
Abstract
Introduction
E
In recent decades, the problem of increasing antibiotic resistance of ExPEC strains has emerged. Since 1983, when the first extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae strains were described in Germany, 24 production of these enzymes have spread worldwide 4 and different types of TEM, CTX-M, and SHV enzymes have been defined. 58 Presently, the most common ESBL appears to be CTX-M-15 and CTX-M-14. CTX-M-15 (first detected in India in 2001 23 ) has become the most common β-lactamase in Italy (2006 36 ; 2008 6 ), Norway (2007 37 ), Switzerland (2007 27 ; 2012 19 ), Korea (2009 51 ), India (2011 47 ), Nigeria (2011 59 ), Czech Republic (2012 14 ), Bulgaria (2012 30 ), and others. On the other hand, CTX-M-14 has become most common in China (2007 21 ; 2011 62 ), Spain (2007 38 ; 2009 40 ; 2010 13 ; 2011 34 ), Lao People's Democratic Republic (2012 53 ), and others. Global expansion of CTX-M-15-producing E. coli is mainly due to a clone belonging to the multilocus sequence type ST131, which exhibits a specific antigen O25b. 44 E. coli O25b-ST131 clones have variable pulsed-field gel electrophoresis (PFGE) patterns with at least 68% similarity and belong to the B2 phylogenetic group. 39 The O25b-ST131 strains do not harbor genes encoding typical virulence factors like P-fimbriae, cytotoxin necrotizing factor 1, and α-hemolysin; however, they do possess less common virulence genes such as fimH, fyuA, iha, iutA, kpsM, malX, ompT, sat, traT, and usp. 39
Some bacteriocin types, including colicin E1 50 and microcins H47, M, and V, are also considered to be potential virulence factors (or virulence markers) in E. coli strains, 1 however, until now, only the distribution of the cvaC gene (encoding the microcin V precursor) in ESBL-producing E. coli has been studied. 12
In this study, uropathogenic ESBL-producing E. coli strains, isolated in Brno, Czech Republic, were characterized in detail, including PFGE profiles, antibiotic resistance, virulence determinants, and E. coli phylogroups, and the prevalence of genes encoding bacteriocin.
Materials and Methods
Bacterial strains
E. coli strains were isolated, between 2008 and 2011, from UTIs of 109 patients (St. Anne's University Hospital, Brno, Czech Republic) living in the South Moravia region of the Czech Republic. Each isolate was collected from a different patient. Clinical samples were collected after patients provided informed consent. The study was approved by the ethics committee of the Faculty of Medicine, Masaryk University, Czech Republic. Bacterial identification was carried out using the ENTEROtest24 (Erba Lachema, Czech Republic). Control bacteriocin producer strains used for the polymerase chain reaction (PCR) method and indicator strains used for screening of bacteriocin production were previously described in detail. 50
Isolation of ESBL- and AmpC-producing E. coli strains
ESBL-producing E. coli strains were identified based on Screening and Confirmatory tests for ESBL (CLSI Document M100-S16). 10 Briefly, a plate containing Mueller Hinton (MH) agar (HiMedia) was inoculated with a bacterial suspension (0.5 McFarland standard). Ceftazidime (30 μg), ceftazidime/clavulanic acid (30/10 μg), cefotaxime (30 μg), and cefotaxime/clavulanic acid (30/10 μg) antibiotic discs were then placed on the MH agar. An inhibition zone diameter ≥5 mm, in the presence of clavulanic acid, indicated an ESBL strain. The results for ESBL-producing E. coli were confirmed on Oxoid Brilliance ESBL agar (OX; Oxoid). 20
PCR amplification and sequencing of blaSHV, blaTEM, and blaCTX-M genes
The 109 ESBL-producing UPEC strains were examined with global CTX-M, 15 SHV, and TEM primers. 5 Moreover, CTX-positive strains were amplified with CTX-M type-specific primers. 46 All obtained PCR products were sequenced and analyzed. 54
PCR detection of the E. coli O25b-ST131 clone
