
Abstract
Helicobacter pylori is the major cause of peptic ulcers and gastric cancer in humans. Treatment involves a two or three drug cocktail, typically including amoxicillin. Increasing levels of resistance to amoxicillin contribute to treatment failures, and higher levels of resistance are believed to be due to multiple genetic mutations. In this study, we examined the progression of spontaneous genetic mutations that contribute to amoxicillin resistance in H. pylori when exposed to increasing concentrations of amoxicillin in vitro. During the selection process, we isolated five strains each of which had progressively higher levels of resistance. Using a whole genome sequencing approach, we identified mutations in a number of genes, notably pbp1, pbp2, hefC, hopC, and hofH, and by sequencing these genes in each isolate we were able to map the order and gradual accumulation of mutations in these isolates. These five isolates, each expressing multiple mutated genes and four transformed strains expressing individually mutated pbp1, hefC, or hofH, were characterized using minimum inhibitory concentrations, amoxicillin uptake, and efflux studies. Our results indicate that mutations in pbp1, hefC, hopC, hofH, and possibly pbp2 contribute to H. pylori high-level amoxicillin resistance. The data also provide evidence for the complexity of the evolution of amoxicillin resistance in H. pylori and indicate that certain families of genes might be more susceptible to amoxicillin resistance mutations than others.
Introduction
C
Previous studies have demonstrated that certain point mutations in penicillin-binding protein-1 (PBP1) confer resistance to β-lactam antibiotics in H. pylori.12,13,20,24,28,29 However, these studies, and others, demonstrate that PBP1 point mutations can only account for low levels of amoxicillin resistance, suggesting that high levels of resistance must require additional resistance mechanisms.7,18
In this study we track the emergence of a high-level amoxicillin-resistant strain of H. pylori in vitro. Using whole genome sequencing and comparison to the parental amoxicillin sensitive strain, we identified changes that occurred within the bacterial chromosome affecting amoxicillin susceptibility. By sequencing strains with intermediate levels of amoxicillin resistance, which emerged on the path to the highly resistant strain, we established the sequence in which these mutations originated. Our results provide evidence for the complexity of the evolution of amoxicillin resistance in H. pylori, and identify a variety of genes, which can contribute to this resistance.
Materials and Methods
Bacterial strains and culture conditions
H. pylori ATCC strain 26695 was used as the amoxicillin-sensitive reference strain and the parental strain to create the in vitro resistant isolates. All bacterial strains were incubated at 37°C in a humidified 10% CO2 incubator. Cultures were streaked for isolation on Brucella agar (Becton-Dickinson Microbiology) supplemented with 5% defibrinated sheep blood (Colorado Serum Company). Liquid cultures were prepared by suspension of H. pylori colonies in Brucella broth (Difco Laboratories) supplemented with 10% heat-inactivated fetal bovine serum (Gibco Bethesda Research Laboratories) and 1% IsoVitaleX (Becton-Dickinson Microbiology). Cultures were routinely passed by dilution into fresh media at 48 hr intervals. Freezer stocks were prepared by resuspending 48 hr bacterial cultures in 1% proteose peptone-20% glycerol, flash frozen in liquid nitrogen, and kept at −80°C.
General DNA techniques
All DNA manipulations were performed according to standard protocols. Primers used in this study were created using the published sequence of genes from 26695 and Vector NTI (Invitrogen) and synthesized by Sigma-Aldrich. Polymerase chain reaction (PCR) was performed in an automated thermal cycler PTC100 (MJ Research), using the Choice-Taq Blue™ DNA polymerase (Denville Scientific). PCR products were run on 1% agarose gels (Biorad) and subsequently cut and purified using the Minelute agarose purification system (Qiagen, Inc.). Sequencing of purified DNA products was performed by the Genomics Core facility at the University of California, Riverside. Sequence analysis and alignments were performed using Vector NTI.
