
Abstract
Pseudomonas aeruginosa is the main cause of mortality in cystic fibrosis patients and eradication of its biofilm represents a substantial problem clinically. In this study, biofilm of a cystic fibrosis strain P. aeruginosa PACI22 was established and confocal laser scanning microscopy was utilized for biofilm visualization. A quantitative time-kill biofilm model was implemented in vitro to assess the biocidal effect of ciprofloxacin, clarithromycin, and their combination at concentration levels ranged from 0.5× to 64× minimum biofilm inhibitory concentrations (MBIC) against the biofilm and the mean log bacterial densities (Log CFU/ml) retrieved from the biofilm were monitored by frequent sampling at 0, 3, 6, 9, 12, and 24 hr throughout the experiment. The results revealed that none of the tested antibiotics alone could completely eradicate the biofilm-ensconced bacteria at 0.5–64× MBIC values after 24 hr of treatment. Conversely, ciprofloxacin-clarithromycin combination at 32–64× MBIC entirely exterminated the biofilm. Furthermore, a substantial in vitro synergism between ciprofloxacin and clarithromycin against the biofilm was experimentally verified. This promising synergism affords scientific rationale for further in vivo investigations to evaluate the therapeutic potential of this combination for treatment of chronic pulmonary infections caused by P. aeruginosa biofilms.
Introduction
B
Materials and Methods
Bacterial strain and antimicrobial agents
P. aeruginosa mucoid strain (PACI22) used in this study was recovered from de-identified cystic fibrosis patients and all experiments of the current study were performed in a biosafety level two laboratory at Eastern Virginia Medical School. Before each experiment, P. aeruginosa PACI22 was subcultured twice on Tryptic Soy Broth (TSB) and incubated for 16 hr at 37°C. The inoculum was prepared in cation-adjusted Mueller Hinton II (CAMH) broth and diluted to visually match 0.5 McFarland equivalence (1.5×108 colony forming units [CFU]/ml) standard (Thermo Scientific Remel, Lenexa, KS) using black-white card provided with the kit. The initial inoculum was confirmed by the conventional plate count onto Mueller Hinton agar (BD, Franklin Lakes, NJ) plates. Ciprofloxacin and clarithromycin powders were purchased from Sigma-Aldrich (St. Louis, MO). Stock solutions (16 mg/ml) of ciprofloxacin and clarithromycin were prepared and stored at −80°C according to the manufacturer's instructions. Samples of the stock solutions were thawed at room temperature and diluted in CAMH broth before each experiment.
Planktonic susceptibility testing
The minimum inhibitory concentrations (MICs) of the tested antimicrobial agents were determined using the broth microdilution method as described by Clinical and Laboratory Standards Institute (CLSI) guidelines. 12 The experiments were performed in polystyrene, round-bottom, 96-well microplates Thermo Scientific™ Nunc (Thermo Fisher Scientific, Inc., Waltham, MA). Twofold serial dilutions of the antibiotics were prepared and the final bacterial count in each well was 5×105 CFU/ml. The MIC was defined as the lowest concentration of the antibiotic that resulted in no visible growth after aerobic incubation at 37°C for 24 hr.
Biofilm susceptibility testing
Biofilm formation
Seventy-five microliter inoculums of P. aeruginosa PACI22 (1.5×108 CFU/ml) in TSBG (TSB supplemented with 2% glucose) were inoculated in polystyrene, round bottom, 96-well microplates (Thermo Fisher Scientific, Inc.) and incubated at 37°C for 24 hr. After incubation, the supernatant was carefully aspirated and the wells were washed twice with sterile phosphate buffered saline (PBS) solution. 10
Minimum biofilm inhibitory concentration
One-hundred microliters of twofold serial dilutions of the antibiotics in CAMH broth were transferred to the wells with the established P. aeruginosa PACI22 biofilm. After incubation for 18 hr at 37°C, the plates were visually inspected for bacterial growth indicated by the presence of turbidity. Owing to the transparent nature of the P. aeruginosa PACI22 biofilm, it had no interference with the visual plate inspection. Similar to planktonic susceptibility testing, 12 microbial growth/inhibition was easily discriminated in minimum biofilm inhibitory concentration (MBIC) experiments. The MBIC was defined as the lowest concentration of the antibiotic that resulted in no visible growth after 18–20 hr of incubation. 11
Minimum biofilm bactericidal concentration
Broth aliquots of 10 μl from wells showed no visible growth were transferred into a sterile 96-well microplate and diluted with 90 μl of CAMH broth to minimize the antibiotic carryover effect. After incubation for 24 hr at 37°C, the plates were inspected for bacterial growth. The minimum biofilm bactericidal concentration (MBBC) was defined as the lowest concentration of the antibiotic (ciprofloxacin or clarithromycin) that prevented visible growth. 44 In MBIC and MBBC experiments, the antimicrobial effects of ciprofloxacin and clarithromycin were assessed on the established P. aeruginosa PACI22 biofilm. Consequently, growth of bacteria in a particular well indicates regrowth of planktonic bacteria from the surviving biofilm. Accordingly, the MBBC value represents the lowest concentration at which bacteria fail to regrow.