Screening for the E. coli O25b-ST131 clone was performed as previously described by Clermont et al. 9 The pabB gene (347 bp fragment) was used as a marker of this clone, which was amplified using a pair of specific primers (O25pabBspe.F: 5′-TCCAGCAGGTGCTGGATCGT-3′ and O25pabBspe.R: 5′-GCGAAATTTTTCGCCGTACTGT-3′). The trpA gene was used as a PCR-positive control and was amplified using trpA.F primers: 5′-GCTACGAATCTCTGTTTGCC-3′ and trpA2.R: 5′-GCAACGCGGCCTGGCGGAAG-3′. 9
Antibiotic susceptibility testing
Antibiotic susceptibility of 109 ESBL-producing UPEC strains was determined using the disk diffusion method in MH agar. Twenty-two antimicrobial agents were tested (ampicillin—AMP 10 μg; amoxicillin/clavulanic acid—AMC 30 μg; piperacillin/tazobactam—TZP 100 μg; cephalothin—KF 30 μg; cefotaxime—CTX 30 μg; cefoperazone/sulbactam—SCF 75/30 μg; ceftazidime—CAZ 30 μg; cefepime—FEP 30 μg; aztreonam—ATM 30 μg; ertapenem—ETP 10 μg; imipenem—IPM 10 μg; meropenem—MEM 10 μg; chloramphenicol—C 30 μg; doxycycline—DO 30 μg; tigecycline—TGC 19 μg; gentamicin—CN 10 μg; amikacin—AK 30 μg; oxolinic acid—OXO 30 μg; ciprofloxacin—CIP 5 μg; colistin—CT 10 μg; trimethoprim/sulfamethoxazole—SXT 25 μg; and nitrofurantoin—F 300 μg) using the zone size criteria based on the Performance Standards for Antimicrobial Susceptibility Testing (CLSI document M100-S20). 11
Pulsed-field gel electrophoresis
XbaI (New England Biolabs) PFGE was performed on 109 ESBL isolates using the PulseNet protocol (CDC 2002). PFGE profiles were analyzed using BioNumerics fingerprinting software (Applied Math). Based on the PFGE data, dendrograms were constructed using Dice's coefficient of similarity and UPGMA clustering at 1% tolerance. E. coli isolates with a Dice's similarity index ≥85% were assigned to one cluster. 57
Multilocus sequence typing
Multilocus sequence typing (MLST) was performed following amplification of DNA corresponding to seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA). One representative strain from each of the 17 PFGE clusters was typed by MLST. Protocols and primer sequences were obtained from the E. coli MLST database website (http://mlst.ucc.ie/mlst/dbs/Ecoli).
Screening for bacteriocin production
Screening for bacteriocin production was performed using a previously described method.33,50 Briefly, each of the 109 E. coli strains was inoculated in parallel on four different agar plates and incubated at 37°C for 48 hours. The bacteria were then killed using chloroform vapors, overlaid with a top agar layer containing indicator bacteria, and incubated at 37°C for 24 hours; then, bacteriocin producers were identified.
PCR detection of bacteriocin genes
Detection of 31 different bacteriocin determinants (colA, colB, colD, colE1, colE2, colE3, colE4, colE5, colE6, colE7, colE8, colE9, colIa, colIb, colJs, colK, colL, colM, colN, colS4, colU, colY, col5, col10, micB17, micC7, micH47, micJ25, micL, micM, and mV) was completed using a previously described method.33,50 Briefly, genomic DNA from bacteriocin-producing bacteria was used for the PCR; all bacteriocin producers were also tested using the colony PCR method. Because microcins H47 and M are sensitive to chloroform vapors, 43 all 109 E. coli strains were subjected to PCR with specific primers capable of detecting these microcin determinants. For sequentially diverse colicin E1 determinants, 49 primers cea2F (5′-GGTGGAACTGGAGGTAGCAA-3′) and cea2R (5′-ACGTCGTTGTTGTTCTGCTTCCT-3′), which amplified the 357 bp PCR product, were used. The 449 bp-long colicin L PCR product was detected with primer pairs Col28b(L)-F (5′-TGCATATTGAAAGCGTCAGC-3′) and Col28b(L)-R (5′-CAGGTTATCCCCTCTCACCA-3′). For microcin M (166 bp PCR product) detection, primer pairs M-DS2009-F (5′-CGTTTATTAGCCCGGGATTT-3′) and M-DS2009-R (5′-GCAGACGAAGAGGCACTTG-3′) were used. The following protocol was used for PCR amplification: 94°C (2 minutes; for colony PCR, the initial step was extended to 5 minutes); 94°C (30 seconds), 60°C (30 seconds), 72°C (1 minute), 30 cycles; 72°C (7 minutes). PCR products of sequentially related bacteriocins (colicins E2-9, Ia-Ib, U-Y, 5–10) were subjected to dideoxy terminator sequencing and sequences were analyzed with Lasergene software (DNASTAR, Inc.).
