
Abstract
This study aimed to identify and characterize class 1 and 2 integrons and plasmid-mediated quinolones resistance (PMQR) genes in a collection of 113 multidrug resistance (MDR) Escherichia coli isolated from farm and wild lagomorphs between 2006 and 2008 in Northern Italy. Strains were examined for antimicrobial susceptibility by agar disk diffusion method and E-test for colistin (COL); integrons and gene cassettes content by real-time polymerase chain reaction (PCR) and DNA sequencing; PMQR genes by PCR and DNA sequencing; clonal relatedness by multilocus sequence typing; and plasmids by PCR-based replicon typing. Class 1 integrons were detected in 69 isolates (47 farm rabbits, 14 wild rabbits, and 8 wild hares). No class 2 integrons were found. Five different gene cassettes arrays were identified (aadA1, dfrA1-aadA1, orf in682-dhfrA5, orf in682-dfrA5-orfD ins21, and dfrA17-aadA5). Fifteen percent (17/113) of isolates carried oqxAB, no other PMQR determinants. All but one oqxAB-positive E. coli strains were recovered from farm rabbits. Seven out of 17 strains were associated with the predominant ST238 and carried from three to six different plasmid types, such as IncF, IncHI1, IncI1, IncN, IncP, IncX1, IncY, and ColE. COL resistance was identified in 6/113 strains (5.3%). This study provides new insights on the resistance phenotypes and the prevalence and dissemination of oqxAB in E. coli from farm and wild lagomorphs, suggesting that these animals may be reservoir of these genetic determinants in Italy and thus a potential source of PMQR E. coli for humans. PMQR mediated by oqxAB has not been detected in farm and wild lagomorphs before.
Introduction
M
Global surveillance studies demonstrate that resistance rates to sulfonamides and aminoglycosides increased in the past several years in almost all bacterial species and are largely attributed to horizontal exchange of transferable plasmids carrying class 1 integrons, site-specific recombination systems capable of capturing and expressing gene cassettes encoding for resistance against antimicrobials and disinfectants.21,23 Resistance to quinolones is primarily attributed to chromosomal mutations in DNA gyrase and/or topoisomerase IV genes or mutations in the regulatory genes of the acrAB efflux pump.17,20 However, plasmid-mediated quinolones resistance (PMQR) genes have been shown to play a relevant role in several members of the Enterobacteriaceae family, including Escherichia coli. 18 Three different PMQR mechanisms have been described: (1) protection of DNA gyrase from quinolone inhibition by Qnr pentapeptide repeat family (PRP) proteins encoded by the qnr genes (i.e., qnrA, qnrS, qnrB, qnrC, qnrD, and qnrVC); (2) quinolone acetylation mediated by a variant aminoglycoside acetyltransferase encoded by the aac(6′)-Ib-cr gene; (3) and quinolone extrusion by multidrug transporters encoded by the qepA or oqxAB genes.18,20
Several monitoring programmes collecting bacteria in animals and determining their susceptibility against antimicrobials used both in veterinary and human medicine have being carried out all over the world, but none of them has been targeted to lagomorphs, both those reared for meat production (i.e., farm rabbits) and those free-living (i.e., wild rabbits and hares). Therefore, knowledge on antimicrobial resistance of E. coli from these animal species is still poor. This study aimed to investigate the prevalence of class 1 and class 2 integrons and PMQR determinants in E. coli strains isolated from farm and wild lagomorphs in Northern Italy between 2006 and 2008.
Materials and Methods
Bacterial strains and antimicrobial susceptibility testing
E. coli strains were isolated from the intestinal content of 73 farm rabbits (O. cuniculus), 29 wild rabbits (O. cuniculus), and 11 wild hares (Lepus europaeus europaeus) from different regions of Northern Italy during the period 2006–2008. Farm rabbits samples were collected from animals showing a diarrheic syndrome suggestive of colibacillosis, whereas those from wild rabbits and hares were recovered from apparently clinically healthy animals hunted during the shooting season.
Samples were cultured onto Eosin-Methylene Blue agar (Oxoid) and incubated aerobically at 37°C for 18–24 h. Suspected E. coli colonies were identified by using the commercial kit RapID E20 (bioMérieux Vitek).
