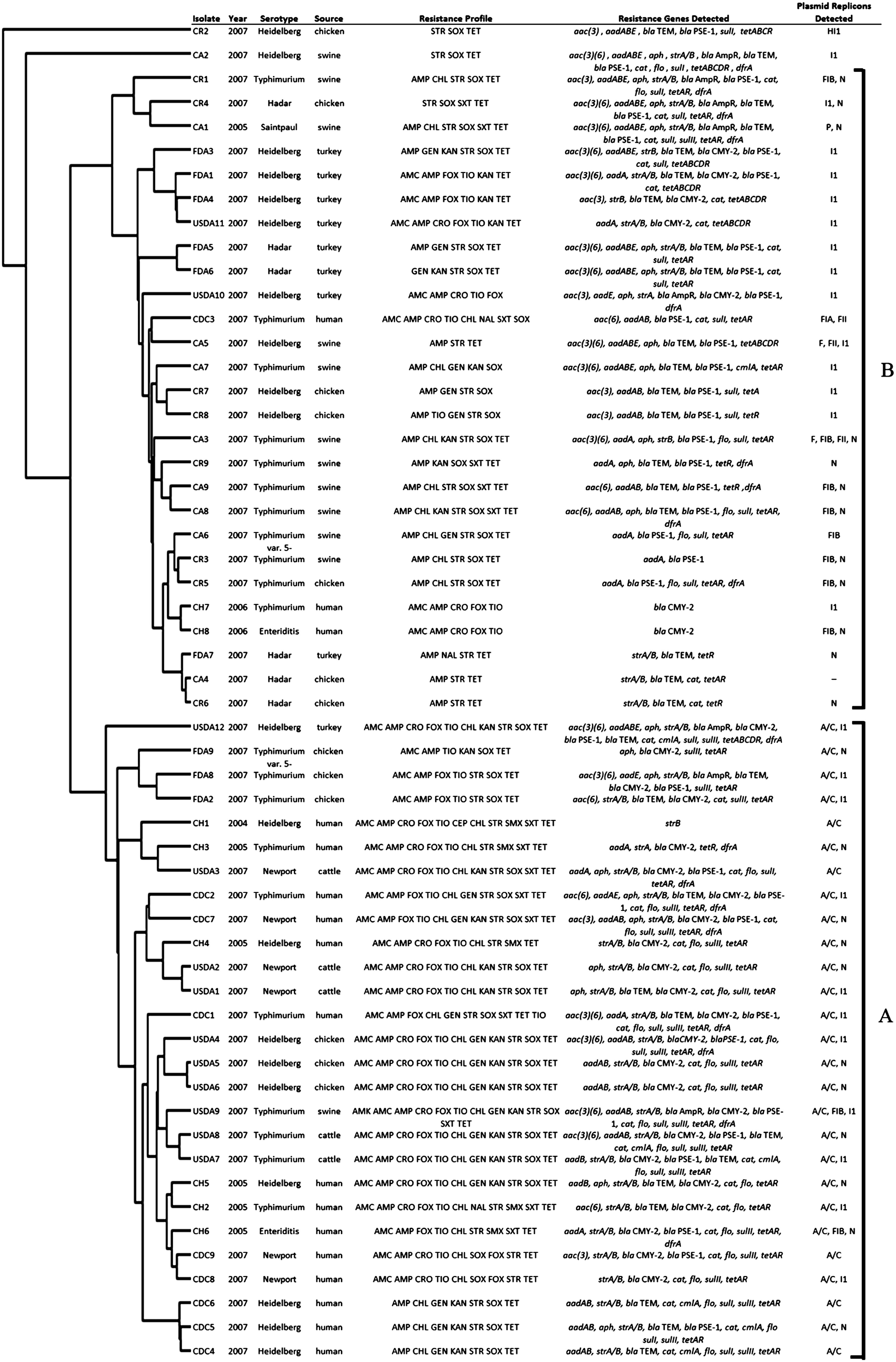
Cluster analysis of Salmonella enterica isolates based on results of microarray detection of antimicrobial resistance and plasmid genes. Isolate names designate the following: Centers for Disease Control and Prevention, isolates from U.S. human illness; U.S. Food and Drug Administration, isolates from U.S. retail meats; U.S. Department of Agriculture, isolates from U.S. animals at slaughter; CH, isolates from Canadian human illness; CR, isolates from Canadian retail meats; CA, isolates from Canadian retail meats. The IncA/C-positive cluster, group A, and IncA/C-negative cluster, group B, are labeled and indicated with brackets. Complete cluster analysis data are available in Supplementary Fig. S1. Resistance profiles of each isolate are listed with the following abbreviations for the antimicrobials: AMC, amoxicillin-clavulanic acid; AMK, amikacin; AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; FOX, cefoxitin; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; SMX, sulfamethoxazole; STR, streptomycin; SXT, trimethoprim-sulfamethoxazole; TCY, tetracycline; TIO, ceftiofur. Antimicrobial resistance genes detected are summarized for each isolate. Genes detected are separated by commas, and genes detected with multiple probes are summarized and presented only once. Names of different alleles of closely related genes detected in an isolate are concatenated together. Full hybridization data are available in Supplementary Table S1. Replicons detected in each isolate by PCR analysis are also listed with (-), indicating that none of the replicons assayed were detected.
