
Abstract
Abstract
The increased resistance of pathogenic microorganisms is frequently attributed to the extreme and inadequate use of antibiotics and transmission of resistance within and between individuals. To counter the emergence of resistant microorganisms, considerable resources have been invested in the search for new antimicrobials. Plants synthesize a diverse array of secondary metabolites (phytochemicals) known to be involved in defense mechanisms, and in the last few years it is recognized that some of these molecules have health beneficial effects, including antimicrobial properties. In this study, the mechanism of action of gallic (GA) and ferulic (FA) acids, a hydroxybenzoic acid and a hydroxycinnamic acid, was assessed on Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, and Listeria monocytogenes. The targets of antimicrobial action were studied using different bacterial physiological indices: minimum inhibitory concentration (MIC), minimum bactericidal concentration (MBC), membrane permeabilization, intracellular potassium release, physicochemical surface properties, and surface charge. It was found that FA and GA had antimicrobial activity against the bacteria tested with MIC of 500 μg/mL for P. aeruginosa, 1500 μg/mL for E. coli, 1750 μg/mL for S. aureus, and 2000 μg/mL for L. monocytogenes with GA; 100 μg/mL for E. coli and P. aeruginosa, 1100 μg/mL and 1250 μg/mL for S. aureus and L. monocytogenes, respectively, with FA. The MBC for E. coli was 2500 μg/mL (FA) and 5000 (GA), for S. aureus was 5000 μg/mL (FA) and 5250 μg/mL (GA), for L. monocytogenes was 5300 μg/mL (FA) and 5500 μg/mL (GA), and 500 μg/mL for P. aeruginosa, with both phytochemicals. GA and FA led to irreversible changes in membrane properties (charge, intra and extracellular permeability, and physicochemical properties) through hydrophobicity changes, decrease of negative surface charge, and occurrence of local rupture or pore formation in the cell membranes with consequent leakage of essential intracellular constituents. The overall study emphasizes the potential of plant-derived molecules as a green and sustainable source of new broad spectrum antimicrobial products.
Introduction
Most of the antibiotics available today come from natural origin, particularly from microbial sources.62,66 However, plants have developed numerous chemical strategies to protect themselves from microbial attacks, including the production of bactericidal products.20,28 Diverse natural products are well known for their antimicrobial activities and are a fundamental source of chemical diversity and important components of the current pharmaceutical products.20,38,61,62
Phenolic products constitute one of the most numerous and ubiquitously distributed group of plant secondary metabolites, commonly found in diverse dietary products, particularly vegetables, fruit, chocolate, and beverages.73,87 Phenolic products are subdivided into three groups: phenolic acids, flavonoids, and tannins.60,87,88 Phenolic acids have one carboxylic acid functional and are hydroxylated derivatives of benzoic (e.g., gallic [GA], protocatechuic, and p-hydroxybenzoic acids) and cinnamic acids (e.g., caffeic, p-coumaric, and ferulic acids [FA]).60,76,90 Although the basic skeleton remains the same, the numbers and positions of the hydroxyl groups on the aromatic ring, and the type of substituents, cause significant changes on the properties of the phenolic products.60,75,76 The site and the number of hydroxyl groups on the phenol group are thought to be related to their relative toxicity to microorganisms, and are the principal structural features influencing the antioxidant capacity of phenolics, with evidence that increasing hydroxylation results in toxicity increase.15,64,68 In addition, some authors have found that more highly oxidized phenols have higher antimicrobial properties.15,64,67 The hydroxycinnamic acids have significantly higher antioxidant activity than the hydroxybenzoic acids, which can be attributed to the presence of the CH=CH-COOH group in the hydroxycinnamic acids and the COOH group in the hydroxybenzoic acids. The presence of the CH=CH-COOH groups in hydroxycinnamic acids provide higher H-donating capability and subsequent radical stabilization than the carboxylate group in hydroxybenzoic acids.34,59
Natural phenolic products can exhibit a wide range of biological effects including: antibacterial, antifungal, antiviral, anti-inflammatory, antiallergic, hepatoprotective, antithrombotic, anticarcinogenic, and vasodilatory action. Therefore, phenolics are considered potential therapeutic agents against a wide range of ailments including neurodegenerative diseases, cancer, diabetes, cardiovascular dysfunctions, inflammatory diseases, and also against aging.31,73,74 Moreover, broad clinical evidences have shown that chemoprevention by phenolics is an economical and applicable approach in chemotherapy management. 29
The aim of this work was to investigate, using several physiological indices, the mechanisms involved in the antimicrobial activity of two selected phenolic acids, FA and GA against four human pathogenic bacteria (Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, and Listeria monocytogenes) of biomedical prominence.