Isolation of amoxicillin-resistant strains
Strain 26695, an amoxicillin-sensitive strain with a fully sequenced genome, 32 was used as the starting culture to create amoxicillin resistant isolates. Serial twofold dilutions of amoxicillin were prepared in Brucella broth supplemented with 10% heat-inactivated fetal bovine serum and 1% IsoVitalex in a 96-well plate. Wells contained sequentially diluted amoxicillin concentrations ranging from 16 to 0.006 mg/L. H. pylori 26695, with an inoculum OD600 (optical density at 600 nm) of 0.5±0.05 was inoculated at a 1:100 dilution (1.5 μl in 150 μl, ∼106 cells). The 96-well plates were incubated at 37°C in a humidified 10% CO2 incubator for 2 days and wells were examined for visible signs of growth. A 1.5 μl aliquot of bacteria from the well with the highest concentration of amoxicillin showing growth was used to inoculate the next plate prepared the same as above. The process was repeated every 48–72 hr for ∼9 months. Isolates were saved at various time points and colony purified starting with bacteria expressing a minimum inhibitory concentration (MIC) of 0.50 mg/L of amoxicillin and resulted in five isolates labeled IS1–IS5. MICs to amoxicillin of all isolates were determined using the broth microdilution method according to a previously published protocol. 10 MIC values were also determined in 0.5 mg/L increments for amoxicillin concentrations between 1 and 8 mg/L.
Whole genome sequencing
An Illumina genomic DNA sample prep kit (Illumina, Inc.) was used to create the genomic library. All subsequent reactions were performed according to instructions. Five micrograms of genomic DNA from the final isolate was sheared by sonication to obtain fragments of about 400 bp. The ends were repaired by converting overhangs from fragmentation into blunt ends, using T4 DNA polymerase and E. coli DNA polymerase I Klenow (New England Biolabs). Further, an A base was added to the 3′ end of the blunt phosphorylated DNA fragments using the polymerase activity of Klenow. Specific DNA adapters were ligated to the ends of the DNA fragments, to prepare them to be hybridized to a flow cell. The ligation product was purified using a PCR purification kit (Promega). This purified product was submitted to the Genome center at University of California at Riverside (UCR) where it was sequenced with the Illumina Genome Analyzer using single read at 32 bp per read. The data were then put through the flowcell pipeline in the form of short reads. These short reads were assembled and analyzed for single nucleotide polymorphisms (SNPs) and single base pair Insertions or Deletions (InDels) by the Bioinformatics department at UCR. The alignment put forth was also visually inspected to locate any changes missed by data mining.
Identifying potential targets and their evolution
SNPs and single insertions and deletions were analyzed for mutations resulting in amino acid changes within genes. Mutations in the last six amino acids of a protein or not within a coding or promoter region were excluded. Potential target genes were sequenced in the parental strain 26695 to identify mutations present prior to the in vitro amoxicillin-resistant isolation process. PCR primers were created for each of the potential targets and synthesized by Sigma Genosys. The potential target genes were then PCR amplified from the genomic DNA of each isolate and sequenced as mentioned above. The sequencing data were analyzed to confirm the presence or absence of the target mutations in each isolate.
Transformation of H. pylori
Transformation of 26695 was achieved by electroporation, following a previously established protocol by Wang et al. 36 Competent cells were prepared by growing 26695 on Brucella agar supplemented with 5% defibrinated sleep blood for 24–36 hr. Cells were isolated and washed thrice with 20% glycerol, pelleting for 5 min at 3,100 g between washes. Washed cells were resuspended in ∼500 μl 20% glycerol and separated into 100 μl aliquots. The entire open reading frame of each gene of interest was PCR amplified from IS5 genomic DNA. PCRs were purified by PCR purification kit (Promega) and ∼500 ng purified DNA was combined with a 100 μl aliquot of prepared electrocompetent cells. The cells and DNA mixture was transferred to a 1 mm gap electroporation cuvette and incubated on ice for 10 min. Using an ECM 630 electroporator, cells were transformed at 2.5 kV, 600 Ω, and 25 μF, which resulted in a time constant ∼13 msec. Cells were resuspended in 200 μl SOC media and plated onto Brucella-sheep blood agar, and recovered for 36–48 hr. Cells were scraped from the plate and resuspended in sterile phosphate-buffered saline (PBS) followed by plating on Brucella-sheep blood agar containing either 0.125 or 0.25 μg/ml amoxicillin. Individual colonies were isolated and targeted genes were sequenced to verify mutations.
[14C] penicillin G accumulation experiments
[14C] penicillin G accumulation experiments were performed using log-phase Brucella-blood agar cultures of 26695, IS3, and IS5 suspended in PBS, pH 7.4 (PBS) buffer to a final concentration of 3–5×109 bacteria/ml, and incubated with [14C] penicillin G to a final concentration of 1 μCi/ml at 37°C. Half milliliter aliquots were removed at 1, 5, 10, and 30 min, the bacteria were pelleted by centrifugation, and washed twice with PBS to remove unaccumulated antibiotic. Cells were subsequently resuspended in 0.1 ml of PBS and added to 10 ml of scintillation fluid (Fisher Scientific). The amount of antibiotic accumulated was measured in counts per minute using a Beckman LS6500 liquid scintillation counter (Beckman Coulter).