Confocal laser scanning microscopy
Although P. aeruginosa biofilms may develop at the air–liquid interface, the medically relevant and experimentally addressed biofilms are those that develop at the liquid–solid interface either under static or dynamic conditions.7,19,26,29,31,40,45 For analysis of P. aeruginosa PACI22 biofilm, overnight broth culture of P. aeruginosa PACI22 in CAMH broth was diluted in PBS to 1.5×108 CFU/ml using 0.5 McFarland equivalence standard (Thermo Scientific Remel). The cell suspension was used to inoculate TSBG broth at final bacterial count of 1.5×106 CFU/ml and aliquots (1.5 ml) of the inoculated TSBG broth was distributed in 24-well polystyrene plates (Corning Costar, Corning, NY). The plates were incubated for 24 hr at 37°C without shaking. After incubation, the supernatant was carefully aspirated and the wells were washed twice with 2 ml PBS to remove the unattached or loosely attached cells. Fixation of P. aeruginosa PACI22 biofilm was conducted at 4°C for 3 hr with 10% glutaraldehyde (Fisher Scientific, Waltham, MA) in PBS. After rinsing the wells with PBS 3×, the biofilm was stained with 0.1% propidium iodide (MP Biomedicals, Santa Ana, CA) in PBS as described previously. 25 All microscopic observations and image acquisitions of the fixed P. aeruginosa PACI22 biofilm were performed at the bottom of the wells by confocal laser scanning microscopy (CLSM) LSM 510 META (Carl Zeiss, Jena, Germany) with an argon laser at 480 nm and a filter at 560 nm. The z-series images were captured at 5 μm intervals and five representative fields measuring 512×512 μm were analyzed for the biofilm. Fluorescence projections and sections through the biofilm were generated using the Metamorph Version 7.5.6 software package (Molecular Devices LLC, Sunnyvale, CA). The program is adapted with the image processing, multidimensional acquisition, color alteration mode, digital contrast adjustment, region configuration, and distance calibration options. Image data including image stack, the number of images in the stack, filtering method, (pixel size)x, (pixel size)y, and (pixel size)z (distance between two consecutive images in the stack) were saved as an Access Database file. Stacking of the consecutive images constructed the three-dimensional matrix of the biofilm.