Materials and Methods
Bacterial strains and growth media
The following strains were used in this study: E. coli CECT 434, P. aeruginosa ATCC 10145, S. aureus CECT 976, and L. monocytogenes ATCC 15313. These bacteria were already used as model microorganisms for antimicrobial tests with phytochemical products.8,61,71 All microbial strains were stored at −80°C in cryovial and 30% (v/v) glycerol, and they were subcultured in Mueller-Hinton Agar (MHA) before testing.
Phenolic acids
GA, 3,4,5-trihydroxybenzoic acid and FA, (E)-3-(4-hydroxy-3-methoxy-phenyl)prop-2-enoic acid were obtained from Sigma-Aldrich. Phytochemicals are routinely classified as antimicrobials on the basis of susceptibility tests that produce inhibitory concentrations in the range of 100 to 1000 μg/mL.69,81 In this work, each product was tested at a concentration of 100, 500, and 1000 μg/mL in dimethyl sulfoxide (DMSO) (Sigma). The chemical structures of GA and FA are shown in Fig. 1.
Chemical structures of gallic acid (GA)
Minimum inhibitory concentration
The bacteria were grown overnight at 30°C in Mueller-Hinton Broth (MHB), from which an inoculum was taken and adjusted to an OD640nm of 0.2±0.02 (1×108 colony forming units [CFU]/mL). Subsequently, for each bacterium at least 16 wells of a sterile 96-well polystyrene microtiter plates were filled with cells (180 μL) and phytochemicals (20 μL). These were tested at three different concentrations (100, 500, and 1000 μg/mL). Cell suspensions with DMSO and cell suspension without phytochemical were used as controls. The microtiter plates were then incubated for 24 hr at 30°C in an orbital shaker (150 rpm). The absorbance was measured at 640 nm using a Microplate Reader (Spectramax M2e, Molecular Devices, Inc.). Minimum Inhibitory Concentration (MIC) of phenolic acids was determined by the microdilution method.6,9,14,16,31,47,92 The MIC was determined as the lowest concentration of phenolic acids at which no growth was detected.6,31,35,65 All tests were performed in triplicate with three repeats.
Minimum bactericidal concentration
Bacterial cells were grown overnight in batch culture using 100 mL of MHB at 30°C and 150 rpm. After the overnight growth, culture was centrifuged (3772g, 6 min), washed twice with saline solution (0.85% NaCl), and resuspended in saline solution to obtain an OD640 of 0.2±0.02 (1×108 CFU/mL). Then, an aliquot of this suspension was collected and maintained 30 min in contact with different concentrations of the phenolic acids (100, 500, and 1000 μg/mL). Subsequently, bacterial suspensions were diluted to an adequate cellular concentration (from 107 to 100) in saline solution. Then, 100 μL of each suspension was transferred onto MHA plates and incubated at 30°C. Colony enumeration was carried out after 24 hr. Cell suspensions without phytochemical were used as controls. The MBC was taken as the lowest concentration of phytochemicals at which no CFU were detected on solid medium.41,52,56 All experiments were performed in triplicate with three repeats.
Physicochemical characterization of the bacterial surfaces
Bacterial suspensions were prepared in ultrapure water (pH 6). No significant osmotic pressure effects were found when comparing the planktonic bacterial viability in water and in saline solution (0.85% NaCl), for a period of up to 150 min (p>0.05). Afterward, their physicochemical properties were determined by the sessile drop contact angle measurement on bacterial lawns, prepared as described by Busscher et al.
10
Contact angles were automatically determined using an OCA 15 Plus (DATAPHYSICS) video-based optical measuring instrument, allowing image acquisition and data analysis. Contact angle measurements (≥25 per liquid and phenolic acids concentration) were carried out according to Simões et al.
72
The liquid surface tension components reference values were obtained from the literature.
30
Hydrophobicity was evaluated after contact angle measurement, following the van Oss approach,84–86
where the degree of hydrophobicity of a given surface (s) is expressed as the free energy of interaction between two entities of that surface, when immersed in water (w)–(ΔGsws mJ m−2). If the interaction between the two entities is stronger than the interaction of each entity with water, ΔGsws<0, the material is considered hydrophobic. Conversely, if ΔGsws>0, the material is hydrophilic. ΔGsws can be calculated through the surface tension components of the interacting entities, according to the equation given below:
where γLW accounts for the Lifshitz-van der Waals component of the surface free energy and γ+ and γ− are the electron acceptor and electron donor parameters, respectively, of the Lewis acid–basef component (γAB), with
where θ is the contact angle and γTot=γLW γAB. At least three independent experiments were performed for each condition tested.