Phylogenetic analysis of E. coli strains
The method published by Clermont et al. 8 was used for phylogenetic analysis of E. coli (A, B1, B2, or D phylogenetic groups). Briefly, E. coli strains were divided into four phylogenetic groups based on the presence of chuA, yjaA genes, and the TSPE4.C2 fragment. 8
Screening for genes encoding virulence factors
All 109 ESBL-producing UPEC strains were screened for the presence of 17 different virulence factors: α-hemolysin (α-hly), afimbrial adhesion I (afaI), aerobactin synthesis (aer, iucC), cytotoxic necrosis factor (cnf1), S-fimbriae (sfaI), P-fimbriae (pap), aggregative adherence plasmid (pCVD432), invasivity antigen locus (ial), heat-labile enterotoxin (lt), heat-stabile enterotoxin (st), bundle-forming pili (bfpA), intimin (eaeA), invasion-plasmid H (ipaH), fimbriae type I (fimA), Shiga-toxin 1 (stx1), Shiga-toxin 2 (stx2), and enterohemolysin (ehly). Details regarding PCR protocols and primers sequences were previously described.3,26,29,31,41,42,48,61
Statistical analysis
For statistical analysis of the bacteriocinogeny, phylogenetic groups, and virulence factors, standard methods derived from the binomial distribution, including the two-tailed Fisher's exact test, were used. STATISTICA version 8.0 (StatSoft) was used for calculations.
Results
Molecular and PFGE analysis of ESBL-producing UPEC strains
The UPEC strains used in this study were isolated from 2008 to 2011 from UTIs of 109 patients being treated at a regional hospital in Brno, Czech Republic.
Among the 109 ESBL-producing UPEC strains, 106 strains (97.2%) produced CTX-M β-lactamases and only three strains (2.8%) produced SHV β-lactamases. The molecular types of identified β-lactamases are shown in Table 1. Most of the strains produced type CTX-M-15 (43 strains; 39.4%) and CTX-M-27 (35 strains; 32.1%) or a combination of these β-lactamase types with type TEM-1, including CTX-M-15/TEM-1 (19 strains; 17.4%) and CTX-M-27/TEM-1 (8 strains; 7.3%). A minority of strains produced SHV-12 (3 strains; 2.8%) and CTX-M-5 (1 strain; 0.9%).
