
Abstract
Twelve nonduplicated KPC-2-producing enterobacterial isolates, including three Escherichia coli, two Citrobacter freundii, two Enterobacter cloacae, four Enterobacter aerogenes, and one Klebsiella oxytoca, were collected from various clinical samples within 18 months (March 2011 to September 2012). Two of the 12 patients died from infections caused by KPC-2-producing pathogens, while the rest of the patients with KPC-2-producing pathogens improved or were cured. The majority of the clinical isolates exhibited a high-level of resistance to oxyimino-cephalosporins and carbapenems, and possessed self-transferable blaKPC-2-carrying plasmids with sizes ranging from 20 to 120 kb. Most isolates carried blaCTX-M and plasmid-mediated quinolone resistance genes, while some isolates produced 16S rRNA methylases (ArmA or RmtB). The genetic environment of blaKPC-2 of most clinical strains was consistent with the genetic structure surrounding blaKPC-2 on the plasmid pKP048, which contains an integration structure of a Tn3-based transposon and partial Tn4401 segment. Inserted fragments (truncated blaTEM) were detected upstream of the blaKPC-2 gene for two E. aerogenes strains. In conclusion, the enterobacterial isolates exhibited sporadic emergence and did not arise by clonal spread at our hospital. The outcome of infections caused by KPC-producing enterobacterial isolates and their mortality were closely associated with the baseline condition of patients. The spread of blaKPC-2 gene between different enterobacterial species in China was mainly mediated by horizontal transfer of the Tn3-based transposons and not the blaKPC-2-carrying plasmids.
Introduction
C
Recently, KPC-producing Enterobacteriaceae, such as K. pneumoniae, Klebsiella oxytoca, Escherichia coli, Serratia marcescens, Citrobacter freundii, Proteus mirabilis, Enterobacter cloacae, and Enterobacter amnigenus, have been identified across China.3,4,9,10,13,14 The blaKPC genes among the KPC-producing Enterobacteriaceae isolated from China were primarily located on various plasmids and were mostly carried on a Tn3-based transposon composed of a partial Tn4401 structure and a Tn3-based element.4,9,10,14 From March 2011 to September 2012, twelve KPC-2-producing enterobacterial isolates, including three E. coli, two C. freundii, two E. cloacae, four Enterobacter aerogenes, and one K. oxytoca, were collected at our hospital. Phenotypic and genotypic features of the isolates were screened. The genetic environments of blaKPC-2 gene were also analyzed.
Materials and Methods
Bacterial strains
All clinical strains were isolated from clinical samples (Table 1) in a 4,000-bed tertiary care hospital. All clinical isolates were identified by VITEK 2 GN ID cards (bioMérieux, Inc., Hazelwood, MO) and VITEK® MS (bioMérieux SA, Marcy-l'Etoile, France). E. coli ATCC 25922 was used as the quality control strain for antimicrobial susceptibility testing. The sodium-azide-resistant E. coli J53 strain was used as the recipient for conjugation testing.
MICs of transconjugants.
CTX, cefotaxime; FEP, cefepime; TZP, piperacillin-tazobactam; IMP, imipenem; MEM, meropenem; ETP, ertapenem; AK, amikacin; LEV, levofloxacin; N/A, not applicable; N/D, not detected; PFGE, pulse-field gel electrophoresis; MICs, minimal inhibitory concentrations.
Antimicrobial susceptibility testing
The minimum inhibitory concentrations (MICs) of cefotaxime (CTX), cefepime (FEP), piperacillin-tazobactam (TZP), imipenem (IMP), meropenem (MEM), ertapenem (ETP), amikacin (AK), and levofloxacin (LEV) were measured by E-test method (AB bioMérieux, Solna, Sweden). All protocols were performed according to the manufacturer's instructions. The phenotypic presence of carbapenemase enzymes was detected by the modified Hodge test according to the guidelines of the Clinical Laboratory Standards Institute (CLSI). 1
Conjugation testing
Conjugation experiments were carried out in Luria–Bertani (LB) broth. Cultures of donor and recipient cells in logarithmic phase (0.5 mL of each) were added to 4 mL of fresh LB broth and incubated overnight without shaking. Transconjugants were selected on China blue agar plates containing MEM (1 μg/mL) and sodium azide (200 μg/mL; Sigma-Aldrich Co., St. Louis, MO).