Bacterial surface charge—zeta potential
The zeta potential of bacterial suspensions, before and after contact with different GA and FA concentrations (100, 500, and 1000 μg/mL), was determined using a Nano Zetasizer (Malvern Instruments). Cell suspensions in ultrapure water (pH 6), without phytochemical, were used as controls. The zeta potential was measured by applying an electric field across the bacterial suspensions. The experiments were repeated at least three times.
Assessment of membrane integrity due to propidium iodide uptake
The Live/Dead BacLight™ kit (Invitrogen/Molecular Probes) assesses membrane integrity by selective stain exclusion. 70 This fast method was applied to estimate both viable and total counts of bacteria. BacLight is composed of two nucleic acid-binding stains: SYTO 9™ and propidium iodide (PI). SYTO 9™ penetrates bacterial membranes, staining the cells green; PI only penetrates cells with damaged membranes, binding to single and double-stranded nucleic acids. The combination of these two stains generates red fluorescing cells. After overnight growth, the cells were centrifuged (3772g, 10 min) and washed once with saline solution (0.85%). Afterward, bacteria were resuspended in saline solution to obtain an OD640 of 0.2±0.02. Then, an aliquot of 1 mL of this suspension was collected and different concentrations of phenolic acid were tested (100, 500, and 1000 μg/mL) for 30 min in contact with the bacteria. Cell suspensions without phytochemicals were used as controls. Afterward, bacteria were transferred to saline solution and diluted 1:10. Three hundred microliters of each diluted suspension was filtered through a Nucleopore® (Whatman) black polycarbonate membrane (pore size 0.22 μm) and stained with 250 mL of diluted SYTO 9™ and 250 mL of diluted component PI. The dyes were left to react for 15 min in the dark, at 27°C±3°C. The membrane was then mounted on BacLight mounting oil, as described in the manufacturer's instructions. The microscope used for the observation of stained bacteria was a LEICA DMLB2 with a mercury lamp HBO/100W/3, incorporating a color camera digital camera to acquire images using IM50 software (LEICA) and a 100× oil immersion fluorescence objective. The optical filter combination for optimal viewing of stained mounts consisted of a 480–500 nm excitation filter in combination with a 485 nm emission filter (Chroma 61000-V2 DAPI/FITC/TRITC). A program path (Scan Pro 5) involving object measurement and data output was used to obtain the total number of cells (both stains) and the number of PI-stained cells (damaged cells). Both the total number of cells and the number of PI-stained cells on each membrane were estimated from counts of ≥20 fields of view. The total number of cells counted per field of view ranged from 50 to 200 cells. Three independent experiments were performed for each condition tested.
Potassium (K+) leakage
Flame emission and atomic absorption spectroscopy were used for K+ titration in solutions of the bacteria in study treated with one concentration (1000 μg/mL) of phenolic acids. The samples were filtrated after contact with the phytochemicals and then were analyzed in a GBC AAS 932plus device using GBC Avante 1.33 software. The experiments were repeated three times.
Statistical analysis
The data were analyzed using the statistical program SPSS version 20.0 (Statistical Package for the Social Sciences). The mean and standard deviation within samples were calculated for all cases. One-way Anova with Bonferroni test was used to assess the statistical significance value (confidence level ≥95%).
Results
Inhibitory and bactericidal concentration of phenolic acids
The MIC is interpreted as the lowest concentration that inhibits visible microbial growth and is expressed in terms of μg/mL, whereas the MBC is interpreted as the lowest concentration that can completely kill the bacteria. The MIC of phenolic acids against the four bacterial strains is summarized in Table 1. The MIC of GA was 500 μg/mL for P. aeruginosa, 1500 μg/mL for E. coli, 1750 μg/mL for S. aureus, and 2000 μg/mL for L. monocytogenes. The MIC of FA for E. coli and P. aeruginosa was 100 μg/mL. FA inhibited S. aureus and L. monocytogenes cell growth at 1100 μg/mL and 1250 μg/mL, respectively. The MBC of the phenolic acids ranged between 500 and 5500 μg/mL (Table 2). The MBC of FA and GA for E. coli was 2500 μg/mL and 5000 μg/mL, respectively. The MBC of FA and GA for S. aureus was 5000 μg/mL and 5250 μg/mL, respectively. The MBC of FA for L. monocytogenes was 5300 μg/mL and 5500 μg/mL with GA. P. aeruginosa had the smallest MBC, 500 μg/mL (5×MIC), of both phenolic acids.
MIC, minimum inhibitory concentration; GA, gallic acid; FA, ferulic acid.
MBC, minimum bactericidal concentration.