In the majority of ESBL UPEC isolates (87.2%; 95 of 109), the pabB gene was detected and isolates were thus considered E. coli O25b-ST131. Based on pulse-field gel electrophoresis, 83 E. coli O25b-ST131 strains belonged to just three clusters with a Dice's similarity ≥85%, while 12 E. coli O25b-ST131 strains belonged to several clusters (Fig. 1 and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr). Cluster 1 contained 27 (100.0%) CTX-M-15-producing strains, cluster 2 contained 20 (87.0%) CTX-M-15-producing and three (13.0%) CTX-M-27-producing strains, and cluster 3 contained 33 (100%) CTX-M-27-producing strains (Fig. 1 and Supplementary Fig. S1). In addition to these three clusters, the remaining 26 E. coli strains belonged to 14 different clusters.
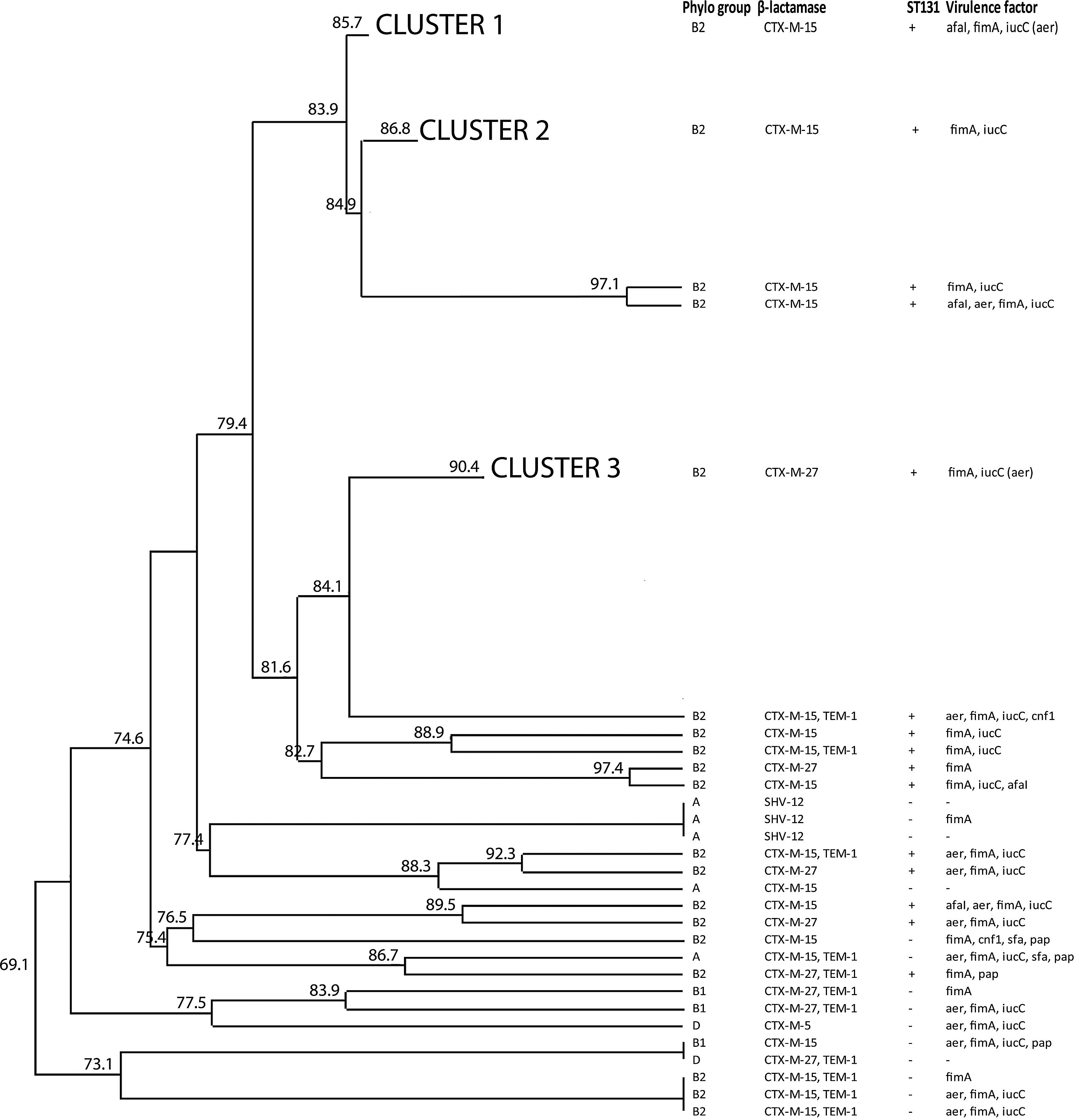
Pulsed-field gel electrophoresis-based dendrogram of 109 extended-spectrum β-lactamase (ESBL)-producing uropathogenic Escherichia coli strains. E. coli isolates with a Dice's similarity index ≥85% were considered to be a cluster. Eighty-three E. coli O25b-ST131 were grouped into three clusters (Clusters 1, 2, 3) with Dice's similarity ≥85%, while 12 other E. coli O25b-ST131 strains were distributed among several clusters.
MLST of E. coli strains from different PFGE clusters
One representative isolate from each of the 17 PFGE clusters comprising E. coli isolates 21, 22, 34, 66, 95, 104, 128, 138, 144, and 155, was analyzed using MLST. All isolates revealed identical sequences identified as sequence type ST131. The allelic profile 53-40-47-13-36-28-29 was identified in all tested isolates.
Antibiotic resistance of ESBL-producing UPEC strains
All the 109 ESBL-producing E. coli strains were resistant to ampicillin, amoxicillin/clavulanic acid, piperacillin/tazobactam, cephalothin, cefotaxime, cefoperazone/sulbactam, ceftazidime, and aztreonam; they were susceptible to carbapenems (ertapenem, imipenem, meropenem) and colistin. Resistance to other tested antibiotic types was different in individual ESBL-producing isolates (Supplementary Fig. S1); isolates resistant to cefepime, ciprofloxacin, trimethoprim/sulfamethoxazole, oxolinic acid, and doxycycline (Table 2) were the most common. Within the PFGE clusters, differences in resistance to tested antibiotic types were found. Taken together, non-O25b-ST131 strains (compared with O25b-ST131 strains) were more frequently resistant to chloramphenicol (Supplementary Fig. S1, p=0.002). Among the O25b-ST131 strains, CTX-M-15 strains were more frequently resistant to trimethoprim/sulfamethoxazole (p=0.01) compared with CTX-M-27 strains.