Biofilm time-kill assays
Time-kill assays for P. aeruginosa PACI22 biofilm were performed according to the previously described method with some adaptations. 44 Briefly, P. aeruginosa PACI22 in TSBG broth was used to inoculate (500 μl/well of 1.5×106 CFU/ml) untreated polystyrene, 48-well microplates Thermo Scientific Nunc (Thermo Fisher Scientific, Inc.). The plates were incubated for 24 hr at 37°C under aerobic condition without shaking for establishment of the biofilm. The supernatant was then carefully aspirated and the wells were washed twice with 750 μL PBS. P. aeruginosa PACI22 biofilm was exposed to ciprofloxacin, clarithromycin, and their combinations at escalating concentrations of 0 (control), 0.5×, 1×, 2×, 4×, 8×, 16×, 32×, and 64× MBIC. All experiments were performed in triplicates. After time intervals of 0, 3, 6, 9, 12, and 24 hr of incubation at 37°C, the wells were sonicated for 3 min using 10% Pulsar of 550 Sonic Dismembrator (Fisher Scientific, Pittsburg, PA) to dislodge and disperse the microbial cells from the biofilm. Concerning control experiments at zero time, further increase of sonication power or time showed reduction of the mean log viable count (7.18 CFU/ml) of the harvested biofilm-ensconced P. aeruginosa PACI22 and this may be attributed to the microbial cell disruption. After sonication, samples of 100 μl were withdrawn and 10-fold serially diluted in sterile normal saline solution to minimize the antibiotic carryover effect through reducing the antibiotic concentration to sub-MIC levels. Samples of the cell suspensions (50 μl) were then plated onto CAMH agar plates to quantify the viable biofilm-ensconced bacteria. After incubation of the CAMH plates at 37°C for 24 hr, the viable cell counts as CFU were determined for different antibiotic-treated and antibiotic-untreated (control) biofilms. The logarithmic decrease in CFU from antibiotic-treated biofilms was calculated by means of the following formula: logarithmic decrease=[log(CFU control) − log(CFU x )], where x corresponds to the tested antimicrobial agent used alone or in combination. Synergistic activity between antibiotics was defined as a reduction in the number of CFU/ml of at least 2 logarithmic scales with regard to the activity of each antibiotic alone. Antagonism between antibiotics was defined as an increase in the number of CFU/ml of at least 2 logarithmic scales with regard to the activity of each antibiotic alone. Indifference was defined as a decrease in the number of CFU/ml of less than 2 logarithmic scales with regard to the activity of each antibiotic alone. 51
Statistical analysis
In this study all experiments were performed in triplicate and means of the kinetic data were presented graphically. Results were analyzed with two-tailed Mann–Whitney nonparametric test. Data output of analyses with p-values less than 0.01 were considered statistically significant. Statistical analyses in the current study were performed through SPSS, version 18.0 (SPSS, Inc., Armonk, NY).
Results
Confocal laser scanning microscopy of P. aeruginosa PACI22 biofilms
CLSM has been previously employed to assess biofilm thickness and surface coverage as fundamental criteria indicating successful biofilm formation.7,19,26,29,31,40,45 The reported biofilm thicknesses of 8 μm, 7 11–17 μm, 19 2–8 μm, 26 5–9 μm, 29 5–21 μm, 31 8–94 μm, 40 and 50–170 μm 45 significantly vary based on the tested P. aeruginosa strains, incubation periods, substratum materials, and other in vitro experimental conditions. In the current study, spatial biomass distribution of 24 hr P. aeruginosa PACI22 biofilm was illustrated by CLSM imaging (Fig. 1). The CLSM image was generated from a stack of images covering an area of 262,144 μm2 (512×512 μm) and each pixel is considered a box (voxel) with the dimensions (pixel size)x, (pixel size)y, and (pixel size)z and color alteration mode was utilized to improve the contrast. Furthermore, mean thickness and substratum coverage were determined for P. aeruginosa PACI22 biofilm. Mean of the maximum thickness (47.67±3.51 μm) reflects the spatial dimensions of the mature biofilm and entire substratum coverage (100%) indicates efficient colonization of the polystyrene surface by P. aeruginosa PACI22 biofilm. Hereby, the calculated biofilm thickness and substratum coverage, as the major criteria for the assessment of biofilm formation, confirmed the in vitro establishment of mature P. aeruginosa PACI22 biofilm.

Confocal laser scanning micrographs of Pseudomonas aeruginosa PACI22 biofilm.
Table 1 illustrates the differences between ciprofloxacin and clarithromycin susceptibilities demonstrated by biofilm-grown P. aeruginosa PACI22 compared to that of the same isolate when grown in planktonic culture. The results demonstrate that P. aeruginosa PACI22 biofilm was less susceptible to growth inhibition and killing by both ciprofloxacin and clarithromycin compared with that of the planktonic culture.
MIC, minimum inhibitory concentration of planktonic culture; MBIC, minimum biofilm inhibitory concentration; MBBC, minimum biofilm bactericidal concentration.