Effects of phenolic acids on physicochemical surface properties of bacterial cells
The hydrophobicity, the polar and apolar components of the surface tension of the bacteria tested, without and with phenolic acids treatment, are presented in Table 3. The bacteria used in this study had hydrophilic properties (ΔGTOT>0 mJ/m2). However, with the application of phenolic acids, particularly GA (p<0.05), it is possible to observe changes in the bacterial membrane physicochemical character (Table 3). E. coli cell surface (30.3 mJ/m2) became more hydrophilic in the presence of GA (44.2 mJ/m2) (p<0.05) and FA had low effects in the surface cell properties (29.0 mJ/m2) (p>0.05). The application of both phenolic acids promoted the increase of hydrophilic character of P. aeruginosa (p<0.05). The opposite effect was observed for L. monocytogenes, that is, GA and FA induced a cell surface hydrophobic character (p<0.05). S. aureus surface became more hydrophobic when exposed to GA (p<0.05) and no alteration was observed with FA (p>0.05). The values of the surface tension components demonstrated that the Gram-positive bacteria acquired polar character after phenolic acids treatment, as reflected by an increase in γAB in comparison without phenolic acids exposure (p<0.05). The apolar component, γLW, of E. coli was almost unaffected by the exposure to both phenolics (p>0.05). For S. aureus and L. monocytogenes, the apolar component was slightly affected due to phenolic acids exposure (p<0.05). However, P. aeruginosa acquired apolar properties after exposure to GA and FA (p<0.05). The electron acceptor, γ+, component increased with phenolic acids application for both Gram-positive bacteria (p<0.05) and decreased for those Gram negative (p<0.05).
The mean±SD for at least three replicates are given.
ΔGTOT>0 mJ/m2 (hydrophilic); ΔGTOT<0 mJ/m2 (hydrophobic).
Effects of phenolic acids on bacterial surface charge
The surface charge of cells is frequently determined based on their zeta potential, which is calculated from the mobility of cells in the presence of an electrical field under defined pH and salt concentrations. The results obtained from the zeta potential measurement (Fig. 2) allowed a better understanding of how the negatively charged GA and FA interacted with bacterial surfaces. All bacterial cells tested had a negative surface charge: −17 mV for E. coli, −12.5 mV for P. aeruginosa, −23.6 for S. aureus, and −21.9 mV for L. monocytogenes. After exposure to phenolic acids, changes in the surface charge of cells to less negative values were verified, particularly for E. coli and P. aeruginosa. For these bacteria, the zeta potential values obtained after exposure to GA and FA were significantly different from the control (p<0.05). In contrast, for S. aureus the zeta potential values are similar to the control, for both phenolic acids (p>0.05). The same scenario was observed for L. monocytogenes with FA and with GA at 500 μg/mL (p>0.05).