All E. coli strains were susceptible to ETP, IPM, MEM, CT and resistant to AMP, AMC, TZP, KF, CTX, SCF, CAZ, ATM. Resistance to CIP, FEP, SXT, OXO, DO, TGC, F, C, CN, and AK differs within E. coli isolates.
PFGE, pulsed-field gel electrophoresis; AMP, ampicillin; AMC, amoxicillin/clavulanic acid; TZP, piperacillin/tazobactam; KF, cephalothin; CTX, cefotaxime; SCF, cefoperazone/sulbactam; CAZ, ceftazidime; ATM, aztreonam; CIP, ciprofloxacin; FEP, cefepime; SXT, trimethoprim/sulfamethoxazole; OXO, oxolinic acid; DO, doxycycline; TGC, tigecycline; F, nitrofurantoin; C, chloramphenicol; CN, gentamicin; AK, amikacin; ETP, ertapenem; IPM, imipenem; MEM, meropenem; CT, colistin.
Phylogenetic groups of ESBL-producing UPEC strains
All ESBL-producing UPEC strains belonging to the three major PFGE clusters were in the B2 phylogenetic group (83/83) (E. coli B2-O25b-ST131) (Supplementary Fig. S1 and Table 2). The remaining 26 E. coli strains were distributed across 14 different clusters. In these strains, the B2 phylogenetic group was most common (61.5%) followed by group A (19.2%), B1 (11.5%), and D (7.7%). E. coli strains O25b-ST131 producing either β-lactamase CTX-M-15 or CTX-M-27 were exclusively (100.0%; 95 of 95) associated with the phylogenetic group B2 (Table 2).
Virulence factors of ESBL-producing UPEC strains
Virulence determinants varied among tested E. coli strains and also among strains within the three major PFGE clusters (Supplementary Fig. S1 and Table 2). Genetic determinants encoding fimbriae type I, afimbrial adhesin I, and aerobactin production were frequently found among the tested strains. All three virulence factors were found in 70.9% (39 of 55) of O25b-ST131 strains producing β-lactamase CTX-M-15, while E. coli O25b-ST131 producing CTX-M-27 encoded these three virulence factors in only 5.0% (2 of 40) (p<0.01) of the strains. Afimbrial adhesin 1 was most commonly found in CTX-M-15 strains (62.9%; 39 of 62), while fimbriae type 1 and aerobactin production was found in 81.4% (35 of 43) of CTX-M-27-producing strains (Supplementary Fig. S1 and Fig. 1).
Bacteriocin production of ESBL-producing UPEC strains
Production of bacteriocins (bacteriocinogeny) was identified in 16 out of 109 ESBL-producing UPEC strains (14.7%); 3 (out of 55; 5.5%) producers were found among the E. coli O25b-ST131 CTX-M-15 strains and 6 (out of 40; 15.0%%) among the E. coli O25b-ST131 CTX-M-27-producing strains (Supplementary Fig. S1). Among non-O25b-ST131 E. coli, a significantly higher incidence of bacteriocinogeny, 50.0% (7 out of 14), was found compared with E. coli O25b-ST131 CTX-M-15 (p<0.01) and E. coli O25b-ST131 CTX-M-27 strains (p=0.02). E. coli O25b-ST131 CTX-M-15 strains were found to encode 2 different colicin types (E7, Ia), while E. coli O25b-ST131 producing CTX-M-27 encoded 3 different colicins (E1, E7, and Ia) and 1 microcin type (B17) (Supplementary Fig. S1 and Table 2).
Discussion
In the last 30 years, E. coli strains producing ESBL have spread worldwide. 24 The largest increase has been in the CTX-M enzyme-producing strains, which are more common in many European countries compared with TEM- and SHV-producing strains. 28
Our study focused on the South Moravian region of the Czech Republic where we mainly found isolates similar to the multiresistant international clone E. coli B2-O25b-ST131. These isolates mostly produced either β-lactamase CTX-M-15 or CTX-M-27. The E. coli B2-O25b-ST131 strains producing CTX-M-15 are well known and have been isolated worldwide. 39 In contrast, the E. coli B2-O25b-ST131 CTX-M-27 clone was first detected in 2011 in China. 55 By 2012, in Japan, these clones had become more widespread (41%) than CTX-M-15-producing ST131 clones (28%). 32 The CTX-M-15- and CTX-M-27-producing E. coli O25b-ST131 isolates were distributed in several PFGE clusters with at least 74.6% PFGE similarity. A similar finding was previously published by Nicolas-Chanoine et al., where E. coli ST131 strains belonged to PFGE clusters at a similarity level of 68%. 39 While the CTX-M-15 isolates were connected to cluster 1, CTX-M-27-producing isolates were found in cluster 3 and both CTX-M-15 and CTX-M-27 isolates were found in cluster 2 and other clusters. Since isolates taken from all PFGE clusters revealed identical sequences belonging to the ST131 clone, this finding suggests a higher discrimination power for PFGE compared with MLST.