Molecular detection of resistance genes
β-Lactamase genes, including blaKPC, blaTEM, blaSHV, blaOXA-1, blaCTX-M, blaPER, blaVEB, and blaGES; plasmid-mediated quinolone resistance (PMQR) genes, including qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr; and 16S rRNA methylase genes, including armA, rmtA, rmtB, rmtC, and rmtD, were screened as previously described. 15
Pulse-field gel electrophoresis analysis
Pulse-field gel electrophoresis (PFGE) typing was performed following the procedures developed by the U.S. Centers for Disease Control and Prevention PulseNet program. 8 DNA fingerprints were obtained from PFGE profiles of genomic DNA digested with XbaI (New England Biolabs, Beijing, China).
Plasmid analysis and Southern blot
A blaKPC probe was constructed by labeling a blaKPC PCR product using the PCR DIG Probe Synthesis Kit (Roche Applied Sciences, Mannheim, Germany). S1-PFGE and Southern blot were performed as previously described methods. 12
Genetic environment analysis of blaKPC gene
The genetic background of the blaKPC-2 gene was studied by the PCR mapping approach. Amplifications were performed using specific primers targeting the blaKPC-2 gene.5,9 All amplification products were sequenced by a DNA analyzer (3730xl; Applied Biosystems, Life Technologies, Foster City, CA).
Results
Emergence of KPC-2-producing enterobacterial isolates
In total, 12 nonduplicated KPC-2-producing enterobacterial isolates were collected from various clinical samples from March 2011 to September 2012. These included three E. coli, two C. freundii, two E. cloacae, four E. aerogenes, and one K. oxytoca. E. cloacae IR1503 and IR1504 belonged to the same clone based on their PFGE patterns. This was also observed in E. aerogenes IR1316 and IR1317. Two of the 12 patients died from infections caused by KPC-2-producing pathogens. Other patients with KPC-2-producing pathogens displayed signs of improvement or were cured (Table 1).
Conjugation of plasmids and antimicrobial susceptibilities