Consistent with the previously described intrinsic resistance of P. aeruginosa to clarithromycin by CLSI M100 S24, the planktonic (MIC=25 μg/ml) and biofilm (MBIC=100 μg/ml) cultures of P. aeruginosa PACI22 in the current study revealed noticeable resistances to clarithromycin. Based on CLSI criteria, the planktonic (MIC=0.05 μg/ml) and biofilm (MBIC=0.40 μg/ml) cultures of the same organism were considered sensitive to ciprofloxacin. On the other hand, MBBC of P. aeruginosa PACI22 biofilm against ciprofloxacin (MBBC=12 μg/ml) significantly exceeds the stated resistance threshold (≥4 μg/ml) of ciprofloxacin. 12 Furthermore, the MBIC and MBBC of ciprofloxacin against the biofilm were 8-folds and 240-folds, respectively, higher than MIC for the planktonic culture. In case of clarithromycin, the MBIC and MBBC against the biofilm were 4-folds and >32-folds, respectively, higher than MIC for planktonic culture of P. aeruginosa PACI22. The MIC and MBIC values are relatively consistent with that previously described by some investigators.3,18,56
Biofilm time-kill assays
Substratum material plays a crucial role in the development of the bacterial biofilms but owing to some technical difficulties in the experimental in vitro models, P. aeruginosa biofilm may be generated and addressed under dissimilar conditions within the same study as illustrated previously.7,19,26,40 For equivalent biofilm formation, polystyrene material was employed in the current study as a sole substratum material for the establishment of P. aeruginosa PACI22 biofilm in MBIC/MBBC experiments and the time-kill experiments and for the CLSM imaging. Biofilm time-kill assays were exploited to evaluate in vitro the ability of ciprofloxacin, clarithromycin, and their combination in eradicating P. aeruginosa PACI22 biofilm. Kinetic study involved administration of ciprofloxacin, clarithromycin, and their combination to P. aeruginosa PACI22 biofilm over 24 hr of treatment Figure 2. The mucoid P. aeruginosa PACI22 strain was used to establish the biofilm and antimicrobial activities of the tested antibiotics were assessed against such clinically relevant biofilm. The tested concentrations ranged from 0.2–12.8 and 50–6,400 μg/ml for ciprofloxacin and clarithromycin, respectively. Such concentration ranges were corresponding to 0.5–64× MBIC values of the tested antibiotics. In this study, the mean log bacterial density (Log CFU/ml) retrieved from P. aeruginosa PACI22 biofilm in the control experiments was 9.81±0.056 (± Standard Deviation) and ciprofloxacin generally demonstrated insignificantly (p>0.01) superior antibacterial profile to clarithromycin against the biofilm. Low concentrations (0.5–2.0× MBIC) of such antibiotics were not effective to induce prominent bacterial death (Log reduction <2) as compared to the control biofilm (Fig. 2). Regarding the antimicrobial activities of ciprofloxacin and clarithromycin after 24 hr of biofilm treatment, none of the tested antibiotics could completely eradicate the biofilm ensconced bacteria at 0.5–64× MBIC values. Conversely, no bacteria could be cultured from the biofilms when P. aeruginosa PACI22 biofilm was exposed to 32–64× MBIC ciprofloxacin-clarithromycin combination. Furthermore, reductions of the bacterial survivors in the biofilm after exposure to ciprofloxacin, clarithromycin, or their combination were distinctly in concentration-dependent manners (Fig. 2).
