
Abstract
Resistance to ciprofloxacin in Escherichia coli is increasing parallel to increased use of fluoroquinolones both in The Netherlands and in other European countries. The objective was to investigate the contribution of active efflux and expression of outer membrane proteins (OMPs) in a collection of clinical E. coli isolates collected at a clinical microbiology department in a Dutch hospital. Forty-seven E. coli isolates a wide range of ciprofloxacin minimum inhibitory concentrations and known mutations in the quinolone resistance determining region were included. A fluorometric determination of bisbenzimide efflux was used two different efflux pump inhibitors and compared to quantitative reverse transcription–polymerase chain reaction (qRT-PCR) for the expression levels of acrA, acrB, tolC, yhiV, and mdfA efflux pump genes and the OMPs ompF and ompX. Six isolates (12.7%) showed increased efflux. Although in 35 isolates (76%), overexpression of ≥1 efflux pump genes using qRT-PCR was present. Only the combined overexpression of acrAB-TolC and mdfA correlated with the phenotypic efflux assay using glucose/carbonyl cyanide m-chlorophenylhydrazone with glucose. Thus, efflux was involved in ciprofloxacin resistance in a limited number of E. coli isolates collected at a clinical microbiology department in a Dutch hospital complementing other resistance mechanisms.
Introduction
In E. coli fluoroquinolone resistance usually occurs due to stepwise mutations in the quinolone resistance determining regions (QRDRs) of the DNA gyrase genes (gyrA and gyrB) and/or topoisomerase IV genes (parC and parE) and the presence of plasmid-mediated quinolone resistance (PMQR) determinants. While mutations in the QRDRs are required to achieve a clinical level of resistance, decreased expression of the outer membrane protein (OMP) OmpF or increased expression of OmpX, as well as increased efflux pump activity have been shown to contribute to fluoroquinolone resistance. There are essentially five families of chromosomally encoded bacterial efflux pumps: the resistance-nodulation-division (RND), major facilitator superfamily (MFS), multidrug and toxic compound extrusion (MATE) and small multidrug resistance (SMR) family. The RND efflux pumps, such as AcrAB and YhiV, and MFS pumps, such as MdfA are TolC-associated efflux pumps. These efflux systems are well characterized and play a major role in E. coli. 22 The overexpression of these efflux pump genes is related to antibiotic resistance in clinical isolates.26,34
A variety of phenotypic methods have been used to identify active efflux systems in E. coli.2,4,5,8,15,16,20,29,30 Most of these studies have utilized ethidiumbromide as a substrate. The bisbenzimide dye Hoechst 33342 is also widely used to characterize efflux pump activity in different species, such as Lactococcus lactis, and E. coli.11,27 This fluorescent marker has several advantages over ethidiumbromide, such as a high quantum yield, and it is less toxic.
A recently described 96-wells plate fluorescence assay allows for a rapid, high throughput evaluation of efflux activity in clinical isolates. Bisbenzimide accumulation and efflux can be studied using bisbenzimide as a fluorescent dye and carbonyl cyanide m-chlorophenylhydrazone (CCCP) and Phe-Arg-β-naphtylamide (PAβN) as efflux pump inhibitors (EPIs). 32 The difference in fluorescence between the steady states of bisbenzimide accumulation in the absence and presence of an EPI reveals the contribution of active efflux in a single isolate. However, there is no uniformity in the interpretation of the results, which makes it difficult to compare the contribution of active efflux between isolates from different studies. Another limitation of phenotypic efflux assays is that they do not provide information about the expression levels of the genes involved in efflux. Most studies that have investigated the contribution of active efflux in E. coli isolates use either phenotypic efflux assays3,5,8,13,20 or quantitative reverse transcription-polymerase chain reaction (qRT-PCR).17,26,31,33,34 A combined phenotypic and genotypic analysis of efflux pump activity in E. coli has been described in a limited number of studies.4,7,12 The purpose of the present study was to investigate the contribution of increased efflux activity and decreased influx to fluoroquinolone resistance in clinical E. coli isolates. A fluorometric determination of bisbenzimide efflux, with CCCP and PAβN as EPIs, was used and compared to qRT-PCR for the expression levels of acrA, acrB, tolC, yhiV, and mdfA efflux pump genes and the OMPs ompF and ompX.