The blaKPC-2-carrying plasmids carried by most clinical enterobacterial isolates (except E. coli IR1405 and C. freundii IR1508) were transferred to E. coli recipient J53 by in vitro conjugation test. Most of the clinical isolates exhibited high-level resistance to oxyimino-cephalosporins (CTX and FEP) and carbapenems, including IMP, MEM, and ETP. E. aerogenes IR1507 and K. oxytoca IR1509 were not sensitive to these compounds (Table 1). The MICs of the transconjugants are also listed in Table 1.
Prevalence of resistant determinants
Table 2 lists the common resistance genes carried by KPC-2-producing Enterobacteriaceae. Most isolates carried the blaCTX-M and PMQR genes, while IR1405, IR1501, and IR1503 produced ArmA or RmtB that mediates the high-level resistance to AMK.
GenBank accession number: FJ628167.
Plasmid analysis and genetic environment of blaKPC-2 gene
S1-PFGE and Southern blot analysis showed that all clinical isolates carry several different-sized plasmids. The blaKPC-2 gene was located on various plasmids with sizes ranging from 20 kb to ∼100 kb (Fig. 1). Strains IR1401 (E. coli), IR1405 (E. coli), IR1502 (E. aerogenes), and IR1509 (K. oxytoca) contained blaKPC-2-carrying plasmids with similar sizes (about 40 kb) (Table 2). Fragments of partial structures around the blaKPC-2 gene were amplified by a PCR mapping approach. Most strains contained a genetic structure consistent with the environment surrounding the blaKPC-2 in the plasmid pKP048. 9 Inserted fragments were detected upstream of the blaKPC-2 gene for strains IR1502 (E. aerogenes) and IR1316 (E. aerogenes), which contain a 321 and 221 bp truncated blaTEM, respectively (Fig. 2). Interestingly, the genetic context of blaKPC-2 gene cannot be identified for isolates IR1405 (E. coli), IR1508 (C. freundii), and IR1509 (K. oxytoca) using previously described primer sets.5,9
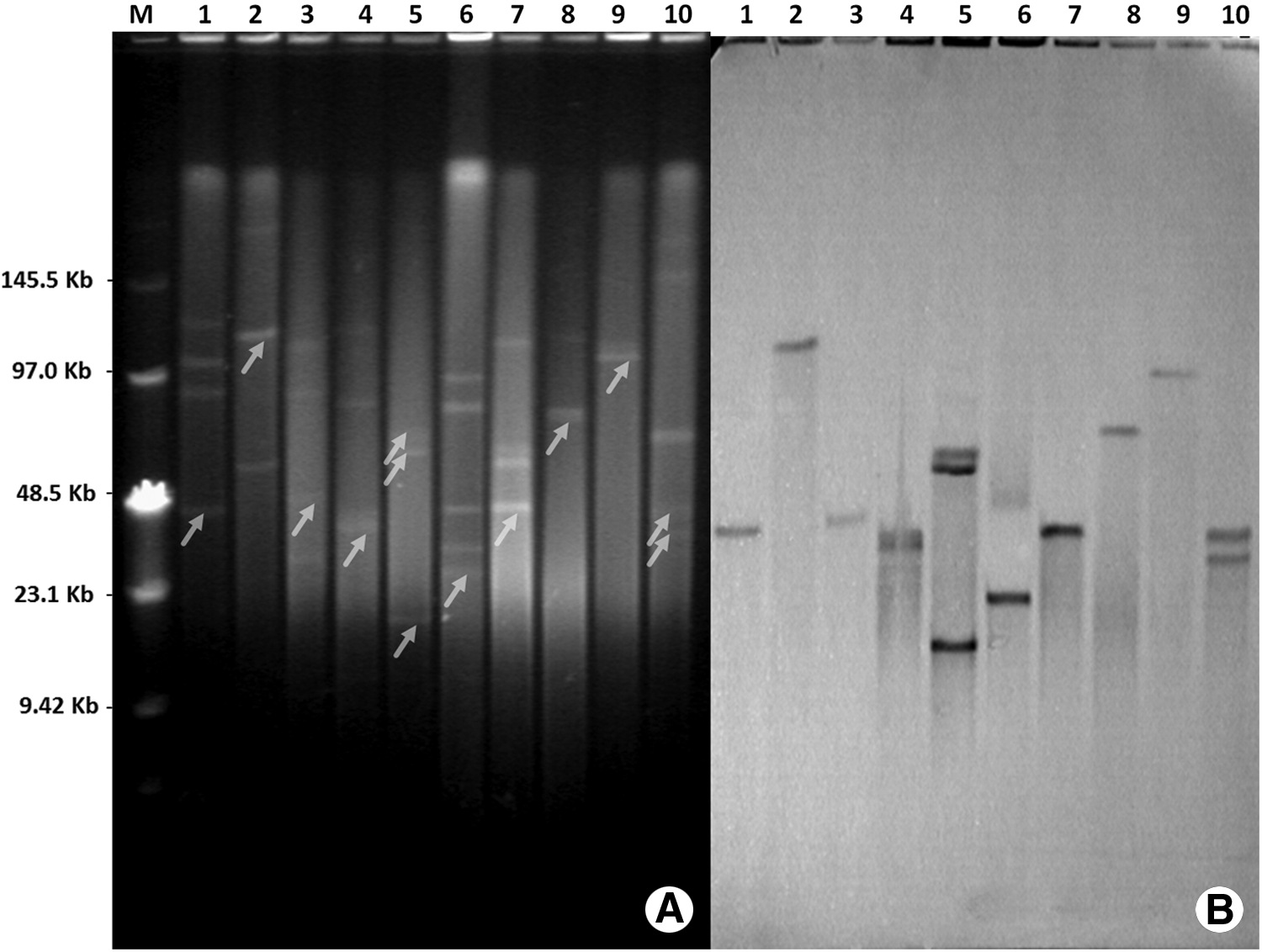
S1 nuclease digestion of DNA, PFGE, and hybridization results for blaKPC-carrying isolates.