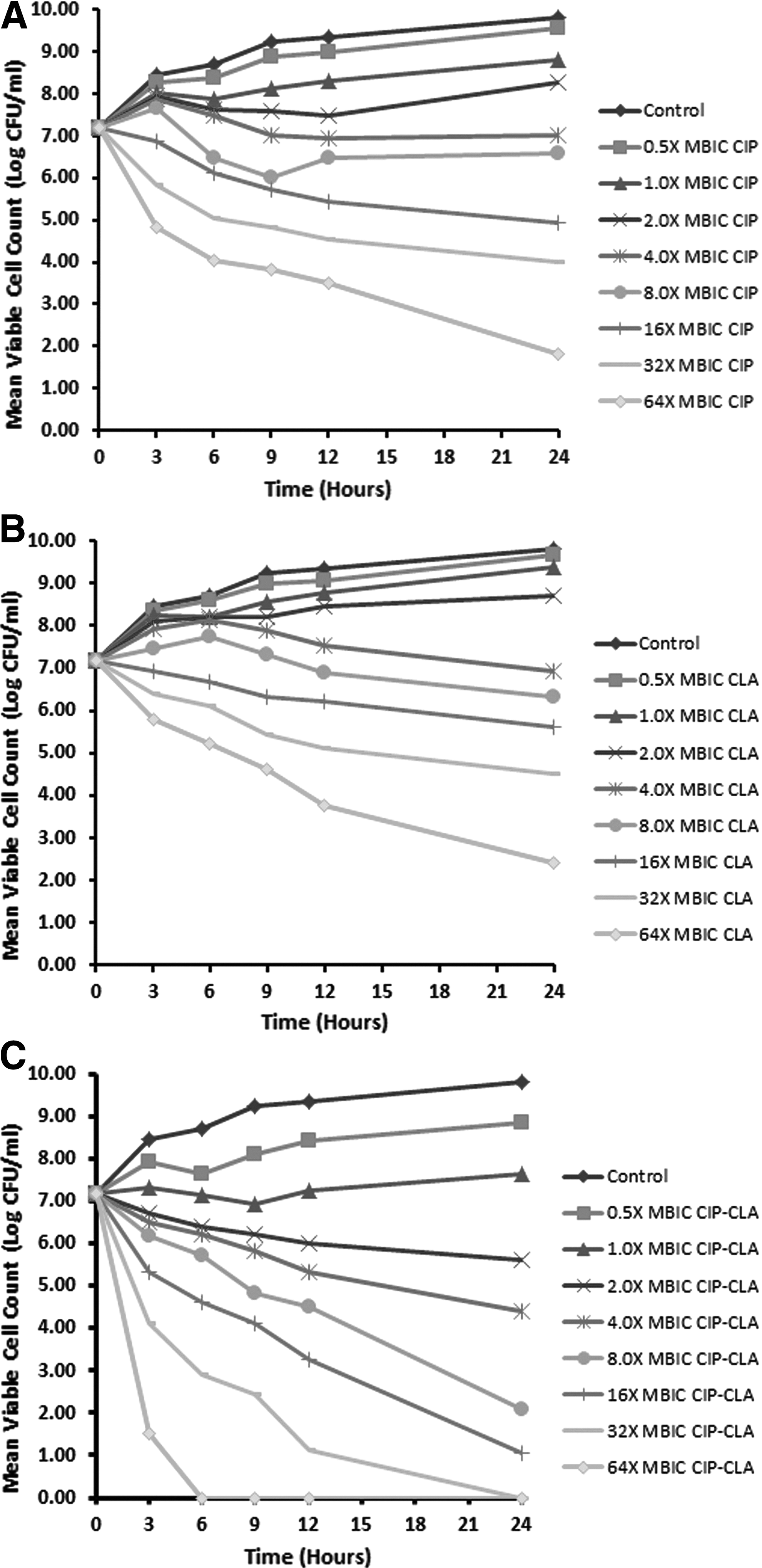
Kinetic study of the antimicrobial effects of ciprofloxacin, CIP
Depending on the interaction assumption, when P. aeruginosa PACI22 biofilm was exposed to ciprofloxacin-clarithromycin combination at different concentrations the results of these drug combination would be either synergistic, antagonistic, or without influence on the bactericidal activity if the effect of the combined drugs is superior than (Combined Log Reduction; C Log R ≥2), inferior than (C Log R ≤−2), or the same as that of the individual drug activities (C Log R <2 to >−2), respectively. As shown in Table 2, significant synergism (p<0.01) between ciprofloxacin and clarithromycin was observed at concentration levels of 2–64× MBIC with maximum calculated values of 4.47 and 4.55 logs at 8× MBIC and 16× MBIC for ciprofloxacin and clarithromycin, respectively.
Logarithmic Reduction=[log(CFUcontrol) − log(CFUx)], where x corresponds to the tested antimicrobial agent used alone (CIP and CLA) or in combination (CIP-CLA).
Combination Logarithmic Reduction (C log R)=[log(CFUCIP or CLA) − log(CFUCIP-CLA)]. C Log R ≥2 indicates synergism, C Log R <2 to >−2 indicates indifference, and C Log R ≤−2 indicates antagonism.
All numbers mentioned in the table are positive values. Results are statistically significant at p<0.01.
CFU, colony forming units; CIP, ciprofloxacin; CLA, clarithromycin; CIP-CLA, combination of ciprofloxacin and clarithromycin.