[14C] penicillin G efflux experiments
Using actively growing cells, 4 ml bacterial cultures of IS5 were suspended in PBS buffer to a final concentration of 3–5×109 bacteria/ml. The cell suspension was divided into two halves and one half was incubated with 40 μM carbonyl cyanide m-chlorophenylhydrazone (CCCP) for 15 min before the addition of [14C] penicillin G to a final concentration of 1 μCi/ml at 37°C to both halves. Half milliliter aliquots were removed at 1, 5, 10, and 30 min. The cells were then washed and analyzed as mentioned above.
Efflux pump inhibitor assay
Using the microdilution MIC method mentioned above, susceptibility of 26695, IS2, IS3, IS4, and IS5 to amoxicillin was studied in the presence or absence of naphthylmethyl-piperazine (NMP) or phenyl-arginine-b-naphthylamide (PAβN) at 100 mg/L. For IS2, IS3, IS4, and IS5, MIC values between 1 and 8 mg/L were determined in increments of 0.5 mg/L. For transformants containing individual mutations, MIC values between 0.03 and 16 mg/L were determined in twofold dilution increments.
Results
Amoxicillin-resistant isolates and mutations in each isolate
Sequential passage of H. pylori 26695 in increasing levels of amoxicillin resulted in a gradual increase in resistance culminating in a strain (isolate 5, IS5), which expressed an amoxicillin MIC 64× higher than the parental strain's MIC value of 0.06 mg/L (see Figure 1). In the process of in vitro selection, various isolates were obtained and colony purified starting with isolate IS1 with an MIC of 0.50 mg/L and ending with IS5 with an MIC of 4.0 mg/L (Fig. 1).
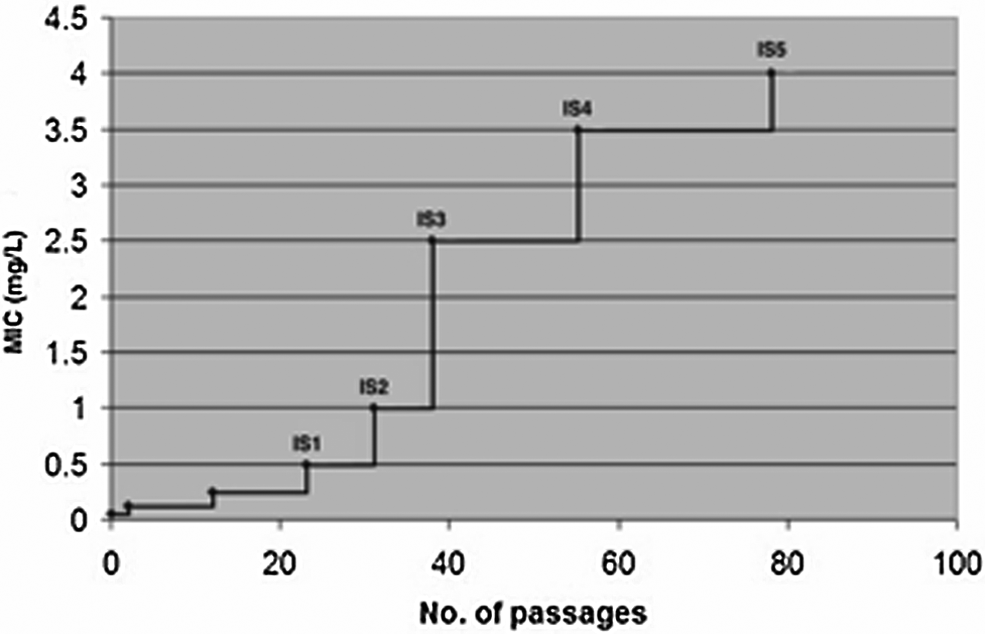
Evolution of amoxicillin resistance in Helicobacter pylori 26695 by sequential passage in increasing levels of amoxicillin. At various levels of amoxicillin resistance, bacteria were isolated and colony purified, starting with isolate IS1 with a minimum inhibitory concentration (MIC) of 0.50 mg/L and ending with IS5 with a MIC of 4.0 mg/L.
Since the 26695 H. pylori genome has been completely sequenced, 32 this strain was an ideal candidate for our studies since it would make comparisons and identification of potential amoxicillin resistance genes easier to identify. Each of the in vitro selected amoxicillin isolates (IS1–IS5) maintained their MIC value after regrowth from freezer stocks, suggesting the presence of stable mutations, rather than transient or unstable changes, which would be difficult to characterize. A stable increase in MIC suggested that the changes occurred at the genomic level as opposed to changes in transcriptional levels with response to stimuli.