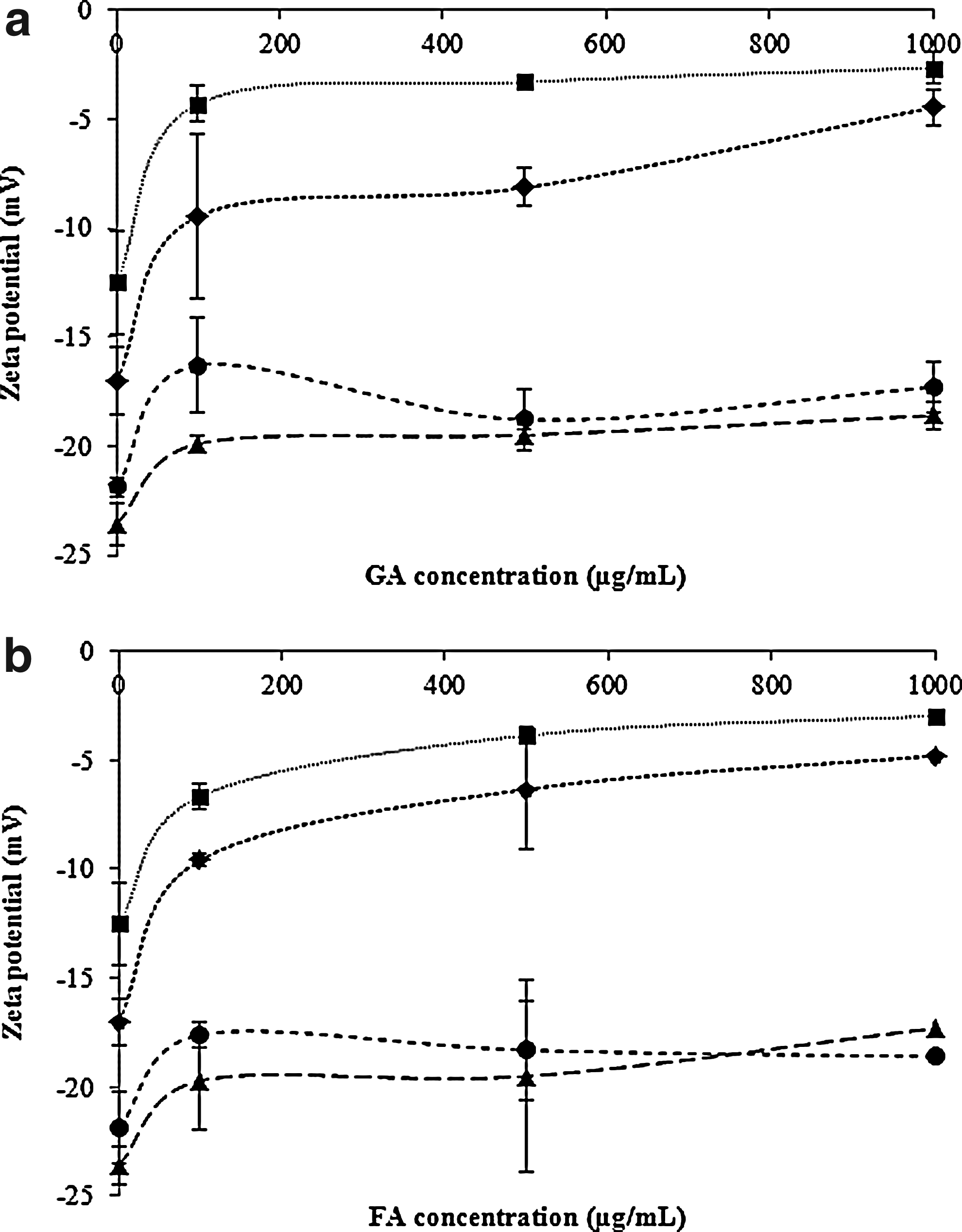
Zeta potential values (mV) of suspensions of E. coli (♦), P. aeruginosa (■), S. aureus (▴), and L. monocytogenes (●) when exposed to different concentrations (0, 100, 500, and 1000 μg/mL) of GA
Effects of phenolic acids on bacterial cytoplasmic membrane integrity
The PI uptake results suggest that GA and FA compromise the integrity of the cytoplasmic membrane (Fig. 3). It is possible to observe that the percentage of cells with membrane damaged considerably increased with phenolic acids concentration (Fig. 3). For a GA and FA concentration of 100 μg/mL, the percentage of cells of E. coli (GA−13%; FA−16%), S. aureus (GA−23%; FA−3%), and L. monocytogenes (GA−10%; FA−1%) stained with PI was reduced (Fig. 3). However, a concentration of 100 μg/mL of both phenolic acids was sufficient to damage the cytoplasmic membrane of about 60% of the P. aeruginosa cells. A concentration of 500 μg/mL significantly increased the membrane damage for several bacteria (p<0.05), with both phenolic acids. For FA and GA at 1000 μg/mL, the percentage of cells of P. aeruginosa, S. aureus, and L. monocytogenes stained with PI was ∼100%. For E. coli exposed to 1000 μg/mL the damage in cytoplasmic membrane was about 62% (GA) and 80% (FA) of the total cells.

Permeability of E. coli, P. aeruginosa, S. aureus, and L. monocytogenes to propidium iodide (PI) after treatment with GA ), 500 ( ), and 1000 (■) μg/mL) for 30 min. The percentage of cells nonstained with PI corresponds to the fraction of viable cells. The mean±SD for at least three replicates is illustrated.
), and 1000 (■) μg/mL) for 30 min. The percentage of cells nonstained with PI corresponds to the fraction of viable cells. The mean±SD for at least three replicates is illustrated.
Effects of phenolic acids in intracellular potassium release
Table 4 shows the effects of phenolic acids on K+ release by E. coli, P. aeruginosa, S. aureus, and L. monocytogenes. The untreated bacterial controls showed residual loss of intracellular K+. Intracellular contents were released due to exposure of the cells to 1000 μg/mL of GA and FA for 30 min. This indicates alteration in the cytoplasmic membrane permeability. For S. aureus, no significant K+ released was found due to phytochemicals exposure (p>0.05). In fact, the release of K+ by Gram-positive bacteria was less than for the Gram negative (p<0.05).
K+
The mean±SD for at least three replicates is illustrated.