Discussion
Bacterial biofilms represent a global and predominant cause of both chronic infections and indwelling medical devices-associated infections. Such infections typically exhibit considerable resistance to antimicrobial agents rendering them challenging, or even impossible, to treat using conventional chemotherapeutic agents. 4 P. aeruginosa is considered as the principal cause of morbidity and mortality in patients with cystic fibrosis. 16 Furthermore, P. aeruginosa is the most common cause of intensive care unit-associated pneumonia and responsible for ∼2 million nosocomial infections that occur annually worldwide. 32 Biofilms of P. aeruginosa significantly impair antibiotic efficacy by protecting the bacteria not only from antibiotics but also from phagocytosis, opsonizing antibodies, complement, and ciliary action of the respiratory tract. Consequently, the frequency of antibiotic-resistant P. aeruginosa isolates present in clinical settings has been dramatically increased, thereby necessitating the demand for the development of alternative therapeutic strategies. 31
P. aeruginosa biofilms demonstrated resistance to the antibiotics ciprofloxacin, ceftazidime, and tobramycin at concentrations far greater than those necessary to eradicate planktonic bacteria.35,53 This antibiotic resistance may be due in part to persisting subpopulations within the biofilms that are protected from the antimicrobial agents.6,20,30,36 Biofilms are considered pertinent to the clinical settings because they play a key role in the ability of the organisms to tolerate antibiotics and persist in long-term chronic infections.9,52 The majority of studies have been conducted with reference to planktonic bacteria and rather less attention has been directed toward bacteria in the biofilm mode of growth. 4 In this study, the in vitro antimicrobial activities of ciprofloxacin, clarithromycin, and their combination against P. aeruginosa biofilm were evaluated.
The results of the current study revealed that biofilm-grown P. aeruginosa PACI22 was less susceptible to both ciprofloxacin (MBIC/MIC=8) and clarithromycin (MBIC/MIC=4) as compared to planktonically-grown culture. Similar studies demonstrated that P. aeruginosa isolates were more resistant to growth inhibition by single antibiotics when grown in biofilm cultures than when grown in planktonic ones.1,18,56 Therefore, the failure of conventional planktonic culture techniques to predict antibiotic susceptibilities may explain inability of the antimicrobial agents to eradicate P. aeruginosa biofilm-associated infections, such as in cystic fibrosis patients. In case of P. aeruginosa PACI22, eradication of the established biofilm was more challenging than its inhibition with ciprofloxacin (MBBC/MBIC=30) and clarithromycin (MBBC/MBIC ≥16) where the antimicrobial susceptibilities of biofilm-ensconced bacteria are severely hindered by the biofilm matrix. These results reaffirm the necessity of early aggressive antimicrobial therapy to avoid biofilm establishment. Once biofilms are well developed, the antibiotic concentrations required to inhibit (MBIC) or eradicate (MBBC) the biofilms may exceed the maximum permissible plasma concentrations and this finding is consistent with some previous reports.18,56 It has been reported that P. aeruginosa planktonic culture from sputum may mislead the clinicians to believe that the micro-organism is susceptible to the tested antibiotics, whereas under in vivo conditions of biofilm growth, the organism could be considerably more recalcitrant to the bactericidal effect of the antibiotics. 1
The development of bacterial biofilms has stochastic variables, and independent rounds of biofilm experiments may result in inexact biofilm architecture. The main factors influencing the biofilm development may include temperature, nutrient composition, flow rate (in dynamic experiments), and history of the cultures used for inoculation, thus considering such factors in the in vitro models significantly improves the experimental reproducibility. 24 In the present study, reproducibility was addressed through controlling the above-mentioned variables in the conducted experiments. Consistent with the results of Heydorn et al., 23 the CLSM revealed that P. aeruginosa PACI22 could effectively and uniformly colonize the substratum within 24 hr of incubation. Generally, the thickness of P. aeruginosa biofilm varies from less than 10 μm to more than 300 μm as described previously,4,19,26,29,41,45,54 and this prominent variability in the biofilm thickness arises from differences in the tested strains, pH, nutrients, oxygen limitation, incubation periods, static versus dynamic assays, and the overall experimental designs. The CLSM measurements for P. aeruginosa PACI22 revealed that average of the biofilm thickness is 47.67±3.51 μm and this is considered a relatively thick biofilm compared to that previously mentioned by some investigators.7,19,26,29,31,40 Clearly, the implemented culture method in this study could efficiently produce in vitro biofilm; however, the thickness of the biofilm using this method was less than the 170 μm height of biofilms, which were generated using continuous-flow cell culture chambers. 