Materials and Methods
Bacterial isolates
Forty-seven clinical E. coli isolates from a previous study were obtained from different inpatients and outpatients from a University Hospital in Leiden, The Netherlands, between January and December 2008. Nonrepetitive isolates with resistance to ciprofloxacin minimum inhibitory concentration (MIC) of ≥0.125 mg/l were included. All isolates showed ≥1 mutation in the QRDRs of gyrase A, parC, and/or parE. The plasmid-mediated quinolone resistance gene aac(6′)-Ib-cr was present in 17 of the isolates. Isolate no. 2404 contained a qnrA1 gene. E. coli ATCC 25922 and four fully susceptible E. coli isolates were included as negative controls.
Susceptibility testing
MICs of ciprofloxacin were determined using the agar dilution method according to the Clinical and Laboratory Standards Institute (CLSI) guideline. MICs were interpreted using EUCAST criteria (www.eucast.org/clinical_breakpoints/). E. coli isolates resistant to at least three different drug classes were defined as multidrug resistant (MDR).
Bisbenzimide accumulation and efflux assays
To determine active efflux, a semi-automated fluorometric method with bisbenzimide as a fluorescence marker was used as previously described. Briefly, cultures were grown in Mueller-Hinton broth to an OD600 of 0.6. Bacterial cells were collected by centrifugation at 13,000 rpm and resuspended in phosphate-buffered saline (PBS; 1 ml). The OD of all suspensions was adjusted to 0.3. Aliquots (0.18 ml) were pipetted in a black 96-well microtiter plate (flat-bottomed; Greiner Bio-One, Frickenhausen, Germany). The bacterial cells were added in the format in duplicate: columns 1–10 isolates with PBS; column 11–12 heat-inactivated cells (10 min 90°C). After adding 45 μl bisbenzimide (25 μM), fluorescence was measured in a microtiter plate reader (Mithras LB 940; Berthold Technologies, Bad Wilbad, Germany). The fluorescence was acquired in cycles during in cycles of 60 sec at 37°C. Fluorescence during influx was measured using an excitation filter of 350 nm and an emission filter of 460 nm over a 13-min period. The relative fluorescence at each time point was calculated in percentages, using the fluorescence values of the heat-inactivated cells divided by the fluorescence signals obtained in the test isolate. These data were plotted on a graph.
The microtiter plate reader was stopped after 13 min to add: (a) PBS (blank), (b) PBS containing glucose as an energy source (final concentration of 0.4%) to measure efflux activity, (c) PBS containing glucose (0.4%) with 172.4 μM PAβN or (d) 38.8 μM CCCP to inhibit efflux. All tests were performed in duplicate. Fluorescence during efflux was measured using an excitation filter of 350 nm and an emission filter of 460 nm. Intrinsic efflux activity was determined using the reference strain E. coli ATCC 25922 and four fully susceptible E. coli isolates. Efflux activity was expressed as the area between curves (ABC), which was calculated by subtracting the relative fluorescence in the wells containing CCCP/PAβN from that in the wells with glucose at each time point. For these five isolates, the mean and standard deviation (SD) of intrinsic efflux activity was calculated. Efflux was considered to be increased at 1.5×the SD above the mean of intrinsic efflux activity in the susceptible isolates.
RNA extraction and quantitative reverse transcription-PCR
Several colonies from overnight cultures were resuspended in PBS to make suspension of 0.5 McFarland. The suspension was diluted 1:1,000 in trypticase soy broth (Beckton Dickinson B.V., Breda, The Netherlands) and incubated at 37°C with shaking (150 rpm). The cultures were grown to a mid-log growth phase, which corresponded to an OD600 of 0.3. Total RNA was extracted using RNeasy kit (Qiagen Benelux B.V., Venlo, The Nederlands) according to the manufacturer's instructions. Genomic DNA was removed by DNase I kit (Invitrogen, Breda, The Netherlands). The concentrations and purity of the resulting RNA samples were measured at 260 nm using a Nanodrop ND-1000 spectrophotometer (Thermo Fisher Scientific, Wilmington, MA).