Discussion
In recent years, drug resistance of human pathogenic bacteria has been extensively reported.3,18,19 Therefore, efforts are being made to discover efficient antimicrobial molecules. Phytochemicals are excellent candidates for antimicrobial chemotherapy.3,16,57,58,65 In the present study, the activity and mode of action of phenolic acids against four bacterial strains (E. coli, P. aeruginosa, S. aureus, and L. monocytogenes) with pathogenic potential were characterized. Different bacterial physiological indexes were studied such as MIC, MBC, PI uptake, intracellular potassium release, surface hydrophobicity, and surface charge. P. aeruginosa was more susceptible bacteria to GA with MIC at 500 μg/mL. This bacterium and E. coli were more susceptible to FA with MIC at 100 μg/mL. The Gram-positive bacteria were less susceptible to both phenolic acids with MIC of 1750 μg/mL (GA)/1100 μg/mL (FA) and 2000 μg/mL (GA)/1250 μg/mL (FA) for S. aureus and L. monocytogenes, respectively. Other studies reported MIC of 490 μg/mL (GA) and 160 μg/mL (FA) for E. coli AW1.7. 65 An MIC of 533 μg/mL for GA against P. aeruginosa ATCC 27853 and S. aureus ATCC 29213 has been reported. Jayaraman et al. 31 obtained an MIC of 2000 μg/mL with GA for five strains of P. aeruginosa including clinical isolates. In a study performed by Hayrapetyan et al., 26 GA at a concentration of 512 μg/mL showed no inhibition of L. monocytogenes ATCC 23074 and ATCC 35152. For L. monocytogenes ATCC 7644 the MIC of GA was reported as 1600 μg/mL. 79 The application of different phenolic products (tea flavonoids) to Streptococcus mutans and Streptococcus sobrinus were inhibitory at concentrations between 250 μg/mL and 1000 μg/mL. 91 Sánchez-Maldonado et al. 65 reported the MIC values of hydroxybenzoic, hydroxycinnamic, and methoxy-benzoic acids (caffeic, p-coumaric, FA, GA, protocatechuic, chlorogenic, cinnamic, syringic, benzoic, dihydrocaffeic and phloretic acids, and catechol) for three Gram-positive bacteria (Lactobacillus plantarum, Lactobacillus hammesii, and Bacillus subtilis) and for E. coli AW1.7. These MIC values are in the range of those found in the present study. The authors found that the Gram-positive bacteria were generally more resistant than the E. coli strain, and that the increase in the number of hydroxyl groups increased the growth inhibitory activity of both hydroxybenzoic and hydroxycinnamic acids. The substitution of hydroxyl groups with methoxy groups increased the activity of hydroxybenzoic acids, but not of hydroxycinnamic. 65 It is important to take into account that the discrepancy in methods for determination of antimicrobial activity combined with the use of different microorganisms and strains can help to explain the different MIC of FA and GA observed in the diverse studies. Further, it is known that no strain can represent the behavior of a species. 23
The bactericidal concentration of both phenolic acids was 500 μg/mL for P. aeruginosa. Moreover, the MBC was 5000 μg/mL for E. coli with GA, and for S. aureus with FA. The MBC for E. coli with FA was 2500 μg/mL. S. aureus with GA (5250 μg/mL) and L. monocytogenes with both phytochemicals (GA–5500 μg/mL; FA–5300 μg/mL) were the most resistant microorganisms. The values of MBC obtained are in range of those described in other studies. In a work performed by Ellnain-Wojtaszek and Mirska, 21 the MBC of FA against E. coli and S. aureus was 1250 μg/mL and 2500 μg/mL, respectively. The MBC of GA when applied against methicillin-resistant S. aureus strains was >3200 μg/mL.37,40 The MBC of fraction of free phenolic acids against E. coli, P. aeruginosa, and S. aureus was 4860 μg/mL, 3780 μg/mL, and 5400 μg/mL respectively. 78 This study shows that FA, a hydroxycinnamic acid, was more active than GA, a hydroxybenzoic acid, on both growth inhibition and bactericidal activity against the bacteria tested. This result is in accordance with the findings of other authors.36,39,48,65 These authors demonstrated, with alkyl gallates and phenolic acids alkyl esters, that their antimicrobial efficacy was dependent on the length of the hydrophobic chain. The antimicrobial activity increased with the higher numbers of carbons in the chain,36,39,48 reaching a maximum at a certain carbon number, at which a cut-off phenomenon appears.36,39 This importance of the chemical structure and composition of the phenolic products on their biological function is emphasized by Wang et al. 91
Antimicrobial activities of phenolics products can involve various modes of action, namely destabilization and permeabilization of cytoplasmatic membrane and enzyme inhibition by the oxidized products, possibly through reaction with sulfhydyl groups or through more nonspecific interactions with the proteins, for example, formation of reactive quinones that can react with amino acids and proteins.7,15,46,50,57 Phenols can also inhibit the synthesis of nucleic acids of both Gram-negative and Gram-positive bacteria. 17
Regarding the effect of phenolic acids on physicochemical surface properties of bacterial cells it was verified that these compounds, in particular GA, changed bacterial hydrophobicity. Phenolic acids changed the polar, apolar, and electron acceptor (γ+) components of the bacterial cells. The electron acceptor ability, after exposition to GA and FA, increased for Gram-positive bacteria and decreased for those Gram negative. This result demonstrates that GA and FA are electrophilic products and seem to significantly interact with the bacterial surface components. Hydroxycinnamic acids are, generally, antibacterial and are less polar than the corresponding hydroxybenzoic acids, due to their propenoid side chain, and this property might facilitate the transport of these molecules through the cell membrane.12,51 In the present work, GA (hydroxybenzoic acid) caused more significant changes on the bacterial physicochemical properties than FA (hydroxycinnamic acid). Nonetheless, the values of MBC and MIC of FA for the several bacteria were lower than those of GA.