Many of the isolates hybridized to multiple probes for IncA/C and HI1 plasmid genes. Twenty-seven of these isolates hybridized to genes in some of the 12 core regions used to define the IncA/C plasmid backbone (Table 1).31,42 Detection of these core regions, based upon sequence data from the pYR1 and pSN254 plasmids, 42 indicate that IncA/C plasmids are likely present in these 27 isolates. IncA/C core regions were detected in eight of nine U.S. human isolates, three of nine U.S. retail meat isolates, and 10 of 12 U.S. animal isolates. Some isolates lacked regions that have been identified as the site of insertions and deletions (indels) in the family of IncA/C plasmids (Table 1).24,31,32,42 Five of eight Canadian isolates from humans contain IncA/C plasmids; however, none of the Canadian retail meat or animal isolates had enough IncA/C genes detected to indicate the presence of IncA/C plasmids. Additionally, one of the Canadian retail meat isolates, CR2, was the only isolate in the study with numerous IncHI1 plasmid genes detected (183/206) (Supplementary Table S1), most likely indicating the presence of an HI1 plasmid. 24
CDC, isolates from U.S. human illness; FDA, isolates from U.S. retail meats; USDA, isolates from U.S. food animals at slaughter; CH, isolates from Canadian human illness; CR, isolates from Canadian retail meats; CA, isolates from Canadian food animals.
IncA/C plasmid core regions are based upon the assay by Welch et al. 42 Core regions are numbered in order 1–12 around the plus strand of the IncA/C plasmid sequence. Regions are scored as present if all genes within them were detected by microarray analysis of the isolate. These regions are identified with the full hybridization data in Supplementary Table S1. Some core regions missing in the isolates (e.g., 6–7, 6–9, 6–10, and others) have been identified as regions of insertions and deletions sites (indels) in the family of IncA/C plasmids.24,31,32,42
CDC, Centers for Disease Control and Prevention; FDA, U.S. Food and Drug Administration; USDA, U.S. Department of Agriculture; nd, none detected.