Relative quantification of gene expression was performed by using the OneStep RT-PCR Kit (Qiagen Benelux B.V.) with SYBR green (stock 10.000×; Sigma-Alderich®, Zwijndrecht, The Netherlands) in a CFX96™ Real-Time PCR Detection System (Bio-Rad Laboratories B.V., Veenendaal, The Netherlands). The genes included in the analysis were acrA, acrB, tolC, mdfA, yhiV, ompF, and ompX; glyceraldehyde 3-phosphate dehydrogenase (gapA) was used as a reference marker.
For primer design, an alignment of gene sequences in GenBank® was made using the AlignX program (Vector NTI Advance 11; Invitrogen). All primer sequences and annealing temperatures are shown in Table 1. All primers were synthesized by Biolegio B.V. (Nijmegen, The Netherlands). The RT-PCR mixture (50 μl) contained 1.2 μl of both forward and reverse gene-specific primers, 2 μl of dNTPs (10 mM), 10 μl of 5×Qiagen OS RT-PCR buffer, 1.5 μl of 2.5×SYBR green, 1 μg of template RNA, 2 μl of Qiagen OS RT-PCR enzyme mix. Each sample was placed on a 96-well plate (Bio-Rad Laboratories B.V.) and subjected to one-step reverse transcription at 50°C for 30 min for cDNA synthesis, 35 cycles of denaturation at 95°C for 15 sec, annealing at 60°C for 30 sec and extension at 72°C for 30 sec. PCR cycling was followed by melting curve analysis of 55°C–95°C (temperature transition rate of 0.5°C/sec). To assess relative gene expression levels, cycle threshold (CT) values normalized against the housekeeping gene (gapA) were calculated and compared with those for E. coli ATCC 25922 using the 2−ΔΔCT formula. Overexpression was defined as ≥1.5-fold increase in the expression of the gene.
All primers were designed in this study.
F, forward; R, reverse.
Statistical analysis
Data analysis was performed using SPSS (version 20.0; IBM Corp., New York, NY). Student t test was performed at a 5% level of significance (one-tailed, unpaired). The strength of a correlation between two variables (effect size) was calculated using the Kendall's tau b. This is a statistic method to measure the association between measured quantities.
Results
Antimicrobial susceptibility testing
The MICs of ciprofloxacin of the 47 clinical E. coli isolates varied from 0.5 to 1,024 mg/l. Forty-two isolates (89%) were classified as MDR (Table 2).
The numbers in bold indicate that the area between curves was 1.5 × SD above the mean for increased phenotypic efflux activity.
CIP mg/l, MIC of ciprofloxacin as determined with agar dilution.
MDR, multidrug resistant:+, if the isolate was resistant to ≥3 different drug classes; −, if the isolate was resistant to <3 different drug classes.
The numbers indicate the number of mutations in the QRDRs of gyrA, parC, and parE as determined in a previous study.21
PMQR, plasmid-mediated quinolone resistance determinant;+, present; −, absent.
Area between the curves in phenotypic efflux activity in the presence of glucose subtracted from glucose with the addition of the EPIs CCCP or PAßN.
As determined by qRT-PCR;+, upregulated gene; −, no upregulation.
As determined by qRT-PCR;+, downregulated gene; −, no downregulation.
QRDR, quinolone resistance determining region; ND, not determined; qRT-PCR, quantitative reverse transcription–polymerase chain reaction; CCCP, carbonyl cyanide m-chlorophenylhydrazone; PAβN, Phe-Arg-β-naphtylamide; SD, standard deviation.
The MICs of bisbenzimide and the EPIs: PAβN and CCCP (Sigma-Aldrich) were determined for all isolates and the quality control strain E. coli ATCC 25922 using the agar dilution method in accordance with CLSI guidelines. Based on these results a concentration of 25 μM for bisbenzimide, 38.8 μM for CCCP and 172.4 μM for PAβN were chosen.
Efflux activity evaluation: accumulation and efflux of bisbenzimide
Table 2 shows the efflux activity, expressed as ABC, for all isolates tested, including the fluoroquinolone-susceptible isolates. The reference strain E. coli ATCC 25922 showed inhibition of efflux activity, which was affected by CCCP and less so by PAβN (Fig. 1). This was considered as the intrinsic efflux activity of E. coli ATCC 25922. The four fully susceptible E. coli isolates showed similar results.