Initial studies focused on the IS5 isolate. To determine the sequence changes conferring resistance to IS5, potential target genes such as PBPs 1–3, Hop A-E, and Mre ABC genes were sequenced and compared to those in the parental strain 26695. However, only three of these genes, pbp1, pbp2, and hopC, from IS5 showed amino acid changes when compared to 26695 (see Table 1). The sequence of PBP1 (HP0597) from IS5 showed two amino acid changes: P372S and T438M. Upon sequencing the pbp1 genes in the five isolates, IS1–IS5, P372S substitution appeared in IS1, whereas the T438M mutation appeared first in IS4 (Table 1). Sequence comparisons of PBP2 in each of the five isolates (IS1–IS5) and parental strain 26695 revealed E536K mutation only in IS5 (Table 1). Sequence comparisons of Hop C in each isolate and 26695 revealed a single mutation resulting in the conversion of an arginine to a histidine, R302H (Table 1), which appeared first in IS2 (Table 1).
MIC determinations and presence of various mutations, which appeared in isolates IS1 to IS5. The bold+sign indicates the mutation first occurred in that isolate.
MIC, minimum inhibitory concentrations.
Judging by the level of total resistance, and the presence of five isolates with distinct MICs, it seemed plausible that there were other changes, which had not yet been detected. In an attempt to locate those changes, whole genome sequencing of IS5 was employed. Unlike de novo genome sequencing, a cost-efficient, template-based short read genome sequencing was employed. Short reads are assembled against a previously published reference genome of H. pylori 26695 NC_000915, which is 1.667 Mbps. The sequence coverage over the entire genome was greater than 30×.
Data mining identified ∼30 SNP mutations and ∼50 single and double amino acid Insertions or Deletions (InDels). Further analysis of SNPs excluded mutations within the coding region that did not translate into amino acid changes, and mutations in the last six amino acids of the coding region; this analysis eliminated about one-third of the SNPs. InDels were further analyzed to exclude any changes that were in the last six amino acids of the coding region or not present within a coding, or promoter region of a gene, allowing us to eliminate two-thirds of the InDel mutations. Twenty-four SNP and InDel mutations remained. These mutations were then sequenced in the H. pylori amoxicillin-sensitive parental strain 26695, to ensure that the changes observed were not present before selection of resistant isolates. Thirteen of these mutations were present in the amoxicillin-sensitive parental strain eliminating them as potential resistance targets.
It can be challenging to look for InDels of unspecified lengths. So to cover the entire genome and acquire any information that data mining may have missed, we scrolled through the alignment of genome with its short basepair reads using samstools on the biocluster server at University of California, Riverside. The alignment file showed high quality short reads and alignment. Scrolling through the alignment helped to identify four deletions of about 20–30 bp. These were also present in the parental strain 26695 and were not further analyzed.
A total of 9 genes and 11 mutations emerged as potential resistance targets including PBP1, PBP2, and HopC. The loci of these changes, the precise amino acid change(s) that occurred, and the function of these genes are shown in Table 1. Two genes contain two mutations each, namely PBP1 and acriflavin resistance protein (HefC). Further sequencing of these mutations in IS1–IS5 showed the order in which they evolved (Table 1). IS1 contains the P372S mutation in PBP1 and a mutation in HP0181, which is a putative colicin V protein. Of the 11 mutations, 4 mutations, HefC, Hop C, Hypothetical Na+/H+ antiporter, and Flagellin A, appeared first in IS2. IS3 has an additional mutation in HefC and a mutation in the promoter region of DNA glycosylase. IS4 contains the T438M mutation in PBP1, while IS5 contains mutations in PBP2 and HofH, a putative outer membrane protein.
[14C] penicillin G accumulation
To determine whether amoxicillin resistance could be attributed to changes in antibiotic access to its periplasmic target, we performed radiolabeled penicillin G accumulation experiments. We had determined in previous studies9,10 that penicillin G could be used as a surrogate β-lactam drug for amoxicillin because it had similar binding affinities for PBP1. As shown in Figure 2, at each time point examined, isolates IS3 and IS5 accumulated less penicillin than the parental amoxicillin-sensitive strain 26695. Over the four time points examined, penicillin G accumulation levels in IS3 averaged 82.5% of that seen in 26695, while IS5 had accumulation levels averaging 68% of that of 26695.