Plasmid replicon typing
Replicons were detected in all isolates except Canadian animal isolate CA4. The IncA/C (27/56), I1 (25/56), N (23/56), and FIB (10/56) were the most prevalent replicons detected among all isolates (Fig. 1 and Table 1). The IncA/C replicon was detected in all of the 27 isolates where large numbers (>52) of IncA/C genes were detected by microarray analysis, supporting the presence of IncA/C plasmids in these isolates. These included U.S. human, retail and slaughter isolates, as well as Canadian human isolates. However, the IncA/C replicon was not detected in Canadian retail or animal slaughter isolates. Plasmid replicons I1 and N were widespread and detected in isolates from all sources in both countries. Replicons FII, FIA, P, F, and HI1 were present in fewer isolates. Most isolates (n=51) contained one or two replicon types, three isolates were positive for three replicon types, and a single isolate was positive for four replicons. Overall, half of these 18 replicon types associated with Enterobacteriaceae were detected in at least one of the isolates. 9
Cluster and LD analysis
Almost all of the Salmonella isolates fell into two major groups based upon the antimicrobial resistance and plasmid genes detected. These were group A (n=27) with 52–180 IncA/C plasmid genes and 5–12 IncA/C core regions detected and group B (n=27) with 52 or less IncA/C genes and one or less core regions detected (Fig. 1 and Table 1). All 27 isolates of group A were IncA/C positive via microarray analysis as well as plasmid replicon typing. Group A consists mostly of U.S. slaughter (10/12), U.S. human (8/9), and Canadian human isolates (6/8), and some U.S. retail meat isolates (3/9) were present in this cluster. Cluster B contained 27 isolates in which IncA/C was not detected by microarray analysis or replicon typing. This cluster consists mostly of Canadian retail (8/9), Canadian animal slaughter (8/9), and U.S. retail meat isolates (6/9). However, U.S. slaughter (2/12), U.S. human (1/9), and Canadian human isolates (2/8) were present in lower numbers. While IncA/C replicons were not detected in group B, replicons I1, FIB, N, P, FIA, and FII were also frequently detected in this cluster. To determine if clustering was due to the analysis of highly related strains, PFGE patterns were compared. No pulsed-field gel electrophoresis (PFGE) patterns were similar enough to indicate that clonal or highly related isolates were present in the study (data not shown). Two isolates, CR2 and CA2, did not group together or with any of the other isolates. For CR2, this was due to the detection of an almost complete HI1 plasmid, making it unique from all other isolates in the study. While isolate CA2 had several IncA/C plasmid genes detected, it did not have enough to cluster with group A, and also was not PCR positive for the IncA/C replicon; however, CA2 had too many IncA/C genes detected to cluster with group B and thus was a lone isolate.
LD analysis detected significant linkage between the isolates' plasmid replicon, serovar, agency, and animal source (cattle, chicken, swine, or turkey and their retail meat products, and also human isolates) (Fig. 2). Significant linkage often corresponded with the separation of isolates into groups A and B by the cluster analysis of the microarray data. Most notably IncA/C had significant linkage with serovar, agency, and animal source, which are reflected in clusters A and B. All six Salmonella Newport isolates were in group A and contained IncA/C, while none of the six Salmonella Hadar isolates contained IncA/C and thus were in group B, leading to LD between IncA/C and these serovars. Significant positive LD was also detected for IncA/C to human isolates, with 14 out of 17 human isolates in group A; likewise, IncA/C was linked to cattle isolates, with all five in group A. The animal or animal meat product from which the isolate was collected also demonstrated a negative association with IncA/C. Only one of the 12 swine isolates and one of nine turkey isolates contained IncA/C plasmids and were in group A, while 10 of the remaining swine and all eight remaining turkey isolates were found in group B. Animal and IncI1 were linked due to eight out of nine turkey isolates having the IncI1 replicon detected; conversely, only 4 out of 12 swine isolates had the IncI1 replicon. IncI1 also had a negative relationship with IncN attributable to only one of 23 IncN-positive isolates also had the IncI1 replicon detected. Replicon FIB showed linkage with serovar, agency, and animal due to eight isolates being Typhimurium, only one isolate was from a U.S. agency (USDA), and six were from swine. Replicon FIB was also detected in eight of the isolates found in group B, and thus had a negative association with IncA/C and group A.

Pairwise linkage disequilibrium based on nine plasmid replicon groups, serovar, agency, and animal source of Salmonella enterica serovars. (+) indicates a p value of ≤0.05 and significant linkage disequilibrium; and a (−) indicates a p value of >0.05 and no significant linkage disequilibrium. Replicon groups B/O, FIC, T, K, W, Y, FIA, X, and L/M were not present in any isolate and were subsequently removed from the analysis.