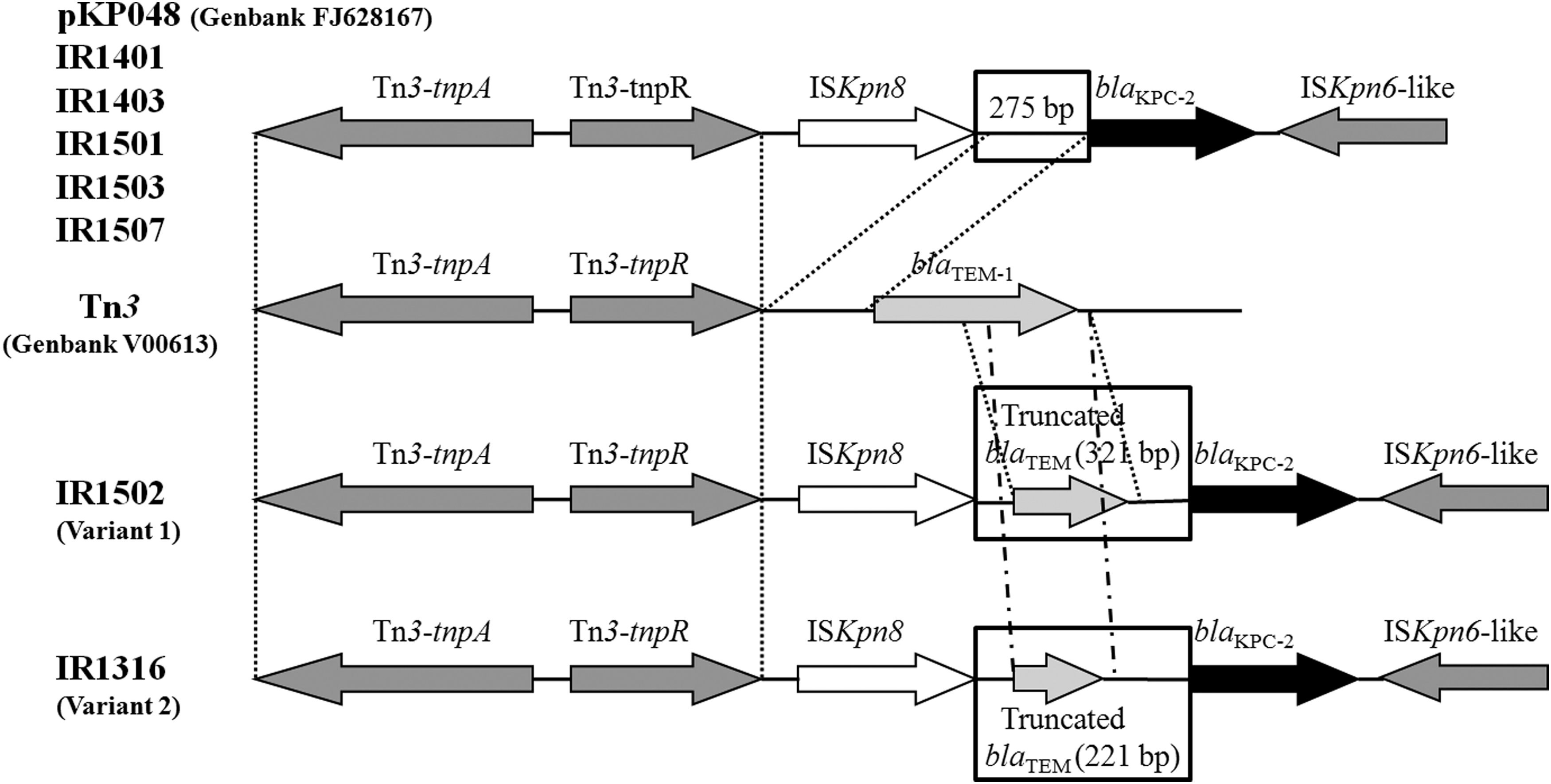
Schematic representation of the genetic structures surrounding the blaKPC-2 gene. Various genes and their directions of transcription are presented as broad arrows. The area delimited by two dotted lines indicates identical regions.
Discussion
In this study, 12 nonduplicated KPC-2-producing enterobacterial isolates were collected from March 2011 to September 2012. Two E. cloacae strains (IR1503 and IR1504) and two E. aerogenes strains (IR1316 and IR1317) were cultured from different specimens of two patients, respectively, while the other eight strains were isolated from eight different patients (Table 1). This indicated that a sporadic emergence of these strains occurred with little transmission of enterobacterial isolates. Besides K. pneumoniae, other common KPC-producing Enterobacteriaceae, including E. coli, K. oxytoca, C. freundii, E. cloacae, E. amnigenus, S. marcescens, and P. mirabilis, have been detected in China.3,4,9,10,13,14 In this study, we report for the first time of the emergence of KPC-producing E. aerogenes in China. Among the 12 patients with KPC-2-producing pathogens, 2 of them died from blood and pulmonary infections. However, these two patients were presented with complex symptoms when admitted to the hospital. Therefore, the extent by which KPC-producing Enterobacteriaceae contributed to the patients' death remains unclear.