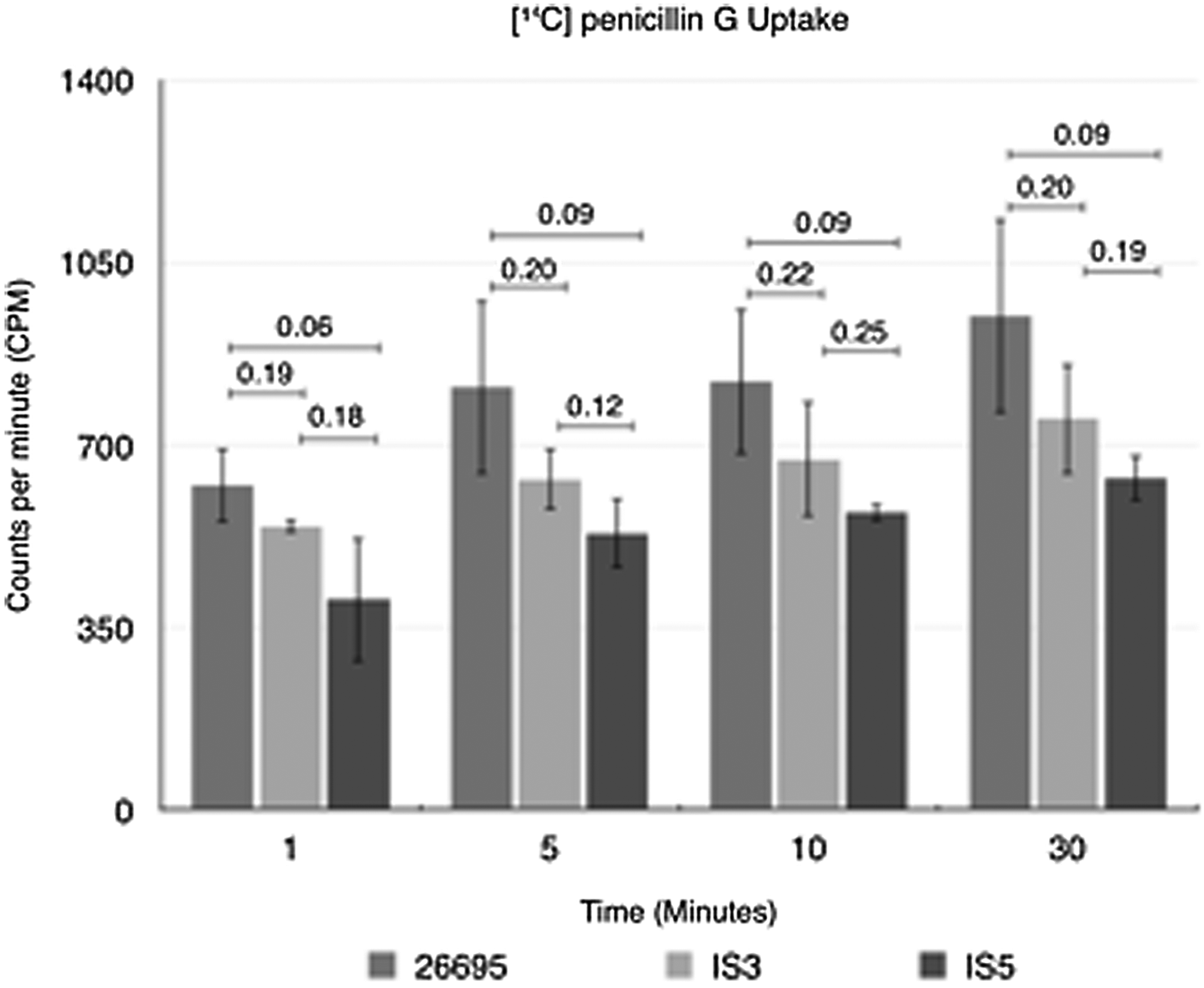
Isolates IS3 and IS5 accumulate less [14C] penicillin G than 26695 at each time point examined. Data shown represent the mean±standard deviation based on at least three separate experiments. Statistical numbers represent p-values determined using the Student's t-test between respective samples.
Changes in penicillin accumulation could result from either reduced entry of antibiotic into the periplasm because of altered porin proteins (such as HopC), reduced binding of antibiotic to PBPs or the active efflux of antibiotic from the periplasm due to an efflux pump mechanism. To address the last possibility, the [14C] penicillin G accumulation assay was performed on isolate IS5 in the presence or absence of the proton pump inhibitor CCCP as described in the Materials and Methods section. However, we did not detect any difference in penicillin accumulation for isolate IS5 with or without CCCP treatment (data not shown).