Discussion
To investigate MDR in S. enterica isolated from food animals, retail meats, and infected humans, a small sampling of the most frequently isolated serovars with the highest proportion of MDR in the United States and Canada was analyzed to determine genetic mechanisms associated with their antimicrobial resistance. Overall, many of the antimicrobial resistance genes detected in the study have been previously found in MDR isolates from the United States and Canada in other studies using similar or complementary methods.24,30,31,42 The large number of genes detected in this study and their combination with plasmid genes and plasmid replicon typing allows a better understanding of MDR in Salmonella isolates from these sources. Cluster analysis based on the genes detected divided the isolates into two large clusters: A, with IncA/C plasmids, and B, without IncA/C plasmids. The IncA/C cluster was mostly animal isolates from the U.S, and human isolates from the United States and Canada. IncA/C-negative cluster B was mostly associated with animal isolates from Canada and isolates from Canada and U.S. retail meats. This study was focused on a small sampling of MDR isolates and it is unknown if these associations would also apply to the larger population of MDR Salmonella from these sources. However, the data emphasize the influence of IncA/C plasmids on MDR in U.S. and Canadian isolates, while implicating other genetic elements also associated with MDR in Salmonella.
The resistance genes detected within group A and group B were shared at a high level, indicating the presence of common mechanisms. Antimicrobial resistance genes usually found in IncA/C plasmids [e.g., blaCMY-2, floR, aac(3), aadA, aphA1, strA/B, sulI, sulII, dfrA, tet(A), tet(B), tet(C), tet(D), and tetR] were detected in most isolates of group A.24,30,31,42 Isolates in group B had resistance genes indicative of integrons like Salmonella Genomic Island-1 (SGI-1) or other MDR plasmids [e.g., blaPSE-1, blaTEM, floR, aac(3), aadA, sulI, dfrA, tet(A/G), and tetR].24,30,34 It is also important to note that total DNA was used for this study; therefore, resistance genes detected may be located in the chromosome of the isolates. Finding the same resistance genes in groups that contain different genetic vehicles for MDR genes24,30,34 may reflect the cassette nature of many of these resistance genes, where the genes encoding MDR can be transferred to plasmids, to integrons, and/or to the chromosome resulting in different genetic elements carrying similar resistance genes. For example, many IncA/C plasmids and a variety of SGI-1 sequences have been shown to share resistance genes cassettes.24,30 In addition, IncA/C plasmids have been shown to mobilize genetic elements such as SGI-1 and antimicrobial resistance gene cassettes in-trans. 14 This type of movement could explain many of the genes found in common throughout these MDR isolates.
Certain resistance genes were also found to be widespread among these isolates and between the two clusters. For example, the blaCMY-2 gene that encodes resistance to extended spectrum cephalosporins and other beta-lactams was found in isolates from both cluster A and B.11,21,22 MDR IncA/C plasmids that carry the blaCMY-2 gene have been reported and characterized as a major contributor to MDR in Salmonella from the United States and Canada.3,11,21,22 In IncA/C-positive group A, 23/27 isolates have the blaCMY-2 gene detected. However, 7/27 isolates in group B also have the blaCMY-2 gene detected. Five of these are Salmonella Heidelberg isolates also have the IncI1 plasmid replicon detected. IncI1 plasmids carrying a blaCMY-2 gene have recently been found in Salmonella Heidelberg human isolates in the United States and Canada.19,20 Therefore, it is likely that there are two or more genetic elements carrying the blaCMY-2 genes in these MDR Salmonella isolates. The prevalence of MDR Salmonella resistant to cephalosporins due to blaCMY-2 encoding IncA/C plasmids has declined recently in animal isolates; however, the spread of a blaCMY-2 gene by other genetic elements, such as IncI1, could cause increased prevalence of Salmonella resistant to extended-spectrum cephalosporins in the future.
A large amount of variability was found in the core backbone regions of the IncA/C plasmids detected in group A. Most of the regions missing are previously described indels within regions 6–10, 1–2, and 7–12.24,31,32,42 The genes encoded in these indels have been observed to have effects on a great diversity of plasmid functions. Some of these include transmissibility of the plasmids, mobilization of other genetic elements, phenotypes conferred by the plasmids, response to selective pressures including heavy metals, chemical sanitizers, and traditional antimicrobials, as well as having phylogenetic implications.24,31,32,42 These aspects of the IncA/C plasmid family have been well characterized in other studies of Salmonella as well as other bacteria containing IncA/C plasmids.24,31–33,41,42 Interestingly, while IncA/C is associated with almost half of the MDR Salmonella isolates in this study, there is a lot of diversity within the structure of the plasmids in this small collection of isolates, possibly indicating several different phylogenetic origins of the IncA/C plasmids in these Salmonella isolates and even within a single serovar of Salmonella. This suggests that there are multiple sources of MDR IncA/C Salmonella in these isolates rather than the spread of a single IncA/C plasmid among the different MDR Salmonella found in animals, retail meat, and human infections. Alternatively, the plasmids may have spread some time ago and then undergone mutations and deletions within certain serovars or in certain environments resulting in the diversity we have observed.