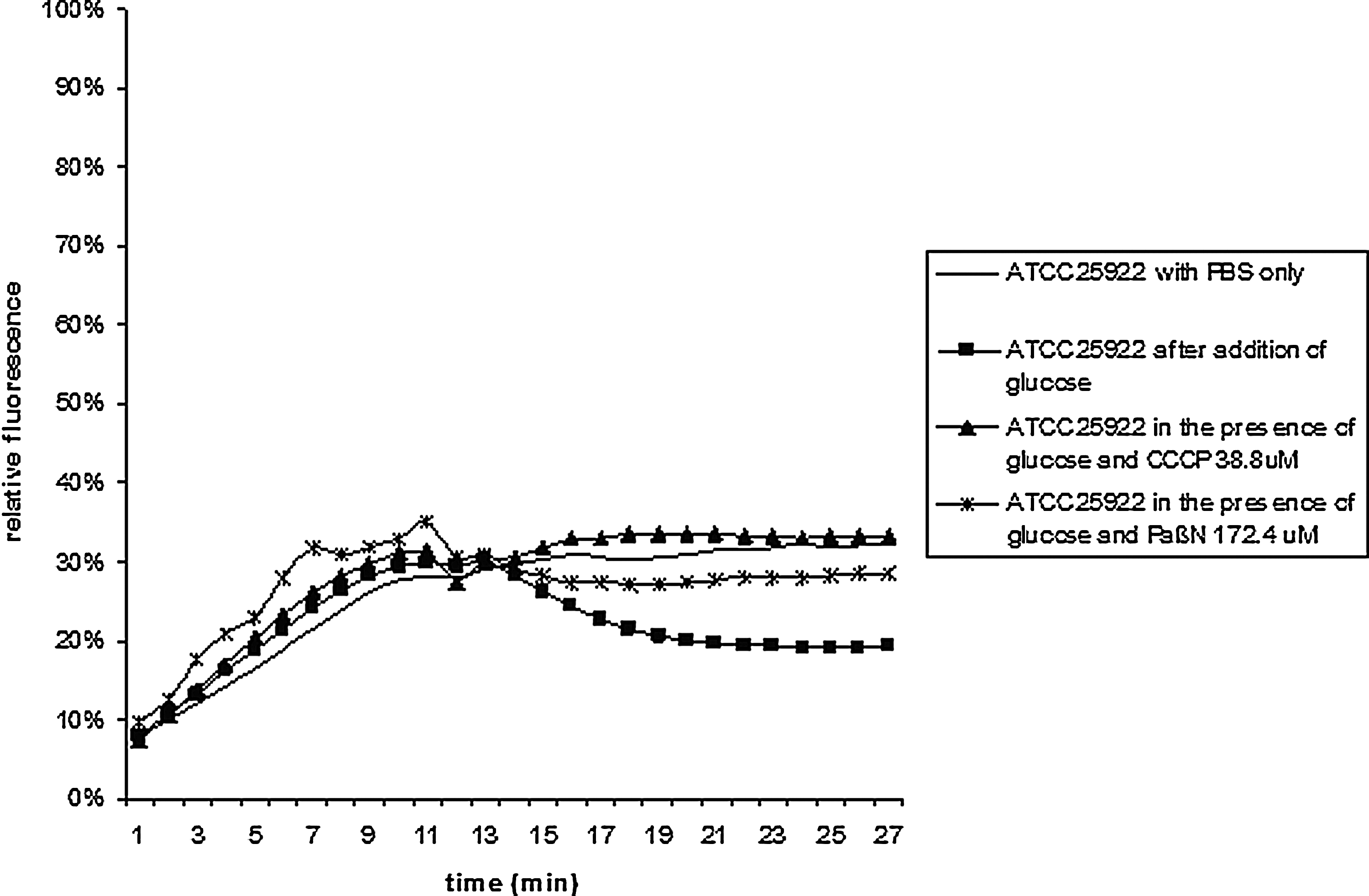
Accumulation (0–13 min) and efflux activity (13–30 min) of bisbenzimide by E. coli ATCC 25922 in the presence of glucose and in the presence of the optimum concentration of the EPIs CCCP and PAβN at 37°C. EPI, efflux pump inhibitor; CCCP, carbonyl cyanide m-chlorophenylhydrazone; PAβN, Phe-Arg-β-naphtylamide.