Finally, we examined the effect of efflux pump inhibitors (EFIs) on amoxicillin susceptibility using isolated strains and the broth microdilution MIC method described previously. To increase sensitivity in this assay, 0.5 mg/L increments were used between 1 and 8 mg/L amoxicillin. The amoxicillin MIC for strains 26695, IS2, IS3, IS4, and IS5 were determined when incubated in the presence or absence of either 100 mg/L NMP or PAβN. As shown in Table 2, the presence of EFI did not affect the MIC for strain 26695 or IS2. The IS2 strain contains the D131E HefC mutation, which, in this circumstance, does not appear to contribute to amoxicillin resistance. In contrast, isolate IS3, which contains both HefC mutations, D131E and L378F, was more sensitive to amoxicillin when grown in the presence of either EFI, reducing the MIC by 1.0 mg/L (from 2.5 to 1.5 mg/L). Similarly, isolates IS4 and IS5, which also contain both HefC mutations, were also slightly more amoxicillin sensitive in the presence of an EFI, with a MIC change of 1.0 mg/L for both IS4 (3.5–2.5 mg/L) and IS5 (4.0–3.0 mg/L).
Representative of at least three replicates of this experiment.
EFI, efflux pump inhibitor; NMP, naphthylmethyl-piperazine; ND, not done; PAβN, phenyl-arginine-b-naphthylamide.
To more directly assess the impact of each of the mutated genes, we attempted to transform 26695 with individually mutated genes isolated from IS5, in particular pbp1, hofHhefC, and pbp2; we have previously demonstrated that HopC mutations directly affect amoxicillin sensitivity. 7 We were able to create individual transformants containing the mutated PBP1, HofH, and HefC mutations found in IS5, but were unable to select a PBP2 mutant transformant. As shown in Table 3, the PBP1 mutated gene, containing both PBP1 mutations, increased resistance 8× (from 0.06 to 0.5 mg/L), whereas the HofH mutation increased amoxicillin resistance 16× (from 0.06 to 1.0 mg/L). Transformation of 26695 with HefC containing the single L378F mutation increased amoxicillin resistance 8× (from 0.06 to 0.5 mg/L); transformation with HefC containing both mutations, D131E and L378F, only increased this resistance to 1.0 mg/L. In addition, the presence of the efflux inhibitor, NMP, did not have a significant impact on amoxicillin resistance provided by PBP1 or HofH; in contrast, this inhibitor caused a dramatic reversion to the 26695 levels of susceptibility for both HefC mutants.
Representative of at least three replicates of this experiment. n/a, not applicable.
Discussion
The goals of this study were to characterize the evolution of amino acid changes in H. pylori genes when exposed to increasing levels of amoxicillin in vitro, and to identify the mechanisms of resistance contributed by these mutated genes. Resistance to β-lactam antibiotics among Gram-negative bacteria is typically due to one or more of the following mechanisms: drug inactivation by β-lactamases, target site (i.e., PBP) alterations, diminished permeability, and efflux. 26 Previous studies in our laboratory 10 and others 18 have demonstrated that H. pylori strain 26695 does not contain β-lactamase activity, one of the most common mechanisms of β-lactam resistance in bacteria.
As described in the Results section we successfully isolated five amoxicillin resistant strains, with progressively increasing resistance levels and identified a total of 11 amino acid changes within 9 genes, which we believe account for the majority of the observed amoxicillin resistance. In addition, using individual mutants, this study has demonstrated the significance of specific mutations within PBP1, HofH, and HefC that impact amoxicillin susceptibility.
IS1 has an MIC value of 0.5 mg/L and contains two mutations. The P372S in HP0597 (PBP1) is located adjacent to the penicillin-binding module (PBM) sequence SAIK, which contains the putative catalytic Ser 368 in the middle of the putative catalytic site. 28 This mutation at this site suggests that it interferes with the binding of Ser 368 to amoxicillin. This finding was not too surprising as changes in PBP1 has been the most common mechanism of amoxicillin resistance seen in H. pylori.12,13,20,24,28,29 Although a variety of specific amino acid changes have been detected, those mutations found adjacent to highly conserved PBMs, such as P372S, are associated with amoxicillin resistance. 13 The second mutation is in HP0181, which is a Colicin V protein. This is a frame-shift mutation, a deletion of a cytosine, 86 amino acids into a 226 amino acid protein, potentially knocking out the function of the protein. It is not apparent how mutations in this protein might play a role in conferring resistance to β-lactams.