It is known that KPCs alone may not confer resistance, but only reduce susceptibility to carbapenems. Impaired outermembrane permeability is one contributing factor associated with the resistance of KPC producers to carbapenems. 6 In this study, most of the clinical strains exhibited high level of resistance to carbapenems (Table 2), whereas their transconjugants were sensitive or had low-level resistance to IPM, MEM, and ETP (Table 2). These observations indicate that different combinations of KPCs and other mechanisms contribute to the carbapenem-resistant phenotype. Besides blaKPC-2, all isolates carried other resistant determinants, including class A and D β-lactamases, PMQR, and 16S rRNA methylase genes (Table 2). These mechanisms might contribute significantly to the resistant phenotypes observed by the isolates analyzed in this study. For example, CTX-M can mediate resistance to oxyimino-cephalosporins (CTX and FEP) or even to carbapenems when found in combination to impaired outermembrane permeability, 7 while ArmA or RmtB can lead to high level of resistance to aminoglycosides.
The blaKPC-2-carrying plasmids vary dramatically in size among the different strains (Fig. 1 and Table 2). For strains with a 40-kb blaKPC-2-carrying plasmid (E. coli IR1401, E. coli IR1405, E. aerogenes IR1502, and K. oxytoca IR1509), the genetic environments of the blaKPC-2 gene were quite variable (Table 2). Similar results have also been observed in another study. 14 Therefore, the different plasmid profiles suggested that the spread of the blaKPC-2 gene might not be mediated by plasmid exchange between the different strains. However, most of the clinical isolates can transfer the blaKPC-2-carrying plasmids to the E. coli J53 recipient, indicating the interspecies transferability of the plasmids.
In 2009, a novel genetic background of the blaKPC-2 gene was observed in China. These included genes on the plasmid pKP048 that encode for an integration structure of a Tn3-based transposon and partial Tn4401 segment. The genes include Tn3-transposase, Tn3-resolvase, ISKpn8, blaKPC-2, and an ISKpn6-like element. 9 For most strains analyzed in this study, the genetic environment of the blaKPC-2 gene was consistent with pKP048. Two other variants that had 321 bp and 221 bp of truncated blaTEM inserted upstream of the blaKPC-2 gene were also identified in this study (Table 2 and Fig. 2). It has also been reported that the genetic environments of the blaKPC-2 among most KPC-producing Enterobacteriaceae isolated from China were similar to pKP048. Most of them carried Tn3-based transposons involving the blaKPC gene, with or without an inserted fragment of truncated blaTEM located upstream of the blaKPC gene.4,9,14 The genetic context of the blaKPC genes among the common clinical enterobacterial isolates from China has similar transposon structures found in Tn4401. These have been identified as an active transposon capable of mobilizing blaKPC genes at high frequency without target specificity. 2 This suggested that the spread of the blaKPC-2 gene between the different enterobacterial species in China is primarily mediated by horizontal transfer of Tn3-based transposons. The genetic environments of blaKPC-2 gene for the three isolates remain unknown (Table 2), indicating that novel structures might contribute to the exchange of blaKPC-2 gene between the different enterobacterial species. Further studies on the genetic environment of blaKPC-2 in these clinical isolates may provide additional insight to its transmission mechanisms.
Footnotes
Authors' Contributions
J.Y. did molecular genetic analysis, plasmid profiling, and drafted the article. L.Y., L.G., Q.Z., R.C., and Y.L. participated in the collection of strains and clinical details. Y.C. and J.Z. participated in the genetic environment analysis. X.H. and S.T. participated in the PCR and PFGE. L.H. participated in the design and organization of the study. All authors read and approved the final article.
Acknowledgments
The study was supported by the National Key Program for Infectious Diseases of China (2008ZX10004-001-C) and National Nature Scientific Foundation No. 81102168 from the Ministry of Science and Technology, China.
Disclosure Statement
All authors declare that they have no conflicts of interest.