The frequent isolation of E. coli B2-O25b-ST131 CTX-M-27 among patients in South Moravia suggests an efficient transfer of this lineage from other regions. Interestingly, E. coli B2-O25b-ST131 CTX-M-27 was found, in 2012, in samples taken from great cormorants and mallards and it is possible that these water birds represent a vector for transport of these particular E. coli strains. 56 It is therefore possible that CTX-M-27 starts to outcompete CTX-M-15 by direct spread of CTX-M-27-producing lineages, by introduction of new E. coli strains, through horizontal gene transfer of the CTX-M-27 resistance plasmid to recipient E. coli cells or by some combination of these factors.
PCR detection of the pabB gene in E. coli isolate 168 was repeatedly negative, while the PFGE profile of this isolate was 86.3% identical to the PFGE profile of isolate 21. Both PCR and MLST typing clearly indicated that isolate 21 belongs to the O25b-ST131 clone. Although the reasons for not detecting the pabB gene in E. coli isolate 168 remain unknown, one possible explanation involves the presence of point mutation(s) in the pabB primer target site(s). Isolates 9 and 72 were in different phylogroups, D and B1, respectively. Although both isolates have identical PFGE profiles, the chuA gene (indicating phylogroup D) 8 was not detected in isolate 72. This negative PCR result in isolate 72 is likely a result of a mutated primer target site.
In our study, ESBL-producing E. coli clones O25b-ST131 belonged to the phylogenetic group B2. This phylogenetic group is typical for extraintestinal pathogenic strains 2 with higher prevalence of virulence factors. 7 Clonal types E. coli B2-O25b-ST131 producing CTX-M-15 39 and CTX-M-27 56 were previously classified to phylogroup B2. Moreover, previous studies have shown that the majority of UPEC strains also belong to the B2 phylogroup. 50 Only a minority of strains populate the other phylogroups, indicating that these are not related to the most frequent isolates.
Previous studies of ESBL-producing ST131 strains identified a number of virulence genes, including fimH, fyuA, iha, iutA, kpsM, malX, ompT, sat, traT, and usp. 39 Most of the strains in our study encoded type I fimbriae (fimA), aerobactin synthesis genes (aer, iucC), genes encoding afimbrial adhesin I (afaI) and, rarely, other virulence factors (cnf1, sfa or pap). The relatively small differences in the presence of virulence determinants found among isolates collected in our study indicate a genetic similarity among these strains.
A high prevalence of bacteriocinogenic strains (54.0%) was found among previously characterized E. coli strains isolated from human UTIs. 50 Moreover, it has been found that two chromosomally encoded microcin (H47 and M) and plasmid-encoded colicin E1 production increase the urovirulence of E. coli strains.1,50 In contrast to these data, we found a low prevalence of bacteriocin genes among ESBL-producing UPEC strains. This finding likely reflects the fact that most of the strains tested in our study have a clonal character. An additional reason is plasmid incompatibility between some ESBL-encoding plasmids and colicin/microcin-encoding plasmids. The ESBL-encoding plasmid pEK516 (encoding CTX-M-15, OXA-1, and TEM-1) 60 belongs to the IncIIF group that is similar to the colicin-encoding plasmid pColB2 17 and plasmid-encoding microcin V. 22 In fact, none of the colicin B producers and only three microcin V producers were found among the strains we tested.
As shown by PFGE, at least three different epidemiologically related lineages belonging to the same sequence type (ST131) were found among ESBL-producing UPEC strains in the South Moravian region of the Czech Republic. Fourteen groups of isolates, comprising a total of 41 individual isolates, were identified as identical when all tested parameters of E. coli were combined. In other isolates, only limited variability was observed. The limited variability of detected molecular types of E. coli phylogroups and of the virulence and bacteriocin determinants suggests a common origin for the analyzed isolates with an ongoing process of genetic diversification during multiplication within and outside of hosts.
Footnotes
Acknowledgment
This work was supported by a grant from the Ministry of Health of the Czech Republic (NT13413-4/2012) to D.S.
Disclosure Statement
The authors declare they have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.