To verify pathogenic properties of isolates, the presence of the virulence gene eae was assessed by polymerase chain reaction (PCR) for the identification of the attaching and effacing Escherichia coli (AEEC), as previously described. 15 This gene encodes intimin, an indispensable protein for the formation of the A/E lesion.
Antimicrobial susceptibility of isolates was assessed by the agar disk diffusion method according to the Clinical and Laboratory Standards Institute. 4 The following antimicrobial disks and concentrations were used: chloramphenicol (CHL) 30 μg; nalidixic acid (NAL) 30 μg; streptomycin (STR) 25 μg; tetracycline (TET) 30 μg; and trimethoprim/sulfamethoxazole (SXT) 25 μg. Breakpoints were interpreted following the CLSI susceptibility criteria. 4 Evaluation of colistin (COL) susceptibility was assessed by using E-test (bioMérieux Italia Spa) with direct reading of the minimal inhibitory concentration according to the manufacturer's instructions (susceptibility ≤2 mg/L, resistance ≥2 mg/L). E. coli ATCC 25922 was used as quality control strain.
Detection and characterization of integrons and PMQR genes
All E. coli strains were screened for the presence of class 1 and class 2 integrons and PMQR determinants. Integrons were detected by real-time PCR assay using previously described primer sets. 7 Specific TaqMan and Molecular Beacon probes were designed for detection of the intI1 (5′-FAM-TGC CCG TTC CAT ACA GAA GC-3′-IBFQ) and intI2 genes (5′-FAM-CGC GAT CCA GCC TGA CCT CTT CAC TGC GAT CGC G-3′-IBFQ), respectively. Real-time PCR amplifications were performed in a LightCycler®480 Real-Time PCR System (Roche Diagnostics). PMQR genes, namely qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxA, and oqxB were searched using primers and conditions as previously described.11,26
Integron-borne gene cassettes and oqxAB amplicons were sequenced as previously described11,12,26 by the BigDye Terminator v3.1 chemistry in the ABI 3100 Genetic Analyzer (Applied Biosystems) according to the manufacturer's instructions. Comparative analysis of nucleotide sequences was performed using the BLASTN software (http://blast.ncbi.nlm.nih.gov/).
Multilocus sequence typing
Sequence types (ST) of all strains positive for class 1 integrons and/or oqxAB genes were determined by the EcMLST scheme. The allelic profiles and the STs were obtained submitting the allele sequences at the EcMLST Web site (www.shigatox.net/ecmlst/cgi-bin/index).
Plasmid characterization and location of integrons and resistance genes
Plasmids from oqxAB positive strains were typed by using the PCR-based replicon typing as previously described. 3 The location of integrons and oqxAB genes was defined by transformation assays. Plasmid DNA from four selected donor strains was purified by the PureLink® HiPure Plasmid Midiprep Kit (Invitrogen Life Technologies) and used to transform High efficiency DH5α/Top10 E. coli chemically competent cells (Invitrogen Life Technologies). Transformants were selected on Luria-Bertani agar plates containing 25 mg/L STR or 30 mg/L NAL. The presence of oqxA, oqxB, and integrons was detected by PCR as previously described.11,12
Results
A collection of 113 E. coli strains showing resistance to at least three different antimicrobial classes among fluoroquinolones, aminoglycosides, tetracyclines, sulfonamides, polymyxins, and phenicols was examined in this study. All strains isolated from rabbits showing a diarrheic syndrome suggestive of colibacillosis caused by AEEC were positive for the eae gene. On the other hand, all strains isolated from clinically healthy animals (i.e., wild rabbits and hares) were negative for this gene.
Figure 1 shows resistance profiles among E. coli isolates stratified by source (i.e., farm rabbits, wild rabbits, and hares) and tested against six antimicrobials of different classes (i.e., NAL, STR, TET, SXT, COL, and CHL). All E. coli strains were resistant to TET. Isolates exhibited high resistance against STR (93.81%), NAL (77.88%), and SXT (68.14%). Resistance to CHL was shown by 39.82% of isolates. Out of 113 E. coli strains, 46 (40.7%) showed MDR against 3 antimicrobial classes, 40 (35.40%) against 4 antimicrobial classes, 25 (22.12%) against 5 antimicrobial classes, and 2 isolates were resistant to all tested antimicrobials. Of note, 5.3% of the isolates showed resistance to COL (MIC range between 3 and 8 mg/L). MDR resistance patterns and the source and the year of isolation of E. coli strains are shown in Table 1. Similar AMR profiles were observed among E. coli isolated from farm rabbits, wild rabbits, and hares.