Other plasmids that could potentially carry MDR genes were detected in each of the large clusters, including IncI1, as discussed above, which is known to carry blaCMY-2 or MDR in bacteria isolated from humans and animals.19,20,25,26 IncN, IncFIB, and other plasmid replicons were also detected demonstrating that the genetic elements carrying MDR, while dominantly associated with IncA/C and I1, are complex and could be associated with other plasmids and genetic elements such as integrons like SGI1.24,30,34 This observation is also supported by the two lone MDR isolates that are genetically distinct from the two large clusters, including one carrying an IncHI1 plasmid that is often associated with MDR in typhoid and paratyphoid serovars of Salmonella.35,40
Within both large clusters, isolates that grouped together into subclusters also had other similarities. The isolates were often the same serovar when isolated from the same type of animal or animal meat, or when isolated by the same agency or in the same country. This was especially true for human isolates that clustered most closely to other human isolates and in some cases human isolates from the United States and Canada clustered together. The clustering did show that 14 out of 17 human isolates were associated with IncA/C, a relationship between source and IncA/C that exhibits significant LD. This likely reflects an association between MDR human salmonellosis and Salmonella serovars, such as Newport, Heidelberg, Typhimurium, and others that carry a plasmid from the MDR IncA/C family.19,20,24 This association has been described in several reports on MDR human and animal isolates collected during this time period (2005–2007), and emphasizes the impact of IncA/C plasmids on U.S. and Canadian human MDR Salmonella infections.4,15,19–21,24,28 U.S. animal MDR Salmonella isolates were also found to be mostly IncA/C positive (10/12), as were three of the U.S. retail meat isolates. In contrast, all Canadian animal and retail meat isolates lacked the IncA/C plasmid.7,21,22,24,28,30 Canadian monitoring of cattle and retail beef had previously detected MDR Salmonella with the IncA/C plasmid; however, sampling of cattle and beef in Canada was discontinued due to the overall low prevalence of Salmonella before isolates were collected for this study.
Most human infections caused by MDR nontyphoidal Salmonella are thought to be foodborne, with animals serving as reservoirs of resistance and retail meats acting as vehicles for human disease.1,2,38,43 Agencies in the United States and Canada have developed programs to monitor the levels of antimicrobial resistance in Salmonella found in food animals, retail meats, and humans. This study investigated MDR Salmonella collected by these programs and determined the genes associated with antimicrobial resistance. Genetic elements, such as IncA/C and IncI1 plasmids were found in most of the isolates, but other plasmids and genetic elements were also associated with MDR in many of these Salmonella. These results suggest that in addition to phenotypic monitoring of antimicrobial resistance in Salmonella, investigations of the genetic elements causing resistance are helpful in understanding sources and spread of MDR and determine the impact of foodborne Salmonella on human health.
Footnotes
Acknowledgments
The authors thank Jennifer Turpin, Georgina Hidalgo, Jovita Haro, Benny Barrett, and Takiyah Ball at USDA for technical assistance. From CDC we thank Jean Whichard for helpful conversations and editing the manuscript. From CIPARS we thank Lucie Dutil for isolate selection, and Laura Martin and Emily Weir for DNA isolation. We would also like to dedicate this article in memory of our colleague, Lucie Dutil, whom we had the privilege to know and work with through NARMS and CIPARS collaborations. This work was supported by USDA project number 6612-32000-006-00.
Disclaimer
The mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture, Centers for Disease Control and Prevention, U.S. Food and Drug Administration, or PHAC.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.