The type of microorganisms and its cell membrane structure and composition assume an important role in the susceptibility to antimicrobials. 26 In Gram-negative bacteria, the passage through the outer membrane (OM) is dependent on the chemical nature of the antimicrobial product and is regulated by the presence of hydrophilic channels, known as porins. These channels normally exclude the entry of hydrophobic substances. However, it is possible to weaken the OM by molecules that disintegrate the lipopolysaccharides (LPS) layer, generically named as permeabilizers.51,57,83 Some phenolic products have been reported as OM disintegrators, releasing LPS and increasing the permeability of cytoplasmatic adenosine triphosphate. 51 Further, certain phenolic products such as carvacrol and thymol, with hydrophobic character, have been reported to be active on the OM and had bactericidal activity.27,57
Bacterial cells have normally negative surface charge, under physiological conditions, due to the presence of anionic groups (e.g., carboxyl and phosphate) in their membranes.24,43,54 Nevertheless, the magnitude of the charge varies from species to species and is possibly influenced by various conditions, namely age of the culture, ionic strength, and pH.2,54 Zeta potential measurements demonstrated that after phenolic acids exposure, the cells become less negatively charged. This surface charge change was particularly verified for the Gram-negative bacteria and is the probable reason to explain their higher susceptibility when compared with the Gram-positive bacteria. The antimicrobial activity of phenolic acids is dependent on the concentration of the undissociated acid. Due to their partially lipophilic character, it is assumed that phenolic acids cross the cell membrane by passive diffusion in their undissociated form, disturbing the cell membrane structure and possibly acidifying the cytoplasm and causing protein denaturation. 11 The disruption of the cell membrane by localized hyperacidification is a potential mechanism to explain the antimicrobial action of phenolic acids against microorganisms. Campos et al. 11 reported that phenolic acids caused K+ efflux on two Gram-positive bacteria and this was related with intracellular acidification. This hyperacidification effect can alter cell membrane potential, making it more permeable. 11 This mechanism would explain the bactericidal effect of phenolic acids observed in this work because the intracellular acidification changes membrane permeabilization and causes irreversible changes, leading to cell death.
Cytoplasmic membrane permeabilization was observed in the uptake of PI, a nucleic acid stain to which cell membrane is usually impermeable. The results obtained demonstrate that phenolic acids compromise the integrity of the cytoplasmatic membrane. The percentage of cells with damaged membrane after exposure to GA was higher than FA for L. monocytogenes at all concentrations tested, and for other bacteria only at 100 μg/mL. But the percentage of cells of Gram-negative bacteria and S. aureus stained with PI after treatment with FA was higher than GA at 500 μg/mL and 1000 μg/mL. In addition, the percentage of cells with damaged membranes can be correlated with phenolic acids concentration. P. aeruginosa was the most susceptible bacterium. Up to now, no clear evidences existed concerning the ability of phenolic products to cause local rupture or pore formation in cell membranes. Typically, the cytoplasmatic membrane of bacteria acts as a barrier between cytoplasm and extracellular medium. The internal ionic environment of prokaryotic and eukaryotic cells, is generally rich in potassium, therefore, leakage of this ion has been used to monitor the membranolytic events in bacteria. On the other hand, K+ leakage is usually the primary indicator of membrane damage in microorganisms. 42 According to Carson et al. 13 marked leakage of cytoplasmic material is considered indicative of gross and irreversible cytoplasmic membrane damage. Some phenolic products are recognized to have membrane active properties against microorganisms, causing leakage of cell constituents. 32 These products could diffuse through the cytoplasmic membrane, increasing its permeability and, consequently, leakage of bacterial cell constituents including proteins, nucleic acids, and inorganic ions such as potassium or phosphate. 