45 This is a potential limitation of the static techniques because biofilms grown under stagnant conditions may be less developed than multilayered biofilms grown using continuous-flow cell culture chambers and may therefore exhibit different metabolic properties. Nevertheless, the use of an individual flow cell culture method to grow biofilms for different antibiotics would be exceptionally impractical, as this technology is not suited for rapid antibiotic susceptibility testing and static biofilm methods would be more convenient and reproducible in this respect. 1
P. aeruginosa biofilm-associated infection in cystic fibrosis patients is challenging to antimicrobial therapy and this may explain the high morbidity and mortality in such infection. Consequently, a treatment capable of eliminating P. aeruginosa biofilm in chronic infections would significantly upturn the survival of cystic fibrosis patients.21,33 Nevertheless, this objective is quite beyond reach because antibiotic therapy for cystic fibrosis patients is bargained by a number of bacterial factors including the improved expression of efflux pumps that eliminate antibiotics and the deficiency of antibiotic penetration into bacterial biofilms that render the microorganism impervious to antibiotics. Extensive antibiotic therapy based on aerosolized antibiotics for instance tobramycin in combination with oral ciprofloxacin for management of the patients with pulmonary infections, has been applied. 8 This therapeutic approach may only postpone the development of chronic infection and just moderate the clinical symptoms or decelerate the loss of lung function. Accordingly, antibiotic therapy appears to be beneficial in different stages of cystic fibrosis, but eradication of P. aeruginosa biofilm in chronic infections is still away from achievement because the optimal protocols of antibiotic administration, either monotherapy or combinations, are hitherto to be determined. 51
Ciprofloxacin is considered as one of the most effective antibiotics against P. aeruginosa infections in cystic fibrosis patients.2,15 It is noteworthy that the clinical efficacy of long-term macrolide including the 14-membered ring clarithromycin on P. aeruginosa pulmonary infections has been reported. 49 To delay the emergence of bacterial resistance and to hinder the biofilm formation, drug combinations, rather than monotherapy, are currently advocated extensively. 51 Ciprofloxacin has been combined with some antibiotics to improve its antimicrobial activity against P. aeruginosa.38,48 In the current study, the results revealed that ciprofloxacin or clarithromycin, as single agents, could not eradicate P. aeruginosa PACI22 biofilm at the tested concentration levels of 0.5–64×MBIC but their combination demonstrated in vitro synergism and could exterminate the biofilm of P. aeruginosa PACI22 at 32–64× MBIC after 24 hr of treatment. It has been reported that tobramycin combination with clarithromycin and azithromycin revealed synergistic and antagonistic antimicrobial activities, respectively, on P. aeruginosa biofilm. Furthermore, the synergistic interaction of clarithromycin was not attributed to the anti-inflammatory properties of this macrolide but to its ability to preclude both the biofilm formation and the expression of quorum sensing system. 51 Some studies reported that macrolides at sub-inhibitory concentrations could modify the expression of multiple genes in P. aeruginosa.28,46 This probably explains, in the present study, why clarithromycin could significantly enhance the antimicrobial activity of ciprofloxacin against the biofilm of, a cystic fibrosis strain, P. aeruginosa PACI22.
Conclusion
In conclusion, results of this study revealed a substantial synergism between ciprofloxacin and clarithromycin, and their combination appears as a promising therapy to be clinically assessed for the treatment of chronic pulmonary infections caused by P. aeruginosa biofilms.
Furthermore, the study finding is therapeutically relevant because in clinical practice P. aeruginosa infections of cystic fibrosis patients have been treated with a minimum of two anti-pseudomonal drugs. Thus, ciprofloxacin-clarithromycin combination would be a potential therapeutic option in such infections. Further pharmacokinetic-pharmacodynamic studies are required for optimization of the dose regimens.
Footnotes
Disclosure Statement
No competing financial interests exist.