IS2 has an MIC value of 1.0 mg/L, and it was found to contain four additional mutations in comparison to strain IS1. D131E is the first of two mutations occurring in HP0607, HefC protein, a multidrug efflux protein, which cooperates with two other proteins forming a multidrug efflux pump (discussed further below). A previously known target, HP0912, also known as Omp20 or HopC, contains a single point mutation at R302H. We have previously demonstrated that mutations in this gene increase resistance in H. pylori and decrease the level of penicillin accumulation within the cell. 7 Decreased membrane permeability is a contributing factor to β-lactam resistance in a wide array of Gram-negative bacteria. 26 IS2 also has a point mutation in HP0946, a conserved membrane protein and a homolog of Nhac; Na+/H+ antiporter; anion permease. A direct connection has not been made between this protein and β-lactam resistance; however, differences in expression levels of similar ion transport proteins have been seen when exposed to amoxicillin. 15 The point mutation seen in HP0958, a post-transcriptional regulator for Flagellin A (FlaA) protein, also seems like an unlikely target to confer resistance to β-lactams; however, a study has shown that two genes involved in production of flagella are differentially expressed when the cells are exposed to amoxicillin. 15
The third isolate IS3 shows an MIC value of 2.5 mg/L and possesses two additional changes from above. HP0602, a 3-methyl DNA glycosylase, possesses a single base pair change in its promoter region. There is no evidence that this gene plays any role in conferring resistance to β-lactams. The L378F amino acid substitution is the second change in HefC protein.
A role for efflux pumps in β-lactam resistance is widely distributed among Gram-negative bacteria. 26 A previous study by Bina et al. 4 identified three putative restriction-nodulation-division (RND) efflux systems in H. pylori—hefABC, hefDEF, and hefGHI—representing analogs of the AcrAB-TolC RND efflux system in E. coli; however, they concluded that active efflux does not affect antibiotic resistance in these bacteria. In contrast, van Amsterdam et al. 35 reported that mutations in the TolC (hefA) protein of H. pylori resulted in metronidazole resistance, while Liu et al. 19 reported that hefA is involved in multidrug resistance in H. pylori. In a related strain H. hepaticus, hefA was found to increase resistance to amoxicillin. 3 Recent studies by Trainor et al. 34 demonstrated that the HefC efflux pump in H. pylori provides some resistance to bile salts and ceragenins. Collectively, this study and previous studies point to a role for active efflux pumps in H. pylori resistance to various compounds, including antibiotics.
To investigate the role of several proteins including HefC, the accumulation of penicillin was measured in 26695, IS3, and IS5. Penicillin accumulation assays showed that IS3 and IS5 had a marked decrease in the amount of antibiotic present within the cell when compared with the parental strain 26695. Accumulation assays, however, do not distinguish between decrease in uptake, reduced binding to PBPs, or presence of an efflux mechanism. Initial attempts to identify efflux mechanisms using the proton motive force inhibitor CCCP in IS5 were unsuccessful. One possible explanation for this is that H. pylori was unable to grow in CCCP, but lack of inhibition with CCCP has also been seen in other bacteria with efflux mechanisms. 25
A more direct analysis was done by measuring the MIC of IS3, IS4, and IS5 in the presence of EFI. Each strain exhibited a marked decrease of 1.0 mg/L, when exposed to either of two EFIs. This reduction was not observed in IS2, the mutant strain containing a single mutation in HefC at D131E. This suggests that the presence of the L378F HefC mutation in IS3, IS4, and IS5 is, in part, responsible for the level of amoxicillin resistance. The use of EFIs is not without precedence. Hirata et al. 16 reported that the MIC for clarithromycin was decreased in H. pylori in the presence of PAβN, the same EFI used in this study. The mutations in HefC were further analyzed by transformation of the mutants into the amoxicillin-sensitive parental strain 26695. Using a 26695 strain transformed with HefC containing both mutations, D131E and L378F, there was a 16× increase in amoxicillin resistance. A similar transformant containing only the L378F mutations in HefC had an 8× increase in amoxicillin resistance. Moreover, the resistance in both transformants is abolished in the presence of NMP further supporting the hypothesis that HefC functions as an efflux pump protein. These results indicate that part of the amoxicillin resistance shown might be related to an efflux mechanism. The crystal structure of AcrB, a HefC homolog, in E. coli shows a symmetric trimer, which complexes with AcrA (HefB) and TolC (HefA) to form an efflux pump.8,27 Previous studies have shown that a single V-F mutation in AcrB increases E. coli resistance to several antibiotics. 6 Using homology modeling we compared the HefC protein in H. pylori to the AcrB protein in E. coli. The L378F mutation mapped within a helix toward the inside of the cavity, a site that might affect amoxicillin efflux through this protein. 33 The Leu to Phe change results in steric clashes with neighboring side chains, forcing a rearrangement of the secondary structure to accommodate the larger phenyl ring. The change in secondary structure could open the cavity to β-lactams,22,27 increasing efflux. This impact would be increased if HefC functions as a homotrimer, like its homolog AcrB.