For the E. coli ATCC 25922 and the four susceptible E. coli isolates, the ABC between glucose/CCCP with glucose ranged from 2.02 to 2.51, with a mean of 2.33 (SD: 0.18). For PAβN, the ABC between glucose/PAβN with glucose with ranged from 0.72 to 1.66, with a mean proportion of 1.36 (SD: 0.38). Efflux was considered to be increased, if the ABC between glucose/CCCP with glucose was ≥2.60 or glucose/PAβN with glucose was ≥1.93.
One of the fluoroquinolone resistant isolates (no. 1629) showed inhibition of active efflux in the presence of both CCCP as with PAβN (Fig. 2). Active efflux in the presence of CCCP only was found in four isolates (9%). In the remaining 33 isolates no increased efflux was found with CCCP. One additional isolate (2525) showed increased efflux with PAβN only. One isolate showed an ABC of 1.63 (no. 1965), which was higher than the intrinsic efflux activity of the reference strain ATCC 25922, but below the level of active efflux. The remaining isolates showed an absence of an inhibitory effect in the experiments with PAβN. There was no association between the increasing MICs values, target alterations or an MDR phenotype and the presence of increased efflux activity.
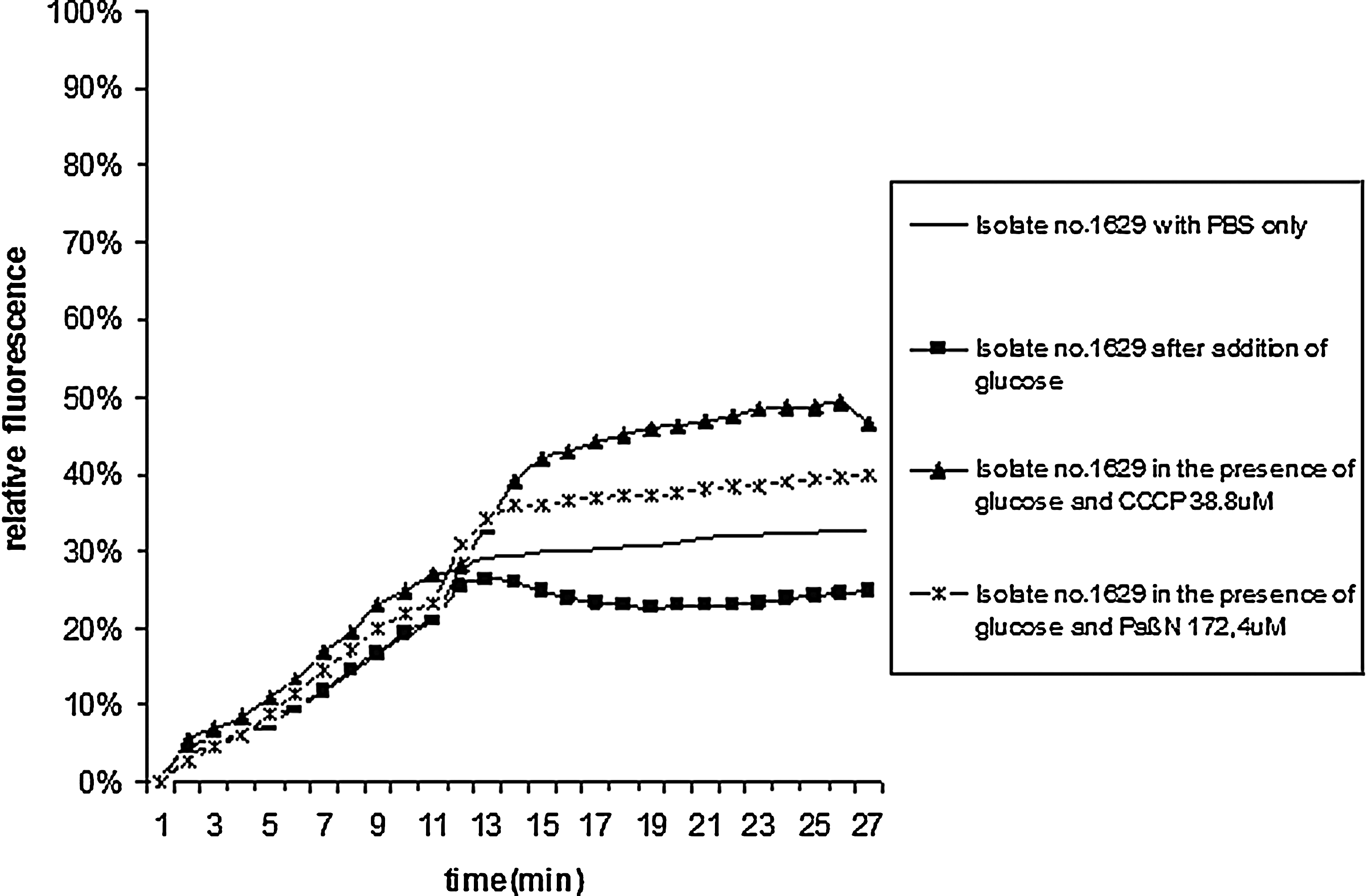
Accumulation (0–13 min) and efflux activity (13–30 min) of bisbenzimide by E. coli isolate 1629 in the presence of glucose and in the presence of the optimum concentration of the EPIs CCCP and PAβN at 37°C.