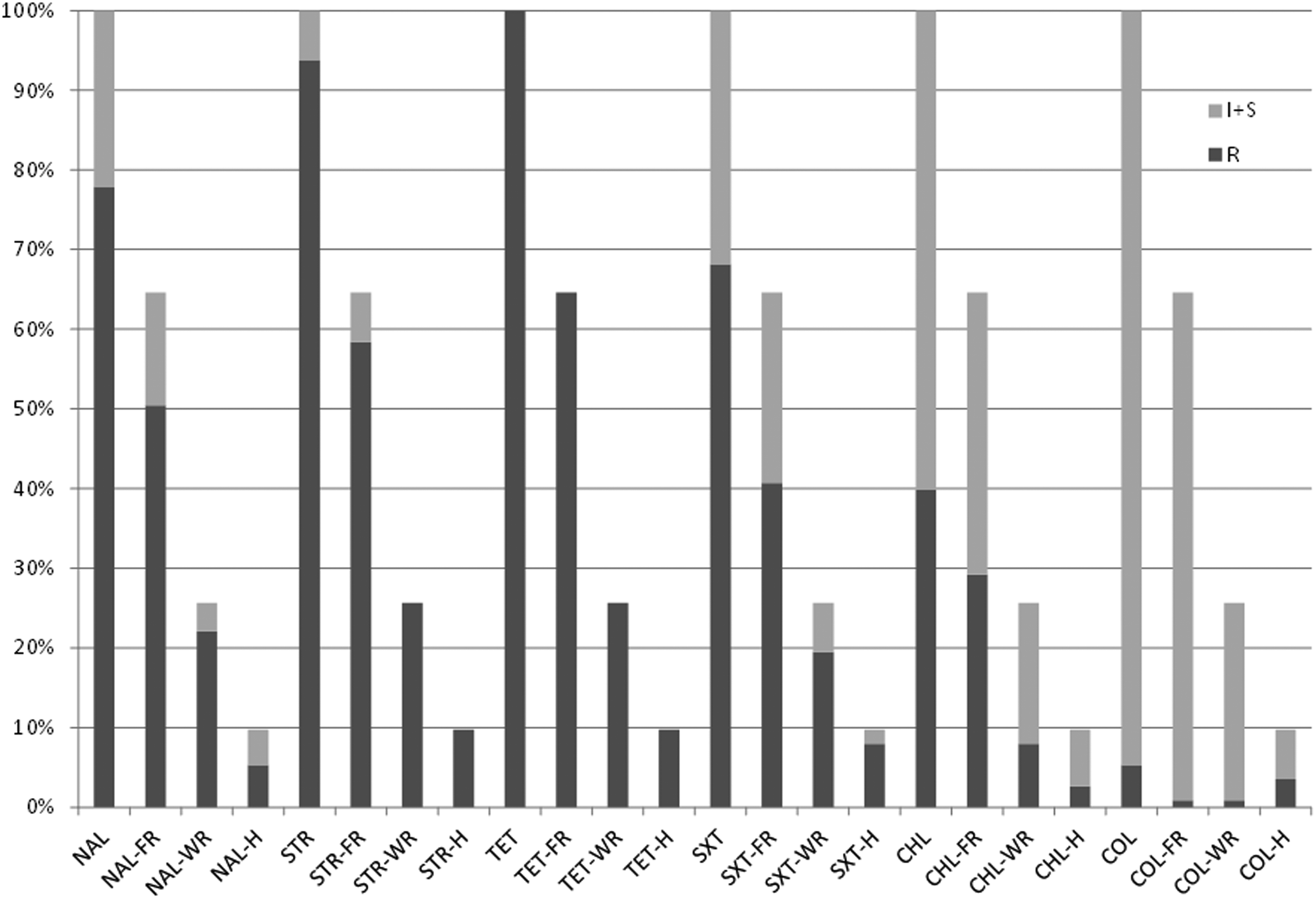
Phenotypic resistance profiles of Escherichia coli strains. E. coli isolates were stratified by source (FR, farm rabbits; WR, wild rabbits; H, hares) and tested against six antimicrobials of different classes (NAL, nalidixic acid; STR, streptomycin; TET, tetracycline; SXT, trimethoprim/sulfamethoxazole; CHL, chloramphenicol; COL, colistin). R, resistant; I, intermediate; S, susceptible.