11 Previous studies performed with other phenolic acids (p-coumaric, caffeic, p-hydroxibenzoic, protocatechuic, vanillic, and syringic acids, thymol, eugenol, and carvacrol) support the main action of FA and GA at the cell membrane.11,57,82,89 Additionally, it was found by Ota et al. 53 that FA, p-coumaric acid and caffeic acid, affects the cell membrane structure by rigidity and alteration of the dynamics of phospholipid chains. Moreover, Tamba et al. 80 verified that (−)-epigallocatechin gallate induced large pore formation in lipid membranes resulting in leakage of the fluorescent probe calcein. The concomitant phenomena after the pore formation were the decrease in the diameter of vesicles and their transformation into small lumps due to the attractive interaction between neighboring lipid membranes. These authors also concluded that the binding of this flavonoid to the external monolayer of the lipid membranes increases its membrane area decreasing the intermembrane distance that induces an increase in its surface pressure. Other authors also referred the strong interaction between polyphenols and lipid membranes.77,93 These phenomena explain the antimicrobial activity of this compound. Kajiya et al. 33 in a study performed with (+)-catechin derivatives demonstrate that the activity of these phytochemicals depends on the number of hydroxyl groups on the B-ring. On the other hand, the affinity for lipid bilayers was augmented with elongation of the alkyl chain lengths of the derivatives. Similar to the referred studies we can hypothesize that the antibacterial activity of phenolic acids is associated with both the affinity for the lipid bilayer and the disruption of the membrane structure. Also, the phenolic acid–lipid interaction can help to explain the higher susceptibility of the Gram-negative bacteria. In fact, the lipid content of the cell walls of the Gram-negative bacteria is substantially higher than that of Gram-positive cell wall. 63 This result together with results from PI uptake, zeta potential, and contact angles assessment demonstrate that FA and GA interacted with the surface of Gram-negative and Gram-positive bacteria, promoting membrane damage, release of intracellular content, and the consequent cell death. This effect was dependent on the bacterial species. Significant effects were found particularly for E. coli and P. aeruginosa, the Gram-negative bacteria. These data corroborate the results obtained in a study performed by Lacombe et al., 41 where transmission electron microscopy micrographs demonstrated disintegration of the OM of cells treated with low concentrations of phenolic acids. Nevertheless, no direct correlation was observed between the structure of both phenolic acids and the alteration of membrane properties (physicochemical properties, surface charge, leakage of potassium, and PI uptake).
Even if FA and GA have antimicrobial effects, the effective concentrations are too high to apply these products per se for antimicrobial therapy. An important potential strategy to help combat the resistance problem involves the discovery and development of new active products capable of partly or completely suppressing bacterial resistance mechanisms. The development of what have been termed resistance-modifying agents (RMAs) represents an attractive strategy to mitigate the spread of bacterial drug resistance since it could facilitate the recycling of well-established antibiotics that are often cheaper and less toxic than new candidate antimicrobials. Phytochemicals can be promising RMAs in antimicrobial therapy, as recently reported by Abreu et al. 1 Moreover, some of these products may provide important structural scaffolds for the development of new antimicrobials. 22 The use of combinatorial and parallel synthesis chemistry techniques, high-throughput screening, and computational design will speed up the discovery of new and effective products to treat resistant infections. Immediate practical applications of FA and GA can be proposed for antiseptic and disinfectant formulations due to their recognized low cutaneous toxicity.45,55
Conclusions
Plant-derived molecules may offer a groundbreaking green approach to the discovery of broad-spectrum antimicrobials. In this study, it was found that FA and GA had antimicrobial activity against four pathogenic bacteria. These molecules led to a significant change in cell surface hydrophobicity, charge, induced PI uptake, and K+ leakage. To our knowledge this is the first study where the action of FA and GA on the surface properties, particularly the occurrence of local rupture or pore formation in the cell membranes, of Gram-negative and positive-bacteria is demonstrated.
Footnotes
Acknowledgments
This work was supported by Operational Programme for Competitiveness Factors–COMPETE and by FCT–Portuguese Foundation for Science and Technology through Project Phytodisinfectants—PTDC/DTP-SAP/1078/2012, and the PhD grants awarded to Anabela Borges (SFRH/BD/63398/2009) and Carla Ferreira (SFRH/BD/64927/2009). Also, this work was undertaken as part of the European Research Project SusClean (Contract n° FP7-KBBE-2011-5, project number: 287514). The authors are solely responsible for this work. It does not represent the opinion of the Community, and the Community is not responsible for any use that might be made of data appearing herein.
Disclosure Statement
No competing financial interests exist.