IS4 contains a single additional mutation T438M in PBP1, which has been previously characterized and shown to have a fourfold increase in resistance to β-lactams. This mutation has been shown to map within the putative binding cleft of PBP1 of H. pylori. 28 When the pbp1 gene with 2 mutations from IS5 was transformed into 26695, it increased amoxicillin resistance 8×, further supporting the important role of PBP1 mutations in amoxicillin resistance.
Two additional mutations are present in the isolate IS5. One of these mutations E536K is present in HP1565 commonly known as PBP2. Because this mutation results in the substitution of the negatively charged glutamic acid with the positively charged amino acid lysine, this change would result in an overall positive charge increase in PBP2. This increased positive charge outside of the binding site can result in deflection of the antibiotic from its intended location. It has been previously shown that a charge increase in a PBP can result in an increase in resistance. 28 The region around the E536K mutation seems to be conserved in some related bacteria (data not shown), suggesting functional importance of this site within PBP2. The role of PBPs other than PBP1 in conferring resistance to β-lactams has been previously shown in several other bacteria. 17 Unfortunately, we were unable to select a 26695 transformant with only the mutated pbp2 gene, so at this time we can only speculate on its importance in amoxicillin resistance.
A G228W mutation is found in HP1167, which is a putative outer membrane protein known as Omp3 or HofH. This point mutation occurs at a highly conserved glycine residue and would likely impact protein structure and potentially function. The presence of this mutation transformed into strain 26695 increases amoxicillin resistance 16×. In addition, the efflux inhibitor NMP was able to decrease amoxicillin resistance in the HofH transformant from 1.0 to 0.25 mg/L. This might explain the decreased penicillin accumulation seen in IS5 relative to IS3 (Fig. 2)—however, further studies are needed to document this role for HofH.
In summary, based on this study and previous literature, it appears that the PBP1 P372S mutation in IS1 is responsible for the first increase in MIC. In IS2 several mutations occur, however, the mutation in HopC, R302H is most likely to contribute to the increased MIC in IS2. As this study has demonstrated, the L378F mutation in HefC contributes to amoxicillin MIC and likely causes the increase in MIC observed in IS3. Since only a single new mutation is introduced in IS4, the mutation in PBP1 at T438M, this mutation obviously contributes to the increase seen at this stage. The increase in MIC observed in IS5 could be caused by a combination of both PBP2 and HofH mutations, though this study has demonstrated the significance of the HofH mutation at G228W.
Therefore, we hypothesize that the majority of high-level amoxicillin resistance in isolate 5 (IS5) can be explained by the combined effects of amino acid changes occurring in PBP1, HopC, HefC, HofH, and possibly PBP2. It remains possible that additional mutations, not identified in our sequencing strategy, can also contribute to amoxicillin resistance in IS5.
The pattern of high levels of β-lactam resistance in H. pylori and other bacteria has been shown to involve a variety of different genes from different classes. 26 In H. pylori, cell division proteins have the highest percentage (∼39%) of essential genes, and cell envelope proteins (∼25%) compared with outer membrane proteins (only ∼3% are shown to be essential). 31 PBP1 is potentially an essential protein with an upper threshold of mutations before normal cell function is affected. A mutation resulting in decreased binding of PBPs to β-lactams could decrease its functionality, limiting the extent to which amino acid changes can be made, and placing selective pressure on other genes. Previous studies have suggested that the fitness cost imposed by antibiotic resistance can be compensated by other mutations allowing the organism to survive. 2
Low levels of β-lactam exposure have been shown to drive the evolution of resistant bacteria. 1 This exposure could be from improper prescription or administration of antibiotics, or from antibiotics found in food or another environmental source. β-lactams are a valuable class of antibiotics due to their widespread use, broad spectrum application, and low cost. We hope that a comprehensive understanding of β-lactam resistance in H. pylori would allow the development of better methods for its control and detection.
Footnotes
Acknowledgment
This study was supported by funding from the Division of Biomedical Sciences, School of Medicine, University of California, Riverside.
Disclosure Statement
No competing financial interests exist.