MDR (i.e., resistance to ≥3 different antimicrobial classes).
MDR, multidrug resistance; NAL, nalidixic acid; STR, streptomycin; TET, tetracycline; SXT, trimethoprim/sulfamethoxazole; CHL, chloramphenicol; COL, colistin.
Class 1 integrons were detected in 69 isolates (61.06%): 47 strains from farm rabbits (68.12%), 14 from wild rabbits (20.29%), and 8 from wild hares (11.59%). No class 2 integrons were found. The most frequently identified gene cassette was aadA1, encoding aminoglycoside 3′-adenyltransferase conferring resistance to spectinomycin and STR, alone (29 strains, 42.03%) or in combination (32 strains, 46.38%) with dfrA1 variant, conferring resistance to trimethoprim. In three strains (4.35%) the dfrA17-aadA5 was also detected. Five strains showed dfrA5 in combination with orfIN682 (1.45%), four of them including also orfD (5.80%) (data not shown). The latter two cassettes encode for hypothetical proteins of unknown function.
Out of 113 E. coli strains, 17 (15.04%) were found positive both for oqxA and oqxB genes. Most of oqxAB-positive strains (16/17) were obtained from farm rabbits, except for one from a wild rabbit. No qnr (qnrA, qnrB, qnrC, qnrD, and qnrS) and qepA genes were detected. Table 2 summarizes characteristics of oqxAB-positive E. coli strains. All oqxAB-positive strains showed resistance to NAL, TET, and STR; 11/17 (64.7%) were resistant to SXT; 9/17 (52.9%) to CHL; and 1/17 (5.8%) to COL. All but 2 (88.2%) oqxAB-positive strains carried class 1 integrons (dfrA1-aadA1, dfrA17-aadA5, and aadA1) and 7 (41.18%) strains were associated with the predominant ST238. Plasmid content of the oqxAB-positive strains demonstrated that all strains carried a variety of plasmid types, such as IncF, IncHI1, IncI1, IncN, IncP, IncX1, IncY, and ColE. All strains carried an IncF plasmid characterized by the replicon FII (repF) in a large percentage, associated with additional replicons of the FIC or FIB type, as previously described for many IncF plasmids. 22 In all but 1 (94.12%) oqxAB-positive strains, the I1 plasmid was detected and shared by different phylogenetically unrelated STs. Transformation assays showed that integrons and oqxAB genes of the selected donor strains were located on plasmids belonging to IncI1 and IncF types, respectively.
Italian region.
ST, sequence type; CC, clonal complex; FR, farm rabbit; WR, wild rabbit; N, negative; N/A, not assigned; MLST, multilocus sequence typing; PBRT, PCR-based replicon typing.
Multilocus sequence typing (MLST) of 71 E. coli carrying integrons and/or PMQR genes showed high genetic heterogeneity among isolates. Figure 2 shows STs and clonal complexes (CCs) identified among the E. coli strains. Overall 30 different STs were recognized, with ST140 (8/71), ST238 (8/71), and ST119 (7/71) being the predominant types. Thirty-seven isolates (52.11%) belonged to novel STs that were submitted to the EcMLST Web site as ST1004 to ST1020. The 30 STs were grouped into 6 CCs, with CC17 and CC31 the most frequently detected (21.13% and 12.68%, respectively), although 41 strains (57.75%) belonged to CCs not defined in the EcMLST database.