Analysis of quantification of RNA expression by qRT-PCR
Table 2 also shows the expression level of the efflux pump genes for all isolates tested, including the fluoroquinolone-susceptible isolates. Overexpression of ≥1 efflux pump genes was present in 35 isolates (76%). Eighteen isolates showed overexpression of acrAB-tolC, 33 isolates showed overexpression of mdfA and 17 isolates of yhiV. Nine isolates showed expression of three efflux pumps, acrAB-tolC, mdfA, and yhiV. Eleven isolates showed no overexpression of any of the efflux pump genes tested. No clear correlation was found between efflux and MICs of ciprofloxacin. Combining (a) the number of QRDR mutations and (b) the presence of PMQR determinants with genotypic efflux results did not reveal any correlation. Decreased expression of ompF was found in 33 isolates, respectively. Twenty-two isolates showed an upregulation of ompX.
Correlation between phenotypic efflux activity and overexpression of efflux pump genes
Nine isolates showed overexpression of all 3 efflux pump genes investigated (group 1), 8 isolates showed acrAB-tolC and mdfA overexpression (group 2), 15 isolates showed either yhiV or mdfA overexpression (group 3) and 1 isolates showed no genotypic overexpression (group 4). A significant correlation between the ABC of glucose/CCCP with glucose and the overexpression of acrAB-tolC with the mdfA efflux pump gene only (group 2 isolates; p=0.018, Kendall's tau b=0.343). The ABC between glucose and PAβN with glucose showed no significant correlations with overexpression with any of the groups tested (p=0.198).
Discussion
In the present study, the contribution of active efflux and expression of OMPs to fluoroquinolone resistance in a collection of clinical E. coli isolates was investigated using a combined phenotypic and genotypic approach.
Efflux pumps are expressed at a basal level in susceptible isolates and can be up-or down regulated in resistant isolates. The current study compared efflux in clinical fluoroquinolone resistant isolates to that in the reference wild type E. coli ATCC 25922 strain with functional efflux genes. In other studies on phenotypic efflux laboratory generated mutant E. coli strains have been used to describe active efflux in clinical isolates.5,7,14 Based on phenotypic tests, six of 47 (12.7%) resistant isolates showed increased efflux activity. The occurrence of increased efflux seems to be variable in clinical E. coli isolates.4,7,9 Christiansen et al. 4 found an indication of efflux in 76.7% of 60 clinical E. coli isolates with an MIC for ciprofloxacin ranging from 0.06 to 512 mg/l using a modified broth microdilution assay with the use of PAβN and 1-(1-naphtylmethyl) piperazine (NMP) as EPIs. Karczmarczyk et al. 7 used ethidiumbromide accumulation experiments to determine the activity of the AcrAB-TolC efflux pump. They found increased phenotypic efflux in 8/11 ciprofloxacin resistant E. coli isolates from food-producing animals using PAβN. Amabile-Cuevas et al. 1 on the other hand using a broth microdilution assay with PAβN found increased efflux in 12% of their clinical isolates, a percentage similar to ours. However, comparison of efflux results in clinical isolates from different studies remains difficult as different phenotypic methods and/or selection criteria have been used to detect phenotypic efflux activity. In the absence of established criteria to define active efflux in clinical isolates, we used the calculation of the ABC.