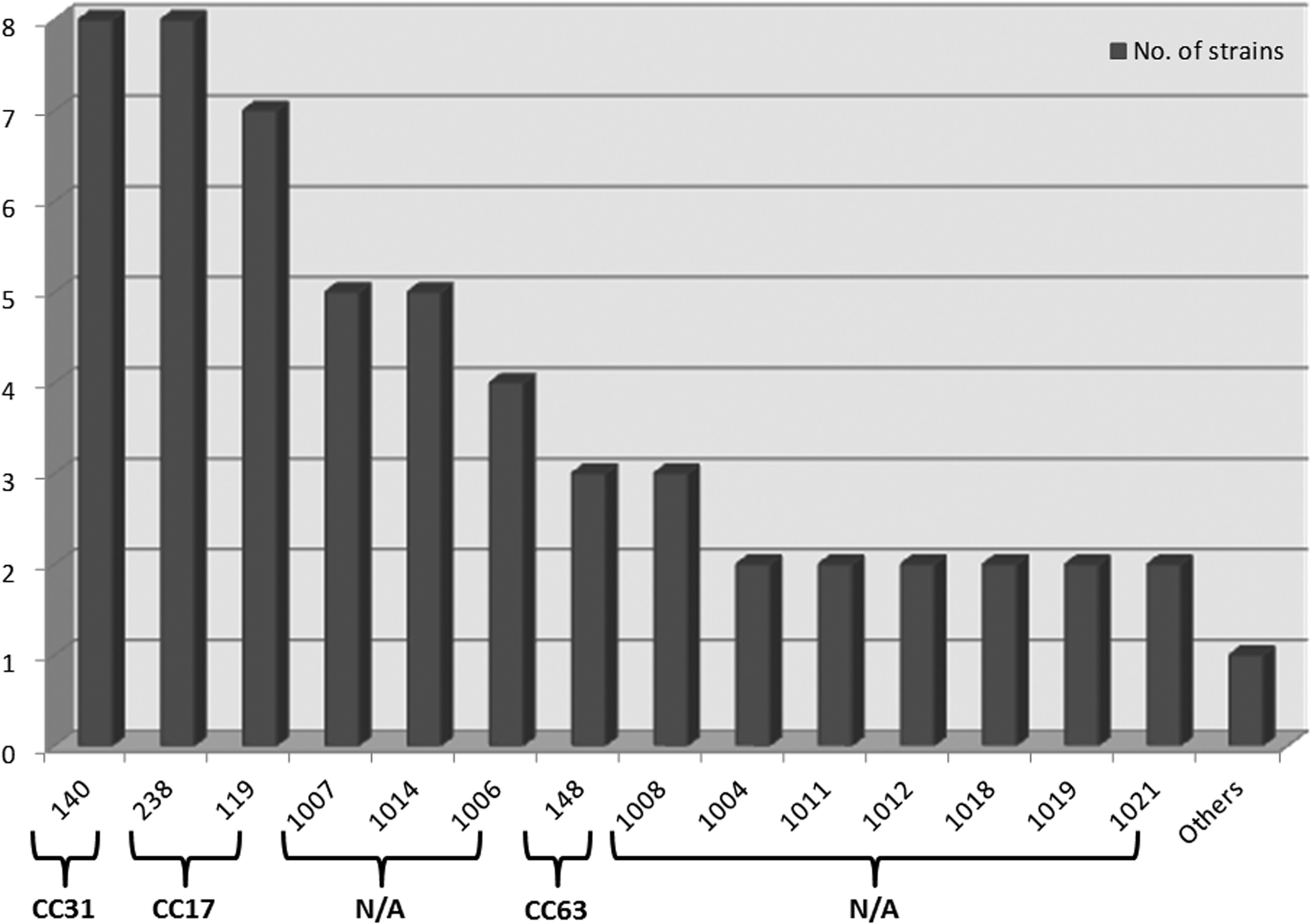
STs and CCs of integrons and PMQR-positive E. coli strains. N/A, not assigned; other STs comprised genotypes identified only once: ST134 (CC30); ST145 (CC31); ST171 (CC23); ST259 (CCN/A); ST317 (CCN/A); ST461 (CC41); ST855 (CCN/A); ST857 (CCN/A); ST1005 (CCN/A); ST1009 (CCN/A); ST1010 (CCN/A); ST1013 (CCN/A); ST1015 (CCN/A); ST1016 (CCN/A); ST1017 (CCN/A); ST1020 (CCN/A). STs, sequence types; CCs, clonal complexes; PMQR, plasmid-mediated quinolones resistance.