The fact that most isolates in this collection already showed a high number of accumulated QRDR mutations could also account for the lack of active efflux. 21 A recent publication of Singh et al. 24 investigated the contributions and interplay of fluoroquinolone resistance mechanisms at various time points during resistance development in a E. coli laboratory standard strain (MG1655) and two isogenic derivatives (ΔacrAB and ΔacrR). Interestingly, they found that in the initial stages of emerging resistance, efflux pump overexpression occurred before gyrase mutations and that once mutations were found, the pump activity had returned to normal levels. The isolates in our collection had accumulated high numbers of QRDR mutations already and may have had their efflux activity already tuned down. However, the six isolates with active efflux were amongst those with a high number of QRDR mutations and high MICs.
Of more than 40 putative transporters in E. coli, efflux pumps of the RND family, which are inhibited by PAβN, affect fluoroquinolone MICs when expressed with their own promotors under laboratory growth conditions.25,33 In both canine/feline and human E. coli isolates PAβN has been shown to reverse or reduce the fluoroquinolone MICs.8,12,23 In contrast, our results show that PAβN did not have much effect on the extrusion of bisbenzimide, suggesting that active efflux by RND efflux pumps did not contribute to ciprofloxacin resistance in the majority of our clinical isolates.
Using the genotypic approach, relative quantification of efflux gene expression levels was normalized against a fully susceptible reference strain, E. coli ATCC 25922, as is customary.12,26,34 In the present collection, various combinations of gene expression levels were found. This included: (a) alterations in the OMPs and overexpression of efflux pump genes; (b) no OMP alterations but only overexpression of efflux genes; (c) only OMPs alterations without overexpression of efflux genes; (d) no OMP alterations or overexpression of efflux genes. The results of the OMP alterations showed that outer membrane influx resistance is also involved. Although a significant decrease of bisbenzimide influx was not found, the lack of correlation may be explained by the fact that bisbenzimide uses other routes to enter the bacterial cell.
Convergence between the phenotypic and genotypic method was observed in isolates with overexpression of acrAB-tolC and mdfA and the ABC of glucose/CCCP with glucose in the phenotypic efflux assay. In E. coli, the tripartite acrAB-tolC system is considered to be the most important efflux pump. 36 Swick et al. 26 investigated fluoroquinolone susceptible and MDR E. coli isolates and found that overexpression of acrAB correlated with an MDR phenotype. In laboratory-generated mutants, overexpression of the efflux pumps AcrAB or MdfA have been shown to cause three to sixfold increase in fluoroquinolone resistance. 33 Previous studies also reported that the simultaneous overexpression of AcrAB and MdfA results in synergistic increases in resistance to fluoroquinolones.10,33 Our results show that simultaneous overexpression of these efflux pumps can be detected using the phenotypic assay with glucose and CCCP as inhibitor, which can be used as a screening tool to detect active efflux. In the remaining isolates, no correlation was found between phenotypic detection of active efflux or MICs with mRNA gene expression levels of the efflux genes investigated. Quantification of gene expression levels does not necessarily provide information on the final expression of the gene product and its functionality. Although mRNA expression of multidrug efflux pumps correlated well with protein expression levels using laboratory strains of Pseudomonas aeruginosa, 35 clinical isolates also showed discrepancies between mRNA expression and the corresponding protein. Molecular mechanisms causing these discrepancies include the half-lives of specific mRNAs or specific protein 28 and the post-transcriptional control of the protein translation rate. 19
In conclusion, active efflux was not the predominant resistance mechanism in clinical resistant E. coli isolates from our hospital. Living organisms have the capacity to adapt to changing environments. Increased efflux is not the only mechanism involved in the physiological adaptation process. There is a well regulated and coordinated interplay of multiple mechanisms of resistance. The determination of active efflux under defined experimental conditions may contribute to a better understanding of the contribution of active efflux in clinical MDR E. coli isolates.
Footnotes
Acknowledgments
We thank Yi Li and Jouraima Ras for their excellent technical assistance and Dr. Wil Goessens and Dr. Miguel Viveiros for the helpful discussions.
Disclosure Statement
The authors have no competing interests to disclose.