Discussion
Resistance rates in pathogenic and commensal E. coli of food-producing and wild animals are becoming an argument of scientific interest. In this study, we examined pathogenic and commensal E. coli strains recovered from intestinal contents of farmed and wild lagomorphs, respectively. Since coliform bacteria are rarely isolated from healthy lagomorphs due to the low number of bacterial cells in normal intestinal contents, 16 the recovery of E. coli was performed from farmed rabbits dead for colibacillosis. Otherwise, we should have euthanized a high number of healthy rabbits to obtain a sufficient number of commensal strains to be analyzed. For ethical reasons this option was discarded. On the other hand, diseased wild lagomorphs are extremely difficult to find in nature; therefore, wild rabbits and hares hunted during the game season were sampled. Commensal E. coli do not cause apparent disease but resistance genes identified on transmissible genetic elements allow their horizontal transfer among pathogenic E. coli and other bacterial species. 5 The results of this study showed that a significant number of pathogenic and commensal E. coli isolates recovered from domestic and wild lagomorphs possessed integrons. Gene cassettes encoding resistance to aminoglycosides and trimethoprim were prevalent. These results are in agreement with previous studies from other countries that frequently found class 1 integrons in bacteria from various sources,21,23,25 suggesting that integrons are playing an important role in the dissemination of trimethoprim and aminoglycoside resistance. The rapid development of fluoroquinolone resistance has been attributed to PMQR determinants since they can confer low-level resistance to quinolones, promoting the selection of high-level resistant strains with mutations on the chromosome in gyrA and parC genes. 17 Some surveys demonstrated a high prevalence of PMQR determinants from diseased food-producing animals in Guangdong, China. Among the E. coli isolates, oqxAB was the most prevalent PMQR gene.14,26 oqxAB is a gene encoding for a multidrug efflux pump, oqxAB, of the resistance-nodulation-cell division (RND) family. 10 First, described by Sorensen et al. 19 in E. coli strains isolated from swine manure, oqxAB can be carried on both plasmids and chromosomes and confers resistance to several antimicrobial agents, including fluoroquinolones, olaquindox, ampicillin, and CHL. 20 Currently, little information is available on the prevalence of oqxAB in E. coli recovered from animal sources and this study reports for the first time the presence of these genes in E. coli from domestic and wild lagomorphs scarcely investigated in global surveillance studies. We detected high prevalence of the oqxAB multidrug efflux pump genes. Generally, there was a slightly greater prevalence of the oqxAB genes among isolates recovered from farm rabbits than among isolates from wild lagomorphs. These genes were associated with the prevalent ST238, but also detected in other unrelated genotypes. No correlation between the STs and the animal geographic or temporal origin was found. In accordance with previous reports that integrons and PMQR genes are often located on plasmids,2,26 in this study transformants carried an IncF or IncI1-type plasmid harboring oqxAB genes or the class 1 integron, respectively. Industrial rabbit farming may be associated with a greater disease potential and therefore a greater tendency for antimicrobial use to control diseases. In particular, enteritis caused by E. coli represents the most common intestinal disease of rabbits. 13 Of note, COL that is frequently used in rabbit farming reached 5.3% of resistance in this collection. This is worrisome, since COL is currently playing an important role in human therapy for infections sustained by Gram-negatives showing resistance to the last generation antibiotics, such as carbapenems. The finding of emerging antimicrobial resistance mechanisms in wild rabbits and hares is of great value and can support further surveillance studies and epidemiological tracing of the potential animal-reservoir of relevant resistance determinants. To the best of our knowledge, this is the first article describing PMQR genes, integrons, and resistance plasmids in E. coli strains isolated from domestic and wild lagomorphs in Italy. The dissemination and the maintenance of these genetic determinants among both pathogenic and commensal bacteria may rebound on public health for the potential horizontal gene transfer between animal and human microorganisms.
Footnotes
Acknowledgments
This work was financed by grants from the Department of Comparative Biomedicine and Food Science, University of Padua and from the University of Padua (CPDA095771/09). We thank Dr. Giacomo Berto (University of Padua, Italy) for his laboratory work; Dr. Marianne Sunde (National Veterinary Institute, Norway), Dr. Stefan Schwarz (Friedrich-Loeffler Institut, Germany), Dr. Beatriz Guerra-Roman (Federal Institute for Risk Assessment, Germany), and Dr. Dik Mevius (Centraal Veterinair Instituut, The Netherlands) who kindly provided integron positive controls used in this study; and Dr. Hans Steinsland (Michigan State University, USA) for his contribution to the ![]() Web site.
Web site.
Disclosure Statement
The authors have no competing financial interests to declare.